
Abstract
Long-distance migration is widespread among birds, connecting breeding and wintering areas through a set of stopover localities where individuals refuel and/or rest. The extent of the stopover is critical in determining the migratory strategy of a bird. Here, we examined the relationship between minimum length of stay of PVC-ringed birds in a major stopover site and the remaining flight distance to the overwintering area in the Eurasian spoonbill (Platalea l. leucorodia) during four consecutive autumn migrations. We also analysed the potential effect of timing (arrival date), as well as the role of experience in explaining stopover duration of spoonbills. Overall, birds wintering in Africa, and facing long-distance travel from the stopover site (ca. 3,000 km) stay for longer (2.7 ± 0.4 days) than Iberian winterers (1.5 ± 0.2 days) that perform a much shorter migration (ca. 800 km). These differences were consistent between years. Stopover duration was not significantly affected by the age of the bird. However, there was a significant reduction as migration advanced. Our results suggest that spoonbills develop different stopover strategies depending on the expected distance to the wintering grounds. Adults, especially long-distance migratory ones, could reduce the potential negative effects of density-dependence processes by avoiding stopover at the end of the migration period. These findings are of significant relevance for understanding differences in migratory behaviour within single populations, especially for declining waterbirds, as well as stress the relevance of preserving stopover localities for the conservation of intraspecific diversity in migratory species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Many animals undertake seasonal movements from the breeding to the wintering grounds, which are regarded primarily as an adaptation for exploiting the best conditions in food availability throughout the year (Alerstam et al. 2003; Newton 2008). Long-distance migration is a costly process that consists in sequential movements interrupted by stopovers for resting and/or refuelling, and is widespread in many animal groups such as mammals, birds, fish and insects (Alerstam et al. 2003).
Migrating birds spend most of the time at stopover sites (Schaub et al. 2001), which are usually discrete and far apart for many long-distance migratory species, especially waterbirds (Pennycuick and Battley 2003; Battley et al. 2005; Gill et al. 2005; Piersma et al. 2005). Stopover duration is the most important factor determining the total length of a migratory journey and the total amount of energy load (Alerstam et al. 2003). Considering the energy-optimisation theory, individuals should stay in a stopover until enough fuel is stored to reach the next stopover site, independently of the rate of fuel deposition (Alerstam and Lindström 1990; Hedenström and Alerstam 1997). On the other hand, in the context of the time-minimisation theory, individuals should be sensitive to the rate of fuel deposition, adjusting their departure fuel load to their fuel deposition rate (Alerstam and Lindström 1990; Hedenström and Alerstam 1997). In terms of energy turnover rate, associated costs may appear in the short and/or long term (Schmidt-Wellenburg 2007). Therefore, timing and conditions experienced at stopover sites may affect not only the success of the migration, but also the future individual reproduction and survival, with potential consequences for population dynamics (Newton 2006).
During migration, many bird species or populations with similar migratory patterns and/or trophic requirements may use the same stopover sites to refuel and/or rest (Newton 2004). Density-dependent processes may thus be promoted at stopovers, and interspecific and/or intraspecific competition could increase (Alerstam and Hedenström 1998; Newton 2006). In this context, early arrival should be under high positive selection since feeding resources may be fewer, or even depleted, when later birds arrive at a single stopover (Newton 2006; Nolet et al. 2006). Therefore, experience accumulated along years/migrations could play a significant role to achieve an optimal migration (Hedenström 2008) and allows the development of different stopover strategies within a population (Scheiffarth et al. 2002; Drent et al. 2003). For long-lived migratory birds, such as cranes and storks, experience (i.e., age of the birds) could be a sign of consistent migration success (Berthold 2001). Accumulated experience may be crucial to achieve a ‘dominant competitive status’ to be able to outcompete conespecifics at stopover and/or wintering localities (Alerstam et al. 2003), especially in species without apparent sexual dimorphism. Experienced birds should be able to reach required fuel loads faster than inexperienced ones, thus reducing stopover duration (Newton 2008). Nevertheless, to date, empirical data about ecological factors affecting stopover duration are scarce and most frequently involve a reduced number of individuals (Schaub et al. 2001; Salewski and Schaub 2007).
Decisions on the necessity for stopover and its duration may depend on the expected length of the migratory bout (Schaub et al. 2001), but to the best of our knowledge no empirical studies have corroborated this hypothesis. A previous study addressed this question, but looking at two different subspecies of the Northern wheatear Oenanthe oenanthe (Dierschke and Delingat 2001) and potentially including some confounding factors (i.e., differential ecological/energetic requirements between subspecies). Therefore, the remaining distance to the wintering area has not been fully considered yet as a main factor explaining stopover duration in birds. In this study, we examined the relationship between the length of stay at a major stopover site in Northern Iberia and the overwintering area (Africa or South Iberia) in migrating Eurasian spoonbills (Platalea l. leucorodia). We predicted that adult spoonbills travelling to overwinter in Africa will stopover for longer in North Iberia than birds overwintering in South Iberia, and that experience (age) of the birds will play a significant role in explaining stopover duration.
Methods
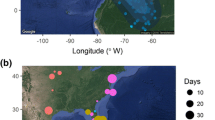
The Eurasian spoonbill (Platalea l. leucorodia) maintains a small breeding population in Western Europe (4,800 pairs; Triplet et al. 2008), catalogued as “Rare” (BirdLife 2004). Fewer than 2,000 pairs breed in northern latitudes, the so-called North-Atlantic population, most (1,600 pairs) in the Netherlands (Triplet et al. 2008; see Fig. 1). These birds spend the winter in two distant geographic areas: about 20% of the birds in the Iberian Peninsula, most of them in the Southwest (Lorenzo and De le Court 2007), and the rest of the population in Northwestern African wetlands, mainly in Mauritania and Senegal (Bauchau et al. 1998; Fig. 1). Around 35–40% of the North-Atlantic spoonbill population makes a strategic stopover in the Santoña Marshes Natural Park (43°30′N 3°30′W; Fig. 1) during the autumn migration (Navedo 2006a), where ca. 80% of these birds came from the Wadden Sea islands, 15% from the inland Netherlands and 5% from France (Navedo 2006a). On average, 90% of the spoonbills using Santoña make their stopover during September (Navedo 2006a).

Main breeding grounds and wintering areas of the North-Atlantic population of the Eurasian spoonbill (Platalea l. leucorodia), depicting the Santoña Marshes Natural Park
Santoña marshes is an estuarine area located in the northern coast of the Iberian Peninsula with semidiurnal tides exposing the intertidal zone (roughly 1,200 ha) daily for 5.5–6.5 h (Navedo and Masero 2007). This estuary is strategically located within the route of the North-Atlantic spoonbill population (Fig. 1), just before the geographical barrier of the Cantabrian Mountains, and about 800 km away from the next main suitable stopover areas at the southern coasts of Iberia (Algarve, Odiel and Doñana). Spoonbill habitat use is invariably driven by the tides, feeding in the intertidal channels during the low tide period and resting at high tide in a supratidal area (Navedo 2006a). Tidal amplitude at Santoña ranges from ca. 2.5 m during neap tides up to ca. 4.5 m during spring ones (mean 3.5 m; Navedo 2006b), so it potentially has a great influence in available foraging possibilities, with periods of spring tides significantly enlarging the potential spatial and time foraging budget.
We have estimated stopover duration of spoonbills in Santoña during September in four consecutive years (2002–2005) on the base of PVC-ringed birds. Every day from 1 to 28 September, and throughout the diurnal low tide period, two experienced observers carefully checked foraging flocks from a boat looking for PVC-ringed birds. Observers were connected by walkie-talkies with people on two little hills overlooking the estuarine area, which informed them about the localisation of all foraging spoonbills in the intertidal channels. Thus, we assumed that almost all the spoonbills using the estuary (mean daily number at low tide was 69.4 birds; J.G. Navedo unpublished data) were checked on a daily basis. When a PVC-ringed bird was located, the boat was slightly beached to identify the code of the PVC ring with the help of a 20×–60× telescope (mean observation distance: 120 m). Approximately 15% of the North-Atlantic spoonbill population is currently marked with individual PVC ring codes (O. Overdijk, unpublished data).
Minimum stopover duration was defined as the time elapsed between the first location and last relocation of a bird in the estuary. Since resighting probability of birds that stopover 3 days or more (n = 74) was 0.89 ± 0.01, and it was not significantly different between years (F 3,70 = 0.81; P = 0.5), it approaches real stopover duration. Overall, we read 974 PVC rings during the study (277, 209, 312 and 176 in 2002, 2003, 2004 and 2005, respectively), comprising 263 accurately identified PVC-ringed spoonbills [69 yearlings (1st year-cal); 39 immatures (2nd and 3rd year-cal) and 155 adult birds (4th year-cal or more)]. Of these birds, 38 were located during more than one autumn at Santoña, corresponding to 34 adults and 4 immatures, all of them 3rd year-cal at the time of their first observation (Dutch Spoonbill Working Group Database). Although most PVC-ringed yearlings were not resighted during the first wintering season, many PVC-ringed birds identified in Santoña were resighted in their major wintering areas a few months later (data obtained from the Dutch Spoonbill Working Group database). Nonetheless, as this population shows high fidelity to wintering area (De Voogd 2004), data were used even if the wintering area was not known for the specific year for which stopover duration was recorded. In these cases, only birds for which wintering area is known, and that have remained constant for 3 years during 2002–2008, were included in the analyses. Under these premises, we analysed information about stopover duration and wintering area for a total of 127 birds (17 yearlings, 17 immatures and 93 adults).
Spoonbills resighted during November and December at Algarve (Portugal), Odiel and Doñana (Spain) were grouped into the Iberian-wintering class (43% of the PVC-ringed birds), whereas spoonbills resighted at that time at Banc d′Arguin (Mauritania) and Senegal Delta (Senegal) were grouped into the African-wintering class (57% of the PVC-ringed birds).
Statistical analysis
Some spoonbills (n = 38) were resighted on more than one migration at Santoña. Therefore, effects of year and wintering area on minimum stopover duration of spoonbills were analysed by fitting a linear mixed model with individual (ring code) as random effect. Wintering area and year were respectively included into the model as two-level (Iberia/Africa) and four-level (2002/2003/2004/2005) fixed factors, respectively. Arrival date, defined as days elapsed from 31 August, was introduced as a potential predictive covariable since timing could significantly affect migratory behaviour (Newton 2008). Similarly, age as well as within-site experience (i.e., number of previous years registered at the area during the study period) were both included as potential predictive variables into the model. Tidal amplitude at arrival, defined as a coefficient ranging from 20 to 120, a measure directly correlated with the height of the tide, was also examined as a predictive covariable due to its potential influence in migratory decisions (Piersma et al. 1990). As arrival date and tidal amplitude were correlated, we have tested their effects separately (not in the same model). However, tidal amplitude was not finally included into the model since it was not correlated with the dependent variable (Spearman r = −0.06; P > 0.05).
Statistical analyses were performed using STATISTICA v6.1 (StatSoft Inc. 2002). In all cases we used a 0.05 level of significance. Values are presented as means ± SE.
Results
Individuals wintering in Africa spent more time (2.7 ± 0.4 days; n = 72) at Santoña marshes during the autumn migration than those wintering in south Iberia (1.5 ± 0.2 days; n = 55) (Table 1; Fig. 2). Minimum stopover duration was similar between years, and longer stays for birds wintering in Africa were always recorded even during the year with the shortest length of stay (2004, Fig. 2). As the migration period progressed, minimum stopover duration decreased significantly (Table 1; Fig. 3). However, neither age nor within-site experience had a significant effect in explaining stopover duration of spoonbills.

Minimum stopover duration (days ± SE) of Eurasian spoonbills (Platalea l. leucorodia) during the autumn migration at Santoña marshes in function of wintering area (Iberia and Africa) and year (2002–2005)

Relationships between minimum stopover duration (days ± SE) of Eurasian spoonbills (Platalea leucorodia) during the autumn migration at Santoña marshes and arrival date (days elapsed since 31 August). Solid circles = birds wintering in Africa; open triangles = birds wintering in Iberia. Note that a given point in the graphic could represent more than one bird (especially for minimum stopover duration = 1)
Discussion
This study empirically shows that stopover duration in a long-distance migratory bird like the Eurasian spoonbill was influenced by the remaining flight distance to the overwintering area. According to our prediction, spoonbills wintering in Africa, and consequently facing a long-distance travel from a crucial stopover site, stay for longer than those birds wintering in south Iberia that performed a shorter migration. Although there are alternative reasons to stay at a stopover site than refuelling (e.g., resting, foraging for maintenance or waiting for favourable winds to continue migration, e.g., Weber et al. 1998), spoonbills were foraging most of the available time during stopover (J.G. Navedo personal observation). Therefore, longer stopover duration would reflect higher energy loads for spoonbills staying at the study area.
Birds wintering in south Iberia seem to minimise the time spent at the stopover locality, flying after a short refuelling. Without large energy requirements, the need to arrive to the best foraging grounds in these short-distance wintering areas ahead of other conspecifics may be the driving force explaining the fast stopover of Iberian-wintering spoonbills. Moreover, as birds wintering in Southern Iberia are close to their destination, they could be less dependent on the foraging conditions at this stopover to successfully complete their migration. Inversely, birds wintering in Africa need to adjust their stopover duration at the Santoña Marshes to the requirements of a high-energy-demanding flight and the prospects for a longer migratory trip (Alerstam and Lindström 1990), thus refuelling for longer. Furthermore, at least some birds might take advantage of the high energy load to ‘skip’ those south Iberian wetlands (Piersma 1987), where their conspecifics overwinter, to perform a non-stop bout from Santoña to north western African wetlands.
Eurasian spoonbills consistently stopover for less time as migration period progresses. Prey consumption at stopover areas would reduce food availability for migratory birds and even deplete them (Gudmunsson et al. 1991; Prop et al. 2003; Nolet et al. 2006). Although we have not measured food availability in our study area (main prey for spoonbills are little fishes and shrimps; J.G. Navedo unpublished data), a significant decrease of food resources for waterbirds occurs in estuaries around north Iberian coasts during autumn migration, and extending to the winter season (Boileau et al. 2001; Viegas et al. 2007). If food is limited, an increase in intraspecific competition resulting from a particularly good breeding season (i.e., high number of breeding pairs and productivity) may have measurable consequences in the migratory performance within a given population (Alerstam et al. 2003), specially for experienced adults (Berthold 2001). The reduced stopover duration observed during 2004 could have been affected by the highest population size during the study period reached in that year (a high number of breeding pairs achieving a high reproductive success; O. Overdijk unpublished data), which also coincided with the maximum number of spoonbills that made a stopover at the Santoña marshes during the study period (Navedo 2006a).
Age-related differences in timing of migration at stopover sites are common in many bird species (e.g. Restani 2000; Ueta and Higuchi 2002; Hake et al. 2003; Meissner 2007; Lee et al. 2008). In our case, a preliminary study addressed the fact that the majority of adult spoonbills (ca. 70%) had currently used the area before the arrival of the majority of yearlings (ca. 60%) at the last week of September (Navedo 2008), when overall stopover duration had markedly decreased. Therefore, yearlings that stopover at the last part of the migration period without knowledge about the remaining distance to wintering areas, associated energy demands and/or suitability of stopover areas may reduce their probability of surviving during a long migration (Newton 2006), which could be viewed as an evolutionary issue regulating long-distance migratory bird populations (Schekkerman et al. 2003). Inversely, adults could minimise the effects derived from density-dependent processes by avoiding stopover at the end of the migration period, in particular African wintering birds dealing with a fairly narrow time window to satisfy their higher energy demands (Skagen and Knopf 1994), that overall arrive a few days earlier (mode: 13 September) than Iberian ones (mode: 17 September).
Notwithstanding the lack of prey surveys and fuelling rates (Hedenström and Alerstam 1997), our findings suggest that spoonbills may experience a high intraspecific competition during stopover that could result in the different strategies observed (Drent et al. 2003). However, further studies are needed to clarify this point and to evaluate potential differences between sexes (Cristol et al. 1999), as well as the role of experience (age) within sexes in these long-lived birds (Berthold 2001), especially for African-wintering spoonbills that refuel for longer.
To sum up, this study provides empirical evidence of different stopover durations in a natural bird population in the function of the remaining migratory distance to overwintering area and remarks the relevance of stopover places for determining migratory route and pace in long-distance migratory birds.
Zusammenfassung
Langstreckenzieher rasten länger: eine Fallstudie an in Nordiberien rastenden Löfflern
Langstreckenzug ist bei Vögeln weit verbreitet und verbindet die Brut- und Überwinterungsgebiete durch eine Serie von Rastplätzen, wo Individuen auftanken und/oder ruhen. Das Ausmaß der Rast ist entscheidend für die Zugstrategie eines Vogels. Hier haben wir die Beziehung zwischen der minimalen Aufenthaltsdauer von PVC-beringten Löfflern (Platalea l. leucorodia) an einem Hauptrastplatz und der verbleibenden Flugentfernung zum Überwinterungsgebiet während vier aufeinander folgender Herbstzüge untersucht. Wir haben auch den potenziellen Einfluss von Timing (Ankunftsdatum) und die Rolle von Erfahrung analysiert, um die Rastdauer von Löfflern zu erklären. Insgesamt bleiben Vögel, die in Afrika überwintern und vom Rastplatz aus eine längere Strecke ziehen (ca. 3000 km), länger (2.7 ± 0.4 Tage) als Vögel, die in Iberien überwintern und eine deutlich kürzere Strecke (ca. 800 km) ziehen (1.5 ± 0.2 Tage). Diese Unterschiede waren über den untersuchten Zeitraum beständig. Die Rastdauer wurde nicht signifikant vom Alter des Vogels beeinflusst. Sie verringerte sich jedoch signifikant mit dem Fortschreiten des Zuges. Unsere Ergebnisse deuten darauf hin, dass Löffler unterschiedliche Raststrategien entwickeln, abhängig von der erwarteten Entfernung zum Überwinterungsgebiet. Altvögel, insbesondere solche, die lange Strecken ziehen, könnten mögliche negative Effekte dichteabhängiger Prozesse reduzieren, indem sie Rasten am Ende des Zuges vermeiden. Diese Ergebnisse sind von signifikanter Bedeutung für das Verstehen von Unterschieden im Zugverhalten innerhalb einzelner Populationen, insbesondere für in ihrem Bestand abnehmende Wasservögel, und betonen die Bedeutung des Schutzes von Rastplätzen für die Bewahrung innerartlicher Diversität bei Zugvögeln.
References
Alerstam T, Hedenström A (1998) The development of bird migration theory. J Avian Biol 29:343–369
Alerstam T, Lindström A (1990) Optimal bird migration: the relative importance of time, energy and safety. In: Gwinner E (ed) Bird migration: physiology and ecophysiology. Springer, Berlin, pp 331–351
Alerstam T, Hedenström A, Åkesson S (2003) Long-distance migration: evolution and determinants. Oikos 103:247–260
Battley PF, Rogers DI, van Gils JA, Piersma T, Hassell CJ, Boyle A, Yang HY (2005) How do red knots Calidris canutus leave Northwest Australia in May and reach the breeding grounds in June? predictions of stopover times, fuelling rates and prey quality in the Yellow Sea. J Avian Biol 36:494–500
Bauchau V, Horn H, Overdijk O (1998) Survival of spoonbills on Wadden Sea islands. J Avian Biol 29:177–182
Berthold P (2001) Bird migration. University Press, Oxford
BirdLife (2004) Birds in Europe: population estimates, trends and conservation status. BirdLife Conservation Series 12. BirdLife International, Oxford
Boileau N, Caupenne M, Le Campion T (2001) Ecologie alimentaire de la spatule blanche Platalea leucorodia en migration postnuptiale dans le marais de Brouage (Charente-Maritime). Ann Soc Sci Nat Charente-Maritime 9:207–218
Cristol DA, Baker MB, Carbone C (1999) Differential migration revisited: latitudinal segregation by age and sex-class. In: Nolan V Jr, Ketterson ED, Thompson CF (eds) Current ornithol 15. Kluwer Academic/Plenum Publishers, New York, pp 33–38
De Voogd M (2004) Methods to sex juvenile Spoonbills and Sex-specific migration in Spoonbills. Master Thesis. Rijksuniversiteit Groningen
Dierschke V, Delingat J (2001) Stopover behaviour and departure decision of northern wheatears, Oenanthe oenanthe, facing different onward non-stop flight distances. Behav Ecol Sociobiol 50:535–545
Drent R, Both C, Green M, Madsen J, Piersma T (2003) Pay-offs and penalties of competing migratory schedules. Oikos 103:274–292
Gill RE, Piersma T, Hufford G, Servranckx R, Riegen A (2005) Crossing the ultimate ecological barrier: evidence for a 11000-km-long nonstop flight form Alaska to New Zealand and eastern Australia by Bar-tailed Godwits. Condor 107:1–20
Gudmunsson GA, Lindström A, Alerstam T (1991) Optimal fat loads and long-distance flights by migrating Knots Calidris canutus, Sanderlings C. alba and Turnstones Arenaria interpres. Ibis 133:140–152
Hake M, Kjellén N, Alerstam T (2003) Age-dependent migration strategy in honey buzzards Pernis apivorus tracked by satellite. Oikos 103:385–396
Hedenström A (2008) Adaptations to migration in birds: behavioural strategies, morphology and scaling effects. Philos Trans R Soc B-Biol Sci 363:287–299
Hedenström A, Alerstam T (1997) Optimum fuel loads in migratory birds: distinguishing between time and energy minimization. J Theor Biol 189:227–234
Lee DL, Black JM, Moore JE, Sedinger JS (2008) Age-specific stopover ecology of Black Brant at Humboldt Bay, California. Ecol 119:9–22
Lorenzo M, De le Court C (2007) Spoonbill winter population in the Iberian Peninsula and the Balearic islands. In: 31st Annual Meeting of the Waterbird Society. Universitat de Barcelona, Barcelona, p. 176
Meissner W (2007) Stopover strategy of adult and juvenile Red Knots Calidris c. canutus in the Puck Bay, southern Baltic. Ardea 95:97–104
Navedo JG (2006a) Identifying stopover wetlands for the conservation of an endangered waterbird species: the role of Santoña Marshes for the Spoonbill Platalea leucorodia during autumn migration. In: Triplet P, Overdijk O (eds) EUROSITE Spoonbill Working Group Newsletter, Vol. 4. pp. 48–51
Navedo JG (2006b) Efectos del marisqueo tradicional sobre la alimentación de las aves limícolas en áreas intermareales: aportaciones para la gestión de los estuarios cantábricos. Ph.D. Thesis, Universidad de Cantabria, p. 165
Navedo JG (2008) Different stopover timing in juvenile and adult spoonbills during southward migration. In: Triplet P, Overdijk O, Smart, M (eds) EUROSITE Spoonbill Working Group Newsletter, Vol. 6. pp. 2–5
Navedo JG, Masero JA (2007) Measuring potential negative effects of traditional harvesting practices on waterbirds: a case study with migrating curlews. Anim Cons 10:88–94
Newton I (2004) Population limitation in migrants. Ibis 146:197–226
Newton I (2006) Can conditions experienced during migration limit the population levels of birds? J Ornithol 147:146–166
Newton I (2008) The migration ecology of birds. Academic Press, London
Nolet BA, Gyimesi A, Klaassen RHG (2006) Prediction of bird-day carrying capacity on a staging site: a test of depletion models. J Anim Ecol 75:1285–1292
Pennycuick CJ, Battley PF (2003) Burning the engine: a time-marching computation of fat and protein consumption in a 5420-km non-stop flight by great knots, Calidris tenuirostris. Oikos 103:323–332
Piersma T (1987) Hop, skip or jump? constraints on migration of arctic waders by feeding, fattening and flight speed. Limosa 60:185–194 (In Dutch with English summary)
Piersma T, Zwarts L, Bruggemann JH (1990) Behavioural aspects of the departure of waders before long-distance flights: flocking, vocalizations, flight paths and diurnal timing. Ardea 78:157–184
Piersma T, Rodgers DI, González PM, Zwarts L, Niles LJ, do Nascimento ILS, Minton CDT, Baker AJ (2005) Fuel storage rates before northward flights in Red Knots worldwide: facing the severest constraint in tropical intertidal environments? In: Greenberg R, Marra PP (eds) Birds of two worlds: the ecology and evolution of migration. John Hopkins University Press, Baltimore, pp 262–373
Prop J, Black JM, Shimmings P (2003) Travel schedules to the high arctic: barnacle geese trade-off the timing of migration with accumulation of fat deposits. Oikos 103:403–414
Restani M (2000) Age-specific stopover behavior of migrant bald eagles. Wilson Bull 112:28–34
Salewski V, Schaub M (2007) Stopover duration of palearctic passerine migrants in the western Sahara—independent of fat stores? Ibis 149:223–236
Schaub M, Pradel R, Jenni L, Lebreton J-D (2001) Migrating birds stop over longer than usually thought: an improved capture-recapture analysis. Ecol 82:852–859
Scheiffarth G, Wahls S, Ketzenberg C, Exo KM (2002) Spring migration strategies of two populations of bar-tailed godwits, Limosa lapponica, in the Wadden Sea: time minimizers or energy minimizers? Oikos 96:346–354
Schekkerman H, Tulp I, Ens B (2003) Conservation of long-distance migratory wader populations: reproductive consequences of events occurring in distant staging sites. Wader Study Group Bull 100(Special issue):151–156
Schmidt-Wellenburg CA (2007) Costs of migration: short- and long-term consequences of avian endurance flight. Ph.D. Thesis, University of Groningen, p. 159
Skagen SK, Knopf FL (1994) Migrating shorebirds and habitat dynamics at a prairie wetland complex. Wilson Bull 106:91–105
Statsoft Inc. (2002) STATISTICA. http://www.statsoft.com
Triplet P, Overdijk O, Smart M, Nagy S, Schneider-Jacoby M, Karauz ES, Pigniczki Cs, Baha El Din S, Kralj J, Sandor A, Navedo JG (Compilers) (2008) International single species action plan for the conservation of the eurasian spoonbill Platalea leucorodia. AEWA Technical Series no 35. Bonn, Germany, p. 40 (plus annexes)
Ueta M, Higuchi H (2002) Difference in migration pattern between adult and immature birds using satellites. Auk 119:832–835
Viegas I, Martinho F, Neto J, Pardal M (2007) Population dynamics, distribution and secondary production of the brown shrimp Crangon crangon (L.) in a southern European estuary. Latitudinal variations. Sci Mar 71:451–460
Weber T, Alerstam T, Hedenström A (1998) Stopover decisions under wind influence. J Avian Biol 29:552–560
Acknowledgments
We thank Alejandro García-Herrera, Álvaro Bustamante, Máximo Sánchez-Cobo, Manuel A. Alcántara, Juan José Aja, Virginia Iturriaga and Ana Casero in addition to all the volunteers from the Spoonbill Migration Monitoring Program (SEO/BirdLife) for field assistance. Emma Dahl, Fernando Arce, Joaquín Bedia, Dra. Auxiliadora Villegas, Dr. Manuel Mota, Dr. Manuel Medina and an anonymous reviewer, helped to improve an earlier version of this manuscript. The Spanish Ornithological Society (SEO/BirdLife) and the Spanish National Parks Service provided us with logistic and financial support during the field work. JGN was currently supported by a postdoctoral fellowship of Plan Regional de Investigación (III PRI+D+I) of Junta de Extremadura.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H. Mouritsen.
Rights and permissions
About this article
Cite this article
Navedo, J.G., Orizaola, G., Masero, J.A. et al. Long-distance travellers stopover for longer: a case study with spoonbills staying in North Iberia. J Ornithol 151, 915–921 (2010). https://doi.org/10.1007/s10336-010-0530-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-010-0530-z