
Abstract
Genetic studies have shown that extra-pair paternity is widespread among socially monogamous bird species. Yet, the role of males and females and their behavior leading to this mixed reproductive strategy is poorly understood. Here, we analyze paternity in relation to male age and mating behavior in the socially monogamous reed bunting (Emberiza schoeniclus). We report a positive relation between male extraterritorial forays and success in obtaining extra-pair fertilizations. Extraterritorial forays tended to increase in frequency with male age and older males sired a larger number of extra-pair offspring than young males. Identified extra-pair sires were old in nine out of ten cases. The likelihood of being cuckolded was not affected by male age. Although based on correlative data, our results highlight age-dependent explorative male behavior as a key determinant for the understanding of extra-pair mating in the reed bunting. We do, however, emphasize the need for further studies to reveal the role of females in extra-pair copulations and fertilizations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Extra-pair mating is a common mating strategy among socially monogamous birds, as it occurs in almost 90% of the species studied so far (reviewed in Griffith et al. 2002). The benefit to males of this mixed reproductive strategy is obvious, as males have the potential of siring a larger number of offspring through extra-pair copulations. Why females engage in extra-pair mating is less clear and debated, but it is generally assumed that they obtain indirect (genetic) benefits by improving the quality of their progeny (cf. Jennions and Petrie 2000). Although it is acknowledged that extra-pair mating involves three main players (the female, her social partner and the extra-pair male) (Lifjeld et al. 1994; Westneat and Stewart 2003), their role in this game is still poorly understood and it has been emphasized a need for additional behavioral studies to supplement the far more frequent genetic studies (Westneat and Stewart 2003).
Previous studies on the behavior of the two sexes during the mating period have revealed that extraterritorial forays occur rarely in females, but are more frequently observed in males (Westneat and Stewart 2003). This suggests a more active role of males in seeking extra-pair copulations. Furthermore, male age seems to be an important factor in extra-pair mating in many species as older males are more successful in acquiring extra-pair fertilizations than young males (reviewed in Griffith et al.. 2002) and extra-pair males are older than the males they are cuckolding (e.g., Weatherhead and Boag 1995; Yezerinac and Weatherhead 1997; Johnsen et al. 2001). These patterns may be explained by an increased ability of males to seek and successfully obtain extra-pair copulations as they get older and more experienced (Weatherhead and Boag 1995). However, it needs to be emphasized that the patterns might also be interpreted as a female mating preference for older and more viable males (cf. Brooks and Kemp 2001).
The aim of this study was to combine a behavioral and a molecular approach to study patterns of extra-pair mating in the reed bunting (Emberiza schoeniclus). We examined whether male exploratory behavior (i.e., extraterritorial forays) had an impact on extra-pair fertilization success and whether this behavior was affected by male age. Furthermore, we investigated the effects of male age on within-pair and extra-pair fertilization success.
Methods
Study species
The reed bunting is a small (adult weight≈18 g), migratory, insectivorous passerine. Males are territorial and pair up with a single female. Reed buntings nest on or near the ground and females lay three to six eggs. Males may assist females in incubation, while both parents provision the young (Cramp and Perrins 1994). Although socially monogamous, reed buntings show high levels of extra-pair paternity (Dixon et al. 1994; Bouwman et al. 2005; Kleven and Lifjeld 2005). Male reed buntings perform mate-guarding and the intensity is positively related to the proportion of young sired in their own brood (Marthinsen et al. 2005).
Study area and fieldwork
We studied a breeding population of reed buntings in the years 2001 and 2002 in a subalpine region of Southern Norway (61°25’N, 8°52’E). See Vik (1978) for a detailed description of the vegetation in the area. Here, we use a subsample of the data collected (i.e., data from 2002), due to the difficulties in determining age of adult reed buntings. The individuals were classified as either young or old, based on whether they had been banded as adults the previous breeding season (in line with, e.g., Yezerinac and Weatherhead 1997). Data on wing length provided support for this method of ageing the individuals. A pairwise comparison of males measured both years showed a significant increase in wing length (t 6=3.4, P=0.015) and, although the sample size was low, males classified as old tended to have longer wings than those classified as young (t-test, t 33=1.8, P=0.076). We emphasize that, through the use of this age-classification method, some old individuals may have been included in the category of young males. However, we consider this to be a conservative approach if differences between the two categories of individuals were to be detected. Adults were captured with mist nets, bled and banded with a unique code including a silver band and three plastic colour bands. A small sample of blood was obtained from the nestlings. Further details about the field methods are presented elsewhere (Kleven and Lifjeld 2004).
Extraterritorial forays
Eighteen males (eight young and ten old) were observed in 2002 from 15 days before until 5 days after the onset of egg laying (for further details, see Marthinsen et al. 2005). A total of 73 observation sessions (4.1±0.53 SE observation sessions per pair; range 1–8) were obtained and each session lasted 12–22 min (16.9±0.40 SE). In our study area, the average distance between nearest neighbors was 135 m (±8 SE, n=94 nests, data from both years). We defined a territory as the area around the nest, extending halfway between adjacent neighbor nests. Extraterritorial forays were defined as events in which the male left his territory, not for the purpose of following (mate-guarding) his social mate.
Paternity analyses
We determined paternity with nine polymorphic microsatellite markers. The details of the genotyping methods have been presented elsewhere (Kleven and Lifjeld 2004), hence they are only briefly described here. The blood samples were diluted and stored in lysis buffer and DNA was extracted using QIAamp DNA mini kit (Qiagen). The DNA fragments were amplified with polymerase chain reaction (PCR) and the PCR products were sized on an ABI 310 sequencer (Applied Biosystems) and analyzed with the software program GeneMapper Version 2.0 (Applied Biosystems). The combined exclusion probability (Jamieson 1994) of the nine markers was high, viz. >0.999 with one parent known.
Statistical analyses
Prior to the analyses, we examined our data for normality and homogeneity of variances, with the Shapiro-Wilk’s W-test and Brown and Forsythe’s test, respectively. Parametric methods were used when the conditions for these tests were met, otherwise non-parametric methods were used. The proportion of within-pair paternity, the likelihood of losing paternity, the likelihood of gaining extra-pair paternity (binomial distributions) and the total fertilization success (Poisson distribution) were analyzed with generalized linear models (GLMs). We used GLMStat Version 5.5 and STATISTICA Version 6.1 to analyse our data. Mean values are given ± one standard error. All tests were two-tailed and the level of significance was set to P=0.05.
Results
Extra-pair paternity was found in 64% (23/36) of the broods and 36% (66/181) of the offspring were sired by an extra-pair male. We identified eight extra-pair males in ten of the 23 broods. These males sired 50% (33/66) of the extra-pair offspring. Males that obtained extra-pair fertilizations sired more offspring in total compared to males who failed to do so (6.9±1.0, n=8 and 3.3±0.4, n=28; respectively; GLM with Poisson error distribution and logarithm link; χ 2=19.2, P<0.001).
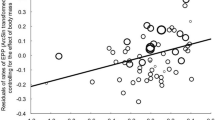
The average rate of extraterritorial forays amounted to 1.1 (±0.5 forays/h, n=18 males). Males increased their likelihood of obtaining extra-pair fertilizations through extraterritorial forays (GLM with binomial error distribution and logit link; χ 2=7.0, n=18 males, P=0.008). The rate of extraterritorial forays also correlated positively with the number of extra-pair offspring a male sired (Spearman rank correlation; r s=0.61, n=18 males, P=0.007; Fig. 1). Old males left their territories and social mate 7 times as frequently during mate-guarding observations compared with young males (1.75±0.76 forays, n=10 males, and 0.25±0.25 forays, n=8 males, respectively; Mann-Whitney U-test; Z=1.7, P=0.08). Consequently, the likelihood of obtaining extra-pair fertilizations and the number of extra-pair young sired were higher for old than for young males (Fig. 2; Table 1). In the ten cases where we knew the identity of the extra-pair male, the cuckolders were old in nine of them, and the males they cuckolded were both young (n=4) and old (n=5). In the single case where the cuckolder was young, the male that was cuckolded was also young.

Extra-pair fertilization success in relation to extraterritorial foray rate in young (open circles) and old (closed circles) male reed buntings (Emberiza schoeniclus). The size of the circles indicates number of data points (open circle and filled circle=1 male, filled circle=4 males, open circle=7 males). A regression line is indicated for the two age-categories combined

Male age and extra-pair fertilization success in reed buntings. Numbers indicate sample sizes of males and vertical lines indicate +1 standard error. The sample of males also includes those that failed in their own breeding attempt, due to, e.g., predation
The social male was young in 20 and old in 16 of the pairs. Young males did not have a significantly higher probability of losing paternity or to sire a lower proportion of young in their own brood compared to old males (Table 1). Also, the proportion of within-pair offspring was similar among broods for males in which we had data on paternity from both years (Wilcoxon’s matched pairs test; Z=0.7, n=11, P=0.5).
On average, the total fertilization success for old males (4.7 young±0.8, n=16) was more than 30% higher than young males (3.6±0.4, n=20), but the difference was not statistically significant (Table 1).
Discussion
Here we show that male reed buntings performed extraterritorial forays, but that there existed considerable variation among males in the frequency of such forays. Extraterritorial forays predicted extra-pair fertilization success in this socially monogamous species. Furthermore, the rate of male extraterritorial forays tended to increase with age and old males sired more extra-pair offspring than young males. There was, however, no difference between old and young males in their success of securing within-pair paternity.
Extra-pair paternity is the outcome of a series of events, that is, encounter, extra-pair copulation and, ultimately, fertilization, in which the influential role of each sex may differ considerably. Although the occurrence of extra-pair paternity is well documented in socially monogamous species, the role of males and females in extra-pair mating events remains unclear (Westneat and Stewart 2003). In the first step, both sexes may either advertise for extra-pair partners on territory or more actively seek extra-pair partners outside their own territory. We have found no indication of female reed buntings performing extraterritorial forays (Marthinsen et al. 2005). In general, the evidence of females actively seeking extra-pair copulations is also scarce and limited to a very few cases (reviewed in Westneat and Stewart 2003). This suggests that females play a minor role in seeking extra-pair copulations. Alternatively, female extraterritorial behavior might be covert and thus have gone undetected. Male extraterritorial forays, on the other hand, seem to be common in socially monogamous species as the behavior has been documented in several species (Westneat and Stewart 2003). In many species it has also been shown that extraterritorial forays actually result in extra-pair copulations (Westneat and Stewart 2003). Our finding of a positive relationship in the reed bunting between the rate of male extraterritorial forays and actual extra-pair fertilization success provides additional support to the importance of male explorative behavior in initiating extra-pair copulations and suggest that males play a more important role than females in shaping the patterns of extra-pair mating observed.
The advantage of extra-pair mating may be higher for males than females, since males will increase the number of descendants through a mixed reproductive strategy, while the benefits to females are less clear and might even be negligible (Arnqvist and Kirkpatrick 2005). In the studied reed bunting population, we have previously found that females do not enhance offspring cell-mediated immunity or growth (Kleven and Lifjeld 2004), or increase offspring heterozygosity through extra-pair mating (Kleven and Lifjeld 2005). Presently, we therefore have no empirical evidence supporting the hypothesis that female participation in extra-pair mating is to obtain indirect benefits. Hence, the selection pressure for seeking extra-pair copulations may be stronger in males than in females. Yet, although extra-pair mating seems to be male driven in the reed bunting, females may still benefit. For instance, if the traits associated with explorative and coercive male behavior are heritable, females accepting copulations from intruding extra-pair males may produce sons possessing similar behavioral traits. However, more data are certainly needed before firm conclusions can be drawn.
Male extraterritorial forays tended to increase with age and older males were more successful in siring extra-pair offspring than young males, thus supporting the hypothesis by Weatherhead and Boag (1995). According to their hypothesis, older and more experienced males might have higher success in extra-pair mating due to being better than younger males at exploiting possibilities for extra-pair copulations and coercing females. Our findings also confirm the results of several previous studies (reviewed in Griffith et al. 2002), suggesting that male age is an important factor in extra-pair mating in socially monogamous birds. It has been hypothesized that females should prefer older males due to their proven viability through a long lifespan (Brooks and Kemp 2001; but see, e.g., Hansen and Price 1995). Hence, females may be less reluctant in engaging in extra-pair copulations since the intruders are almost exclusively old males and their extra-pair offspring may inherit genes for longevity.
Previously, we have shown that male mate-guarding has a positive effect on securing within-pair paternity in the reed bunting (Marthinsen et al. 2005). In that same study, we also found that mate-guarding was performed discontinuously and some males probably obtained extra-pair fertilizations around the time when their social female was fertile. Together with the data provided in the present study, male reed buntings seem to improve their fitness both by mate-guarding their social partner and seeking extra-pair copulations off territory. This will most likely result in a trade-off in performing these activities as they are mutually exclusive. Extraterritorial forays seem to be performed mainly by old males and such a trade-off may thus be more favourably solved within this age-category of males. Accordingly, it might be expected that old males should guard their females less intensely than young males, as shown in, e.g., bluethroats (Luscinia svecica) (Johnsen et al. 2003), but that was not found in our previous study on reed buntings (Marthinsen et al. 2005). The sample size (n=12) for the test in that study may have been to low to detect a potential difference in male mate-guarding intensity among age-categories. The intensity and dynamics of male mate-guarding may even be modified in a more intricate pattern, also depending on the age of the female (cf. Dietrich et al. 2004).
If extra-pair copulations and fertilizations were the result of a female preference for older males, we might have expected to find this to be reflected in within-pair paternity. The lack of a difference between old and young males in securing paternity in own broods suggests that female choice for older males play a minor role in explaining the patterns of extra-pair paternity in this population of reed buntings. However, since extra-pair sires were of the same age or older than the males they cuckolded, we cannot exclude the possibility of a female preference for older males as extra-pair sires.
In this study, we identified the sires for only half of all extra-pair young. The genetic fathers of the remaining extra-pair young may have been other territorial males breeding outside our study area or, potentially, non-breeding ‘floater’ males. If the unidentified extra-pair sires were mainly young floater males, making the ‘best-of-a-bad-job’, this would have some bearing on our findings and conclusions. We therefore emphasize that the implications drawn from the results of this study are based on the assumption that unidentified extra-pair sires are territorial males. We consider this assumption to be reasonable since non-breeding males in other socially monogamous species do not seem to succeed very well in obtaining fertilizations (e.g., Whittingham and Dunn 2005; Kempenaers et al. 2001).
In conclusion, the combination of behavioral and molecular data used in this study reveal the significance of male extraterritorial forays and age as important correlates of extra-pair mating success in the socially monogamous reed bunting. Age-related male explorative behavior as a key factor for understanding extra-pair mating may have a broader generality than presently acknowledged.
Zusammenfassung
Extraterritoriale Streifzüge, Alter und Vaterschaft der Männchen bei der sozial monogamen Rohrammer (Emberiza schoeniclus)
Genetische Untersuchungen haben gezeigt, dass paarfremde Vaterschaft unter sozial monogamen Vögeln weit verbreitet ist. Trotzdem ist die Rolle von Männchen und Weibchen und ihrem Verhalten, das zu dieser gemischten Fortpflanzungsstrategie führt, wenig verstanden. Wir untersuchten die Vaterschaft bei der sozial monogamen Rohrammer (E. schoeniclus) in Abhängigkeit von Alter und Paarungsverhalten der Männchen. Dabei fanden wir einen positiven Zusammenhang zwischen extraterritorialen Streifzügen der Männchen und dem Erfolg, eine paarfremde Befruchtung zu erzielen. Mit steigendem Alter der Männchen nahm die Anzahl extraterritorialer Streifzüge zu und ältere Männchen zeugten mehr paarfremden Nachwuchs als jüngere Männchen. Individuell identifizierte paarfremde Väter waren in neun von zehn Fällen alte Männchen. Die Wahrscheinlichkeit, betrogen zu werden, hing nicht mit dem Alter der Männchen zusammen. Obwohl auf Korrelationsdaten beruhend, verdeutlichen unsere Ergebnisse, dass das altersabhängige exploratives Verhalten der Männchen ein Schlüsselfaktor zum Verständnis paarfremder Paarungen bei der Rohrammer ist. Allerdings muss auch die Rolle der Weibchen bei paarfremden Kopulationen und Befruchtungen untersucht werden.
References
Arnqvist G, Kirkpatrick M (2005) The evolution of infidelity in socially monogamous passerines: the strength of direct and indirect selection on extra-pair copulation behavior in females. Am Nat 165:S26–S37
Bouwman KM, Lessells CM, Komdeur J (2005) Male reed buntings do not adjust paternal effort in relation to extra-pair paternity. Behav Ecol 16:499–506
Brooks R, Kemp DJ (2001) Can older males deliver the good genes? Trends Ecol Evol 16:308–313
Cramp S and Perrins CM (1994) The birds of the western Palearctic, vol. 9. Oxford University Press, Oxford
Dietrich V, Schmoll T, Winkel W, Epplen JT, Lubjuhn T (2004) Pair identity—an important factor concerning variation in extra-pair paternity in the coal tit (Parus ater). Behaviour 141:817–835
Dixon A, Ross D, O’Malley SLC, Burke T (1994) Paternal investment inversely related to degree of extra-pair paternity in the reed bunting. Nature 371:698–700
Griffith SC, Owens IPF, Thuman KA (2002) Extra pair paternity in birds: a review of interspecific variation and adaptive function. Mol Ecol 11:2195–2212
Hansen TF, Price DK (1995) Good genes and old age: do old mates provide superior genes? J Evol Biol 8:759–778
Jamieson A (1994) The effectiveness of using co-dominant polymorphic allelic series for (1) checking pedigrees and (2) distinguishing full-sib pair members. Anim Genet 25:37–44
Jennions MD, Petrie M (2000) Why do females mate multiply? A review of the genetic benefits. Biol Rev 75:21–64
Johnsen A, Lifjeld JT, Andersson S, Örnborg J, Amundsen T (2001) Male characteristics and fertilisation success in bluethroats. Behaviour 138:1371–1390
Johnsen A, Lifjeld JT, Krokene C (2003) Age-related variation in mate-guarding intensity in the bluethroat (Luscinia s. svecica). Ethology 109:147–158
Kempenaers B, Everding S, Bishop C, Boag P, Robertson RJ (2001) Extra-pair paternity and the reproductive role of male floaters in the tree swallow (Tachycineta bicolor). Behav Ecol Sociobiol 49:251–259
Kleven O, Lifjeld JT (2004) Extrapair paternity and offspring immunocompetence in the reed bunting, Emberiza schoeniclus. Anim Behav 68:283–289
Kleven O, Lifjeld JT (2005) No evidence for increased offspring heterozygosity from extrapair mating in the reed bunting (Emberiza schoeniclus). Behav Ecol 16:561–565
Lifjeld JT, Dunn PO, Westneat DF (1994) Sexual selection by sperm competition in birds: male-male competition or female choice? J Avian Biol 25:244–250
Marthinsen G, Kleven O, Brenna E, Lifjeld JT (2005) Part-time mate guarding affects paternity in male reed buntings (Emberiza schoeniclus). Ethology 111:397–409
Vik R (1978) The Lake Øvre Heimdalsvatn—a subalpine freshwater ecosystem. Holarctic Ecol 1:84–320
Weatherhead PJ, Boag PT (1995) Pair and extra-pair mating success relative to male quality in red-winged blackbirds. Behav Ecol Sociobiol 37:81–91
Westneat DF, Stewart IRK (2003) Extra-pair paternity in birds: causes, correlates and conflict. Annu Rev Ecol Evol Syst 34:365–396
Whittingham LA, Dunn PO (2005) Effects of extra-pair and within-pair reproductive success on the opportunity for selection in birds. Behav Ecol 16:138–144
Yezerinac SM, Weatherhead PJ (1997) Extra-pair mating, male plumage coloration and sexual selection in yellow warblers (Dendroica petechia). Proc R Soc Lond B 264:527–532
Acknowledgements
We are grateful to Frode Fossøy, Jon B. Kirkebø, Roy Mangersnes, Henrik Pärn, Roar Rismark and Dave Showler for their help and support during the field work. We are especially indebted to Erik Brenna for help both in the field and with the lab work. The study was supported by a grant from the Research Council of Norway and was carried out in accordance with ethical guidelines stated by the Norwegian Animal Research Authority (Licence no. S-1236/01).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bairlein
Rights and permissions
About this article
Cite this article
Kleven, O., Marthinsen, G. & Lifjeld, J.T. Male extraterritorial forays, age and paternity in the socially monogamous reed bunting (Emberiza schoeniclus). J Ornithol 147, 468–473 (2006). https://doi.org/10.1007/s10336-005-0039-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-005-0039-z