
Abstract
In this study, we examine acoustic individuality in male duet songs of wild, non-habituated Bornean southern gibbons (Hylobates albibarbis) and identify contributing acoustic variables. We recorded 174 male duet songs from nine groups in a rainforest in Central Kalimantan, Indonesia. Each male portion of the duet was analysed for 14 acoustic variables at three levels of variation, including six note-specific variables (start frequency, end frequency, minimum frequency, maximum frequency, average frequency and duration), four phrase-specific variables (minimum frequency, maximum frequency, duration and number of syllables) and four song-specific variables (minimum frequency, maximum frequency, duration and number of syllables). Principal component analysis was performed to summarise each of these sets of variables into a total of six principal components (PCs). Strong acoustic individuality was found in all PCs and at all three levels: note, phrase and song (all p < 0.001). Furthermore, a particularly high magnitude of individuality was found in PC 1 of the song-specific analysis, defined by the acoustic variables of duration and number of syllables. Due to the high levels of individuality, we suggest that these acoustic variables may be used by Bornean southern gibbons for individual discrimination. As well as furthering our biological understanding of male gibbon song with regards to individuality and associated conspecific recognition, these findings also have the potential to help improve population survey methods, such as the acoustic sampling method using listening points, by offering a more accurate method of individual recognition.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Vocal communication is an important behavioural phenomenon in primates. A whole spectrum of abilities exists, including the speech of Homo sapiens, the pant hoots of chimpanzees (Mitani et al. 1996) and the long calls of male orangutans (Lameira and Wich 2008). All species of gibbon produce loud, complex and stereotyped vocalisations or ‘songs’ (Marshall and Marshall 1976; Haimoff 1984a; Geissmann 1995; Geissmann and Orgeldinger 2000). Song bouts are usually produced at dawn or in the early morning, last for 10–30 min on average and involve a male singing alone in the form of a solo or a mated pair singing together in the form of a duet, although Kloss’s and Javan gibbons do not duet (Whitten 1982; Dallmann and Geissmann 2009). Whitten (1982) has shown that Kloss’s gibbons sing at times and frequencies which are optimal for sound transmission and hence communication with neighbours. There is much debate regarding the function of gibbon song. It is thought that two main functions exist: (1) territory maintenance and (2) pair-bond maintenance (Chivers 1974). However, many other sex-specific alternative functions have been proposed; For example, male (solo and duet) song may be involved in mate attraction and female duet song in the declaration that the male is already paired (Gittins 1978, 1980, 1982). Mitani (1988) suggests that there is no evidence to support the theory that male song performs an intersexual mate attraction function, although this is likely to be the case for unmated males. Mitani also succinctly summarises the function of gibbon song as the conveyance of information regarding the location and sex of the individual (Mitani 1988).
In this study, the Bornean southern gibbon (Hylobates albibarbis) is used as a model species for study of the level of individuality in male song pattern. This species is endemic to Borneo, and its geographic distribution is restricted to the southwest of the island. The Bornean southern gibbon is light brown in colour, with dark-brown underparts, hands, legs and cap, black fingers and toes and a white brow (Hirai et al. 2009). Bornean southern gibbons produce male solo song bouts which usually occur before dawn and female–male duets which occur post-dawn. Duets are characterised by the female great call, which is preceded and ended by the male duet song.
The individuality of the female great call has been widely studied in the genus Hylobates, perhaps due to its conspicuous nature (Haimoff 1984a, b, 1985, 1986; Raemaekers et al. 1984; Haimoff and Tilson 1985; Dallmann and Geissmann 2001a, b, 2009; Cheyne et al. 2007a; Oyakawa et al. 2007); For example, Dallmann and Geissmann (2001a) found that inter-individual variability of great calls was significantly higher than intra-individual variability in female wild silvery gibbons (H. moloch) on Java, thus proving that individual female gibbon calls are relatively fixed over time. Oyakawa et al. (2007) found that the song of H. agilis females had a high level of acoustic individuality in the climax part of the great call.
Much less is known about the male counterpart. Mitani and Marler (1989) have examined the phonological structure of male song in the agile gibbon (H. agilis) and showed that it is produced by reference to a set of rules which indicate the position and order of elements in the song. There have only been two published papers concerning the individuality of male gibbon song. The first of these was conducted by Mitani (1987), who studied male song variation in gibbons from three different populations: Müller’s gibbons (H. muelleri) in East Kalimantan, and agile gibbons (H. agilis) in South Sumatra and West Kalimantan (the latter population since renamed H. albibarbis; Groves 2001). Mitani found significant inter-individual variation in male gibbon song within populations in all measures. The second was conducted by Geissmann et al. (2005), who carried out a quantitative evaluation of the syntax of the male solo song in the Javan silvery gibbon (H. moloch) and found a high degree of inter-individual variation in the note repertoire of silvery gibbon males. The male song of H. albibarbis has not been analysed since the species was split from H. agilis by Groves (2001) and has never been studied in a mixed peat-swamp forest.
Oyakawa et al. (2007) suggest that individual gibbon song, if used in recognition, may serve the purpose of maintaining relationships between individuals, as well as distinguishing intruders from relatives, hence maintaining both spatial and social organisation. If this is true, there should be strong selection for acoustic individuality in the songs of both sexes.
H. albibarbis is classed as endangered in the IUCN Red List 2012 (IUCN 2012). It is threatened throughout its range by logging, forest conversion and, especially in peat-swamp forests, by fire. Long-term monitoring and assessment of population numbers is essential, as are simple methods for assessing this. Unfortunately, population survey methods are still hampered by the labour- and time-intensive nature of the triangulation method, as well as the error associated with estimating the number of groups heard (Brockelman and Ali 1987; Brockelman and Srikosamatara 1993; Nijman and Menken 2005; Cheyne et al. 2007b; Hamard et al. 2010). New methods which may facilitate population surveys are worth testing, as many areas where H. albibarbis may occur are unsurveyed and outside protected areas. This study proposes a new method for accurately surveying H. albibarbis by recording and analysing the songs of males for specific acoustic features.
Methods
Study site and subject animals
A population of wild Bornean southern gibbons was investigated in a previously unsurveyed rainforest in central Kalimantan, Indonesia (1°12′S, 114°6′E; Fig. 1). The forest contained a whole gradation of forest types from kerangas (heath) forest and peat-swamp forest, to very dense low pole in a relatively small area. Gibbon groups were only found in the former two habitat types. Research took place from July 10 to August 30, 2010. The average daily temperature during this period was 27.5 °C, and the average daily rainfall was 12.3 mm. Preliminary triangulation surveys suggested a total of 20 gibbon groups in the study area; these were mapped and territory sizes estimated (Cheyne et al. 2007b; Hamard et al. 2010). This study focussed on nine groups (A, B, C, D, E, F, G, H, I), all living within 2 km of the field station; however, there were an additional ten groups in the area (J, K, L, M, N, O, P, Q, R, S). All groups were wild and non-habituated.

Location of study site (star) in relation to regional capital, Palangka Raya (circle)
Vocal recording
Vocalisations were recorded between 0400 and 0900 hours, when the gibbons were vocally active. Two or three researchers walked to a particular listening point in the rainforest, where they made recordings of vocal activity using a Zoom H2 Handy Recorder and stand, with a sampling rate of 48 kHz and a 16-bit resolution. The recorder was set to the two-channel setting with a Mic Direction of 90° to the front (“FRONT 90”), fixed to a stand and pointed in the direction of vocal activity. Only groups which were close enough for clear recording (i.e. no more than 500 m away) were recorded. The start time, end time and location of the recording were also noted. The direction of the song was determined using a magnetic compass, and the distance of the group from the listening point was estimated. The type of song was also noted (i.e. duet or non-duet), and the number of great call repeats was recorded for all duet songs. Groups were identified by the location of the vocalisation (territories tend to be well defined) and group composition (number of singing individuals).
Data were collected on a total of 29 days, resulting in a total recording time of 1,856 min. Male songs for which the sound quality was poor or the individual identity was ambiguous were eliminated from the acoustic analysis. A total of 174 male songs were recorded, consisting of approximately 20 (and not <17) songs from each of the nine different groups. Each of the nine subject groups contained one adult male whose duet song was recorded; therefore individual male and group names are used interchangeably in this paper.
Male Bornean southern gibbon song structure
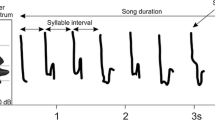
This study focusses on the duet song of the male Bornean southern gibbon and uses the acoustic terminology proposed by Haimoff (1984b). Haimoff (1984b) defines a note as “any single continuous sound of any distinct frequency or frequency modulation, which may be produced during either inhalation or exhalation”. The average note in the song of a male Bornean southern gibbon has a duration of 0.4 s and a frequency range of 391.7 Hz (Table 1). A phrase is defined as “a single vocal activity consisting of a larger or loose collection of notes” (Haimoff 1984b). The duet song of the male Bornean southern gibbon is made up of two distinct phrases: (1) male phrase 1: before the female great call, and (2) male phrase 2: after the female great call (Fig. 2).

An example sonogram showing the position of the phrases in the male duet song of the Bornean southern gibbon
Male phrase 1 is characterised by having an average of 5.9 notes, an average frequency range of 643.0 Hz and an average duration of 3.5 s. Male phrase 2 is characterised by having an average of 3.1 notes, an average frequency range of 372.5 Hz and an average duration of 2.2 s. Therefore, phrase 1 is longer, made up of more syllables and spans a larger frequency range than phrase 2 (Table 1).
Lastly, a song is defined as “a series of notes, generally of more than one type, uttered in succession and so related as to form a recognisable sequence or pattern in time” (Haimoff 1984b) and is a shorter segment of a song bout. The average song of a male Bornean southern gibbon has a duration of 5.7 s and a frequency range of 705.2 Hz and consists of 9.0 notes (Table 1).
Acoustic analysis
General settings
Sonograms of the recorded vocalisations were generated using sound analysis software (Raven Pro 1.3). A view range of approximately 2000 Hz and a window length of approximately 30 s were used (Fig. 3).

Representative sonograms for three subject males (time in seconds on the horizontal axis)
Acoustic variables
To compare the sonograms and calculate the individuality of the male duet song, a total of 14 acoustic variables were selected. This included six note-specific variables (start frequency, end frequency, minimum frequency, maximum frequency, average frequency and duration), four phrase-specific variables (minimum frequency, maximum frequency, duration and number of syllables) and four song-specific variables (minimum frequency, maximum frequency, duration and number of syllables). Acoustic variables were chosen by inspecting sonograms and examining the methods of similar studies (Mitani 1987, 1988; Cheyne et al. 2007a; Oyakawa et al. 2007) so as to ensure comparability and continuity with previous results. These variables were also chosen to capture the message being projected in the song rather than merely detecting the song. Three levels of acoustic variability were analysed: note, phrase and song, to capture as much variation as possible. Each acoustic variable was defined by a frequency (Hz) or time (s) component. Note-specific variables were measured for all notes in the song. The average frequency of a note was calculated by averaging the minimum and maximum frequencies. Phrase-specific variables were measured for both phrases 1 and 2 (Fig. 4).

Representative sonograms of a a single note from a male duet song and the six note-specific acoustic variables measured, b a single phrase from a male duet song and the four phrase-specific acoustic variables measured and c a single male duet song and the four song-specific acoustic variables measured
Statistical analysis
Principal component analysis (PCA) was performed to divide the acoustic variables into distinct groups, or so-called principal components (PCs). By definition, each PC is independent and contributes a certain proportion of the total explained variation found in the data set. PCs with significant contributions to variation were deduced from the Scree plot. The relative effect of each variable on the associated PC was deduced from the loading factor. Only variables with loading factor >0.5 were considered significant.
A one-way, random-effects model analysis of variance (ANOVA) was then performed on each set of PC scores. Between-individual variation was inputted as the main effect and within-individual variation as the error to deduce which PCs contributed most to individuality.
A variance component analysis (VARCOMP) was also performed to generate variance components for both within-individual and between-individual variation. Percentages were calculated using the sum of both variance components.
These statistical analyses were performed on note-specific, phrase-specific and song-specific variables separately. All statistical analyses were carried out using Minitab 16. All acoustic variables were checked for a roughly normal distribution, and outliers were removed from the data set before statistical analyses were performed.
Results
PCA and ANOVA: note-specific, phrase-specific and song-specific variables
Note-specific variables
A total of 1,562 notes, from 174 sonograms, from the nine males studied were measured and analysed to deduce whether the songs of H. albibarbis males show individuality at the level of the note.
Firstly, a PCA was performed to summarise the six acoustic variables at the level of the note into two PCs (Fig. 5) that explained 77.9 % of the variance in the data (Table 2). PC 1 was dominated by the variable of average frequency and PC 2 by the variables of start frequency, end frequency and minimum frequency (Table 2). The ANOVA showed significant differences in the means of both PCs across all nine individuals (Table 3). This suggests that the song of H. albibarbis males does indeed show individuality at the level of the note and particularly in the acoustic variables of average frequency, start frequency, end frequency and minimum frequency. However, due to genetic constraints (Geissmann 2000), individuality at the level of the note is visually less clear in the bivariate scatterplot (Fig. 5). Variance component analysis revealed that PC 1 and PC 2 represent a lower percentage of between-individual variance and a higher percentage of within-individual variance (Table 3).

A bivariate scatterplot of the two note-specific principal component (PC) scores for each individual
Phrase-specific variables
A total of 348 phrases (phrase 1 and phrase 2), from 174 sonograms, from the nine individuals studied were measured and analysed to find out whether the songs of H. albibarbis males show individuality at the level of the phrase. The PCA summarised the four acoustic variables at the level of the phrase into two PCs that explained 88.7 % of the variance in the data (Fig. 6; Table 2). PC 1 was dominated by the variables of duration, number of syllables and maximum frequency, and PC 2 was dominated by the variable of minimum frequency (Table 2). The ANOVA showed significant differences in the means of both PCs across all nine individuals (Table 3). This suggests that the song of H. albibarbis males does indeed show individuality at the level of the phrase, and in all phrase-specific acoustic variables measured. Some separation between groups in the bivariate scatterplot is evident (Fig. 6). Variance component analysis revealed that PC 1 and PC 2 represent a lower percentage of between-individual variance and a higher percentage of within-individual variance (Table 3).

A bivariate scatterplot of the two phrase-specific principal component (PC) scores for each individual
Song-specific variables
A total of 174 songs, from 174 sonograms, from the nine individuals studied were measured and analysed. The PCA summarised the four acoustic variables at the level of the song into two PCs that explained 81.1 % of the variance (Fig. 7; Table 2). PC 1 was dominated by the variables of duration and number of syllables, and PC 2 was dominated by the variable of minimum frequency (Table 2). The ANOVA also showed significant differences in the means of both PCs across all nine individuals (Table 3). This suggests that the song of H. albibarbis males does indeed show individuality at the level of the song and particularly in the variables of duration, number of syllables and minimum frequency. The separation between groups in the bivariate scatterplot is evident (Fig. 7). Variance component analysis revealed that PC 2 represents a lower percentage of between-individual variance and a higher percentage of within-individual variance whereas PC 1 represents a higher percentage of between-individual variance and a lower percentage of within-individual variance (Table 3). PC 1 at the song-specific level is the only PC which shows a higher proportion of between-individual variance than within-individual variance in the variance component analysis, suggesting a particularly high magnitude of acoustic individuality in this PC.

A bivariate scatterplot of the two song-specific principal component (PC) scores for each individual
Discussion
The aim of this study was to quantify the level of individuality found in the male duet song of the Bornean southern gibbon. This study has summarised 14 acoustic variables, at three levels of variation (note, phrase and song) and in three different analyses into a total of six PCs. All six of these PCs show significant differences between individuals, suggesting that the male duet song of H. albibarbis shows a significant level of acoustic individuality at all three levels (all p < 0.001). The note-specific acoustic variables which show significant individuality are start frequency, end frequency, minimum frequency and average frequency. All four phrase-specific acoustic variables show significant individuality, and the variables of duration, minimum frequency and number of syllables show individuality at the level of the song. The acoustic variables which were not included in the PCs and therefore did not show a significant level of individuality were note-specific and song-specific maximum frequency and note-specific duration. This study also found different magnitudes of acoustic individuality among the six PCs. PC 1 of the song-specific analysis (defined by the acoustic variables of duration and number of syllables) showed lower within-individual variance and higher between-individual variance, suggesting a particularly high magnitude of acoustic individuality. This may be because the level of the song is the highest and most interpretable level of variation.
The dense forest habitat of the gibbon is a challenging environment for sound transmission. Whitten (1982) points out that a signal can be weakened by factors such as foliage, air turbulence, temperature gradients, ground effects and other animal vocalisations which absorb, reflect and refract sound waves. Evidence for a so-called sound window in forest habitats can be found in several previous studies and is summarised by Waser and Brown (1984). In relation to gibbon song, Whitten (1982) suggests that sound transmission is maximised when a gibbon produces relatively low-frequency notes, from sites high above the ground and at a time just before dawn. One hypothesis, proposed by Haimoff and Tilson (1985), is that selection for individuality may therefore be limited when acting on frequency, to maintain the communicative function of the song. Furthermore, there may be greater selection for individuality on the durational variables and number of syllables in the song, features which are more easily manipulated (Haimoff and Tilson 1985). The results of this study therefore support this hypothesis. This hypothesis may also explain why maximum frequency did not show a significant level of individuality at the level of the note and the song. If it is important for gibbons to sing within a particular ‘sound window’ and lower frequencies optimise sound transmission, then high frequencies such as those sampled in the maximum frequency variable are likely to be poorly transmitted through the forest. If so, this may have an effect on recordings, making the maximum frequency variable less reliable than the other variables measured. It is possible that significant individuality would be found in this variable if this error was removed, for example by setting a lower limit on the distance of the group relative to the listening point when recording. The lack of significant individuality in note-specific duration may be due to limitations imposed on note length by vocal tract functioning and breathing of gibbons.
The findings of this study are consistent with those of Mitani (1987), who found significant inter-individual variation in songs of male H. agilis (since renamed H. albibarbis; Groves 2001) and H. muelleri in Indonesia. Similarly, Geissmann et al. (2005) found a high degree of inter-individuality in the note repertoire of H. moloch males. The findings of this study are also consistent with other studies of individuality in songs of female gibbons, including H. syndactylus, H. agilis (Haimoff and Gittins 1985; Oyakawa et al. 2007), H. lar (Raemaekers and Raemaekers 1985), H. moloch (Dallmann and Geissmann 2001a, b), H. klossii (Haimoff and Tilson 1985), H. muelleri (Mitani 1987) and H. albibarbis (Cheyne et al. 2007a). Cheyne et al. (2007a) demonstrated high individuality in songs of female H. albibarbis in all variables of the great call except duration. Similar studies have also found individuality in the vocalisations of different species such as the chuck vocalisations of female squirrel monkeys (Boinski and Mitchell 1997), marmoset long calls (Jorgensen and French 1998) and the vocal repertoires of wild chimpanzees (Mitani et al. 1996). The phenomenon of vocal individuality is therefore not exclusive to gibbons.
The function of gibbon song has long been debated. Gibbons are extremely territorial primates with a social structure based around the family unit. This is a very complex and demanding social and spatial structure requiring constant maintenance. One hypothesis is that individual gibbon song (both male and female) may help maintain this structure by advertising the individual’s presence in space and other important information regarding that individual (Haimoff and Tilson 1985). It has also been suggested that gibbon song strengthens the bond between a pair, with those pairs performing for longer having a stronger bond (Chivers 1974).
Furthermore, despite the lack of sexual dimorphism in size and the presence of an approximately equal sex ratio, mated male and mated female gibbons still experience very different selection pressures. Differences in the roles of the sexes in the duet suggest that selection for song individuality also differs between the sexes. However, this is likely to be a difference only in degree and not in any large qualitative sense, because of the approximately equal sex ratio. It is also likely that both males and females compete for mates but that different traits are valued and selected for in each sex. However, there is currently no evidence for non-monogamy in the species H. albibarbis. The long-term data set for Sabangau involves 6 habituated groups in adjacent territories which have been followed consistently since 2005. At no time have extra-pair copulations (EPCs) been observed. While we recognize that this is not conclusive proof, genetic samples are being collected to test for reproductive monogamy. At this time we have no reason to believe that the level of EPCs in Sabangau (and in the rest of the species) is as high as has been reported in other study sites for other gibbon species (Cheyne, personal observation). Nevertheless, we suspect that male song has both intra-group and inter-group functions. The male song carries different information at different distances, leading to specific information for within- and between-group communication (Cheyne, personal observation).
Whether H. albibarbis males can distinguish neighbouring males by their song patterns could be tested using playback experiments in future studies. Other playback experiments with gibbon song have found evidence for recognition among individuals (Mitani 1984; Raemaekers and Raemaekers 1985). However, one must always consider the possible long-term negative effects of playback experiments on their subjects before performing them. These may include confusion of the primates as well as other unclear impacts on their social structure and/or behaviour. We think this is particularly important to consider because the function of gibbon song is still not clearly understood.
The significant level of individuality found in the male songs of H. albibarbis also has important implications for conservation. One of the most important and challenging tasks is that of population surveying. Population surveys are a crucial method of monitoring the status of a species and informing conservation action. Censuses of primates, including density data and derived parameters, are badly needed (Nijman and Menken 2005). However, forest primates such as gibbons are particularly difficult to survey accurately (Brockelman and Ali 1987). Methods of estimating population numbers using the vocalisations made by gibbons are already widely used, but these rely on simple auditory methods such as the listening point sampling method. This method requires an estimation of the total listening area, and an estimation of the number of different groups within it, to calculate density. The method also requires triangulation for mapping of groups to estimate their locations. Listeners must stand at particular listening points and collect the following information: start time of each vocalisation, compass direction and estimated distance of each vocalising group or individual (Brockelman and Srikosamatara 1993). In the triangulation method as adapted to flat, peat-swamp forest, these listening points are arranged in a triangular layout. One limitation of this method is the difficulty of distinguishing gibbon groups from each other because a group may often sing more than once and from different places. Most groups can only be distinguished if they sing bouts that overlap in time, or if the calls map too far apart to be of the same group. Therefore, this method may underestimate the number of groups in the area by lumping bouts (Brockelman et al. 2009). Our technique may provide a way of more accurately determining the number of groups in the “listening area”.
We have shown the potential for identifying H. albibarbis males by recording and analysing their songs for the following acoustic features: note-specific start frequency, end frequency, minimum frequency and average frequency; phrase-specific duration, minimum frequency, maximum frequency and number of syllables; song-specific duration, minimum frequency and number of syllables and in particular, song-specific duration and number of syllables. Therefore, this method may make it possible to distinguish groups more easily from their individual song patterns (Brockelman and Srikosamatara 1993); For example, if each individual male (mated and lone) could be identified acoustically, then the number of groups could be deduced from the number of mated males. Furthermore, the number of individuals could be estimated by using a simple equation, such as
where N I is the no. of individuals, N MM is the no. of mated males, N LM is the no. of lone males and \( \overline{\text{gs}} \) is the average group size.
Average group size could be estimated or observationally deduced if sufficient time and resources were available. Another benefit of this method is that lone males could be sampled as well as mated males. This is particularly important when thinking about the dispersal of a population. From simple listening data, only mated males (groups) are sampled and lone males are ignored or are estimated from the number of groups heard (Cowlishaw 1992, 1996).
A method such as this would greatly facilitate accurate population surveys of H. albibarbis and would have considerable implications for the conservation of this species. It could also be extended by using a software package called Sonic Visualiser 1.9, which can automatically statistically analyse sound recordings for differences, making this acoustic method even more applicable. Of course, this method does not eliminate the desirability of mapping groups and solo males by triangulation; it is simply an additional tool which may facilitate the identification of groups.
One limitation of this method is the need for the listening point to be relatively close to the group for a clear enough recording to be made for sound analysis. In this study, all groups recorded were within 500 m. However, the method has not yet been tested explicitly against distance from the listening point. Another limitation is the need to process the data in the field and to map each individual bout recorded. This still needs to be resolved.
Furthermore, this study has also revealed the huge intra-individual variation, particularly in the notes and phrases that make up the male song of H. albibarbis. This has wider implications for the communicative functions of these songs and certainly warrants further investigation.
References
Boinski S, Mitchell CL (1997) Chuck vocalizations of wild female squirrel monkeys (Saimiri sciureus) contain information on caller identity and foraging activity. Intl J Primatol 18:975–993. doi:10.1023/a:1026300314739
Brockelman WY, Ali R (1987) Methods of surveying and sampling forest primate populations. In: Mittermeier RA, Marsh RW (eds) Primate conservation in the tropical rainforest. Alan Liss, New York, pp 23–62
Brockelman WY, Srikosamatara S (1993) Estimation of density of gibbon groups by use of the loud songs. Am J Primatol 29:93–108
Brockelman WY, Naing H, Saw C, Moe A, Linn Z, Moe TK, Win Z (2009) Census of eastern hoolock gibbons (Hoolock leuconedys) in Mahamyaing Wildlife Sanctuary, Sagaing Division, Myanmar. In: Lappan S, Whittaker DJ (eds) The gibbons: new perspectives on small ape socioecology and population biology. Springer, New York, pp 91–110
Cheyne SM, Chivers DJ, Sugardjito J (2007a) Covariation in the great calls of rehabilitant and wild gibbons Hylobates albibarbis. Raff Bull Zool 55:201–207
Cheyne SM, Thompson CJH, Phillips AC, Hill RMC, Limin SH (2007b) Density and population estimate of gibbons (Hylobates albibarbis) in the Sabangau Catchment, Central Kalimantan, Indonesia. Primates 49:50–56
Chivers DJ (1974) The Siamang in Malaya: a field study of a primate in tropical rainforest. In: Contributions of primatology, vol 4. Karger, Basel, pp 1–335
Cowlishaw G (1992) Song function in gibbons. Behaviour 121:131–153
Cowlishaw G (1996) Sexual Selection and Information Content in Gibbon Song Bouts. Ethology 102:272–284
Dallmann R, Geissmann T (2001a) Different levels of variability in the female song of wild silvery gibbons (Hylobates moloch). Behaviour 138:629–648
Dallmann R, Geissmann T (2001b) Individuality in the female songs of wild Silvery Gibbons (Hylobates moloch) on Java, Indonesia. Contrib Zool 70:41–50
Dallmann R, Geissmann T (2009) Individual and Geographical Variability in the Songs of Wild Silvery Gibbons (Hylobates moloch) on Java, Indonesia. In: Lappan S, Whittaker DJ (eds) The gibbons: new perspectives on small ape socioecology and population biology. Springer, New York, pp 91–110
Geissmann T (1995) Gibbon systematics and species identification. Int Zoo News 42:467–501
Geissmann T (2000) Gibbon songs and human music from an evolutionary perspective. In: Wallen NL, Merker B, Brown S (eds) The origins of music. MIT Press, London, pp 103–124
Geissmann T, Orgeldinger M (2000) The relationship between duet songs and pair bonds in siamangs, Hylobates syndactylus. Anim Behav 60:805–809
Geissmann T, Bohlen-Eyring S, Heuck A (2005) The male solo song of the Javan silvery gibbon (Hylobates moloch). Contrib Zool 74:1–25
Gittins SP (1978) Hark! The beautiful song of the gibbon. New Sci 80:832–834
Gittins SP (1980) Territorial behaviour in the agile gibbon. Int J Primatol 1:381–399
Gittins SP (1982) Feeding and ranging in the agile gibbon. Folia Primatol 38:39–71
Groves CP (2001) Primate taxonomy. Smithsonian Institute Press, Washington
Haimoff EH (1984a) The organization of song in the agile gibbon (Hylobates agilis). Folia Primatol 42:42–61
Haimoff EH (1984b) Acoustic and organisational features of gibbon song. In: Preuschoft H, Chivers DJ, Brockelman WY, Creel N (eds) The lesser apes: evolutionary and behavioural biology. University of Edinburgh Press, Edinburgh, pp 333–353
Haimoff EH (1985) The organisation of song in Müller’s gibbon (Hylobates muelleri). Int J Primatol 6:173–192
Haimoff EH (1986) Convergence of duetting of monogamous old world primates. J Hum Evol 15:51–59
Haimoff EH, Gittins SP (1985) Individuality in the song of the agile gibbon (Hylobates agilis) of Peninsular Malaysia. Int J Primatol 8:239–247
Haimoff EH, Tilson RL (1985) Individuality in the female song of wild Kloss’ gibbons (Hylobates klossii) on Siberut Island, Indonesia. Folia Primatol 44:129–137
Hamard MCL, Cheyne SM, Nijman V (2010) Vegetation correlates of gibbon density in the peat-swamp forest of the Sabangau catchment, Central Kalimantan, Indonesia. Am J Primatol 72:607–616
Hirai H, Hayano A, Tanaka H, Mootnick AR, Wijayanto H, Perwitasari-Farajallah D (2009) Genetic differentiation of agile gibbons between Sumatra and Kalimantan in Indonesia. In: Lappan S, Whittaker DJ (eds) The gibbons: new perspectives on small ape socioecology and population biology. Springer, New York, pp 37–49
IUCN (2012) IUCN Red List of threatened species, version 2012.2. http://www.iucnredlist.org
Jorgensen DD, French JA (1998) Individuality but not Stability in Marmoset Long Calls. Ethology 104:729–742
Lameira A, Wich S (2008) Orangutan long call degradation and individuality over distance: a playback approach. Int J Primatol 29:615–625
Marshall JT, Marshall ER (1976) Gibbons and their territorial songs. Science 193:235–237
Mitani JC (1984) The behavioural regulation of monogamy in gibbons (Hylobates muelleri). Behav Ecol Sociobiol 15:225–229
Mitani JC (1987) Species discrimination of male song in gibbons. Am J Primatol 13:413–423
Mitani JC (1988) Male gibbon (Hylobates agilis) singing behaviour: natural history, song variations and function. Ethology 79:177–194
Mitani JC, Marler P (1989) A phonological analysis of male gibbon singing behavior. Behaviour 109:20–45
Mitani JC, Gros-Louis J, Macedonia J (1996) Selection for acoustic individuality within the vocal repertoire of wild chimpanzees. Int J Primatol 17:569–583
Nijman V, Menken SBJ (2005) Assessment of census techniques for estimating density and biomass of gibbons (Primates: Hylobatidae). Raff Bull Zool 53:169–179
Oyakawa C, Koda H, Sugiura H (2007) Acoustic features contributing to the individuality of wild agile gibbon (Hylobates agilis agilis) songs. Am J Primatol 69:777–790
Raemaekers JJ, Raemaekers PM (1985) Field playback of loud calls to gibbons (Hylobates lar): territorial, sex-specific and species-specific responses. Anim Behav 33:481–493
Raemaekers JJ, Raemaekers PM, Haimoff E (1984) Loud calls of the gibbon (Hylobates lar): repertoire, organisation and context. Behaviour 91:146–189
Waser PM, Brown CH (1984) Is there a “sound window” for primate communication? Behav Ecol Sociobiol 15:73–76
Whitten AJ (1982) The ecology of singing in Kloss gibbons (Hylobates klossii) on Siberut Island, Indonesia. Int J Primatol 3:33–51
Acknowledgments
This work was carried out with the OuTrop-CIMTROP multi-disciplinary research project in the Bawan Forest, Central Kalimantan, Indonesia. Special thanks go to all OuTrop volunteers and organisers for their help in this project. Thanks to Science Oxford for kindly hosting our fundraising event and to BBC Radio Oxford for their help publicising it. Funding was provided by a St. John’s College Special Grant, the Mike Soper Bursary Fund and through private donations. We would especially like to thank A & B Ekert for their considerable private donation. Finally, we thank Professor Simon Bearder and others who helped to review this work.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Wanelik, K.M., Azis, A. & Cheyne, S.M. Note-, phrase- and song-specific acoustic variables contributing to the individuality of male duet song in the Bornean southern gibbon (Hylobates albibarbis). Primates 54, 159–170 (2013). https://doi.org/10.1007/s10329-012-0338-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-012-0338-y