
Abstract
Three levels of hamadryas social structure—the one male unit (OMU), the band, and the troop—have been observed at all sites studied, but a fourth—the clan—has been observed at only one site, Erer-Gota, Ethiopia, during a longitudinal check of the dispersion of identified individuals. The clan is important since it appears to provide the basis for male philopatry, although comparative data is needed from other sites to confirm this. We studied a huge commensal group of hamadryas baboons (over 600 animals) in Saudi Arabia. We put ear tags on baboons between 1998 and 2004 and analyzed social structure, relying on the interactions of these tagged animals by focusing especially on their dispersal patterns from OMUs. OMU membership tended to be looser than that of the Ethiopian hamadryas. Females tended to shift between OMUs on an individual basis in our study group, whereas the collapse of an OMU was a major occasion of adult female transfer in Ethiopia. We found neither stable bands (a “band” in our study group was defined as a regional assemblage of OMUs) nor clans that lasted for several years. Some OMUs moved and transferred into neighboring areas over both the short and long term. Further, some post-adolescent males appeared to move out of the study area. The ratio of adult females in an OMU in our study group was larger than for any other documented study site, and this may be the reason for enhanced female transfer between OMUs. A large proportion of the adolescent females showed no clear membership to OMUs, and no “initial units” (commonly observed in Ethiopia) were discernible. The ease with which young males acquired adult females at the study site must have disrupted the formation of a clan, a “male-bonded society.”
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Among primates, hamadryas baboons (Papio hamadryas) have a unique multilevel social structure where one male units (OMU), bands, and troops have been found at all sites studied (Kummer 1968; Swedell 2002a, 2002b; Zinner et al. 2001a, 2001b). An OMU is a clear social unit where members always move together. The band can be defined as a social unit, as its members (OMUs) move in the same direction and meet together at midday at a water site, though they sometimes separate. Although bands move independently, they tolerate each other at the same sleeping site, forming a troop. The Swiss team observed the presence of a clan level between the OMU and band levels at a site in Erer-Gota, Ethiopia (Sigg et al. 1982; Abegglen 1984). Males are philopatric in their native clan, whereas females transfer between units, clans and bands. A clan is formed from 3–4 OMUs, whose leader males are proposed as being bonded in a kin group (Abegglen 1984). The clan is somewhat difficult to understand, as it cannot be readily identified in the short term; long-term observations of the dispersion of identified individuals are required to identify clans. Abegglen (1984) stated that, “these groupings, which we named “clans,” rested and traveled together within the band.” This is not a clear-cut definition, so we can only suppose that a compact group inside a band is a clan.
Male philopatry is a rare social characteristic among the Cercopithecoidea (Wrangham 1980; Pusey and Packer 1987). The closely related species Papio anubis exhibits a mono-layer, multi-male, multi-female society with female philopatry. The two Papio species are estimated to have separated 330,000 years ago based on blood protein analysis (Shotake 1981). At their species border, however, they form a hybrid zone, giving rise to some very complicated hybrid social structures (Nagel 1973; Sugawara 1979, 1982, 1988; Bergman and Beehner 2004). As these species were separated only rather recently, yet they display very different social structures, we can expect some variations in social structure among hamadryas populations. Kummer et al. (1985), for example, found some different social behavior patterns in Saudi Arabian populations. Our study focused on determining whether or not the social structure elucidated at Erer-Gota is also found in Saudi populations. Groups favorable for observation in Saudi Arabia, however, are commensal groups that depend on human food waste (Kummer et al. 1985; Biquand et al. 1992a; Boug et al. 1994). These groups have huge population sizes and it is difficult to identify individual baboons. To enable us to analyze social structures and dispersal patterns of individuals more easily, we attached ear tags (Yamane et al. 2003).
Methods
Taif City is situated on a plateau at an altitude of 1,700 m and is 16 km east of the escarpment that rises from the Red Sea. Annual rainfall is around 233 mm (Biquand et al. 1992a). The altitude just above the escarpment is much higher, ex. 3,000 m at Al Shafa, and much heavier rain is experienced. A dike for the dam was constructed in the dry river to the south-east end of the city (Fig. 1). This river flows only when it rains up-river. The dam lake was present until 2001, and the water spread 1.5 km up the dike. The lake was drained in 2001 and has never recovered. After the dam was drained, the baboons drank at one part of the mostly dry riverbed, where a small amount of water emerged from below the ground, flowed for 50 m, and then disappeared into the sand again. This was the only water site for baboons in the area, except for the dirty water at the dike, which they did not drink. The study group, the dam site group, slept on the rock face at the dry river (the water site), and fed on human food waste at Al Ruddaf Park, 3 km northwest of the sleeping site, and also on natural food from an Acacia woodland surrounding these two sites (Fig. 1). The population size was a maximum of 475 and then 610 in 2000 and 2004, respectively. Adult sex ratio was heavily biased towards females: 4.79 females/male on average over three periods, August 2000, January 2001, November 2002 to January 2003.

Map of the study site
Tagged baboons were observed for about one month each summer during 2000–2004 (August 16–September 12, 2000; August 9–22, 2001; July 29–August 26, 2002; August 5–September 17, 2003; July 22–August 26, 2004). Many tourists visited the park and dam in summer and people spending the evening or sleeping in the park produced large quantities of food waste. The baboons visited the park very early in the morning and fed on this waste before service workers could remove it.
Ear tags were attached in 1998 and 1999, before the present observations, and also between November 6–14, 2000, January 20–27, 2003, and on December 3, 2003, thereby avoiding the summer observation periods.
Capture
We used a steel mesh cage (3 × 2 × 1.5 m) to enable us to capture a whole unit at once and thereby minimize the effects of capture. We lured them into the cage using peanuts as bait and shut the door after the leader male entered; the leader male tended to be more cautious than the others and entered last (see details in Yamane et al. 2003). Recapture of tagged animals occurred in only three cases over the whole study. We did not use data on the composition of OMUs at the time of capture, and observation of tagged animals did not begin until the next season, so the effects of capture on tagged animals were presumably small, except for the three cases of recapture.
Ear tags
Each tagged animal was identified according to five characteristics: sex, age, the type and color of the right and left ear tags, and any other distinguishing physical characteristics. An ear-tagged animal was identified by the following formula: right tag : left tag : sex : age : other characteristics
Letters indicated the colors of the tags: R: red, O: orange, G: green, Y: yellow, B: blue, W: white, NO: no ear tags. Tags were from Allflex (Dallas, TX, USA), and were usually round and 30 mm in diameter (small tag). This type of tag was expressed only by a color. The L-Color, a narrower type of tag where a round tag was cut down at two ends to form a narrow tag (about 1.7 cm wide) was introduced in January 2003. The SQ-Color, a rectangular (or pentagonal) (Hog Tag 3.5 × 4.5 cm) tag, was introduced in 2000. Color#: this category was introduced in December 2003, because the manufacturer changed the tags to paler colors. This type appears only in the data from 2004. Thus, one ear with a tag was expressed as “(type)color(#)” or “NO,” though type or # in parentheses are absent in particular cases. Sex: M: male, F: female. Age: A: adult males have the fully developed silvery gray mantle. S: subadult males can be categorized from the first sign of a mantle on the shoulder to the well-developed brown mantle of the adult. Adolescent males: we defined adolescent males as males that have not yet reached the subadult state and are the same size or larger than adult females. They are categorized as being 5–6 years old. Juveniles are 1–4 and do not include black and brown infants. Females were categorized as being 1–4 years old or as A (adult). Although Y was used to depict young adult males and females, they were included in the adult (A) category in the calculation.
The problem with using ear tags is that they sometimes fall off and we cannot necessarily discriminate between the loss of ear tags and the disappearance of a baboon. Further, when a tag falls off one ear but not the other, the identification of the animal changes completely, and, as a result, a few individuals sometimes displayed the same ear tag pattern. In order to understand the complexities relating to tagged animals we must be able to follow all existing tagged animals in a study period. We could sometimes identify a newly emerged tagged baboon as a previously tagged animal of corresponding age and sex that had disappeared. When an OMU continued to exist over two consecutive years, an animal that had lost one of its ear tags could be identified with certainty by identifying other marked individuals within this group.
A new identification characteristic was introduced in January 2003; large numbers were hand-written on the tags. These numbers could be read over a short distance, and some adult males were identified in this way during the observations of 2004.
Although ear tags were usually attached in November and December, the capture was late in 2002 and could only be conducted in January 2003, but we nevertheless recorded this tagging event as 2002 in Table 1. We did not observe all previously tagged baboons (Table 1) in each study period, as some ear tags had fallen of and some baboons really had disappeared. The number of baboons identified in each study period was used as the basis for sociological analysis (Table 2).
Observation of ear tags
Ear tags were identified from a distance of 30–50 m using a Nikon Field Scope ED (×20) while sitting on rocks facing the water sites in the park or in the dam.
Observations in 2000 and 2001
All baboons moved swiftly at dawn from the sleeping/water site in the dam in the direction of the park (Fig. 1). Upon arriving at the park, they searched for the huge amount of garbage left there overnight. After feeding, they visited the natural spring (pool) in the park to drink, especially after feeding on dried bread discarded in the park. They took a long rest near the spring until noon. Observations were conducted from a rock overlooking the spring. The major objective of the observations made in the summer of 2000 was to identify the OMUs of the baboons that moved in file together, and to count the number of adult females in the OMUs. The ear tags were identified in preliminary work in 2000, and we recorded tagged baboons only when we noticed them in an OMU, so not all tagged baboons were recorded.
Observations in 2002
In the summer of 2002 baboons stayed in the park for shorter durations than before, arriving at around 6:00 a.m. and departing at around 7:30 a.m. We checked the tagged animals at two sites, both the park and the dam, and these two sets of data were analyzed separately. We waited at the park’s spring during the early morning and checked all tagged baboons. All baboons observed in the park on the same day were regarded as one group in the analysis. When the first baboon group started to move out of the park, we drove the 3 km to the dam water site and waited for the returning groups. In many cases, however, we found that a group of baboons was already at the dam, and presumed that these baboons had not visited the park that morning.
Observations in 2003 and 2004
The behavior of the group changed greatly in 2003/2004. Baboons rarely visited the park during these study periods, probably because tourists could drive cars to the water site of the dam. We conducted observations only at the water site of the dam. Baboons departed from the sleeping/water site before 6:30 a.m. to feed a little farther away. They then completely vacated the surrounding area; the earliest returning group sometimes came back around at 10:00 a.m.; at other times, around noon; and, after that, small groups came back one by one. Almost all known marked individuals had come back by 2:00 p.m., when we stopped observations. After drinking water at the riverbed, baboons climbed the hills and stayed on the hills surrounding the water site. When a new group came to the water site to drink, or when people brought food, many OMUs that had taken rest in the surrounding hills also came down. Thus, the definitions of the groupings changed in 2003 and 2004. As we wanted to know which groupings moved in the home range together, the newly arrived groupings were important, so those baboons which joined the arriving group after resting in the surrounding hills were not considered to be a part of this. Every time baboons came down to the water site, tagged baboons were checked, but individuals that had already come down that day were excluded from the newly arriving group in the data analysis.
Identification of units
When a group of baboons came down to the water site, they came down separately in several smaller compact groups, which included one adult male and several adult females, often descending in file. Animals filing down together were regarded as members of the same OMU. However, upon close observation, there were still some females left behind or that moved apart from other members, and these were not regarded as being part of the same OMU. When artificial food was distributed over a small area where only one adult male and several adult females could feed together, we regarded these co-feeding individuals as being members of the same OMU. After pooling these data, we extracted OMUs. Juveniles were sometimes observed to sit close to or groom adult females, whereas they were observed only rarely in the moving file of an OMU. Though many of the OMUs we identified did not include juveniles as members, this should be regarded as an observational artifact because mother/juvenile relationships did, of course, exist.
Names of an OMU are given as OMU (year number). The years 2K, 2K1, and 2K2 stand for 2000, 2001, and 2002, respectively.
Results
Persistence of ear tags and tagged animals
By summing the number of ear tags observed each year and the number of newly attached ear tags for that particular year, we can determine the maximum number of ear tags which could be observed in the following year. By comparing the expected number with the number of observed tags in the next observation period, we can calculate the persistence ratio of ear tags (Table 3). The persistence ratio of square tags was low compared to other forms of tags, as their corners are easily caught. Most of the square tags were put on adult individuals, and the persistence ratio among females (14%) was far lower than that among males (56%). As the difference in the tendencies of males and females to lose ear tags was expected to be similar for both square and round ear tags, we expected a high tendency for females to lose even round tags. In contrast, the persistence ratio of round tags among females, 75%, was far higher than that among males, 43%. The persistence ratios of round ear tags among adolescent and subadult males were 58 and 44% respectively, which is lower than that found among adult females, 75%. The low persistence ratio of round ear tags among males is most likely due to the disappearance of male individuals. Narrow tags were not put on adult males. The persistence ratios were lower for juvenile males, 51%, than for juvenile females, 81%.
The number of OMUs that could be identified with marked individuals was 8, 19, 14, 15, and 24 in 2000, 2001, 2002, 2003, and 2004, respectively. The change in the number of identified OMUs is partly due to an increase in identified individuals and partly due to better efforts made to identify the individuals. Members of an OMU tended to be captured at the same time, and these marked individuals tended to disperse among different OMUs as the years passed, as indicated by the number of OMUs with only one marked individual, which increased over the course of the study: five in 2002, two in 2003 and ten in 2004.
Characteristics of OMUs
The frequency of female transfer between OMUs
We compared the membership of an OMU over two consecutive years in order to ascertain the stability of an OMU. This could be determined only in OMUs that had at least two tagged females. The criteria for transfer between OMUs were: (1) if two marked females were present in the same OMU but had separated into two OMUs by the following year, then we assumed that one of them had transferred and the other did not; (2) if one of the subsequent OMUs included the original leader male, then we assumed that some females had not changed their OMU, whereas the others had; (3) when a daughter OMU without the original leader male after division was overtaken by another adult male, the invading leader male and his OMU females were not regarded as cases of transfer; (4) however, when the number of invading females was much smaller than the number of females overtaken, then the invading group was regarded as a case of transfer. In such a case, the females overtaken were not regarded as a case of transfer, even if they were in a divided group without a leader male. To avoid overestimating unstableness in OMUs, we used the most conservative estimate of the rate of transfer in every case.
The numbers of OMUs with more than two marked individuals (+: including other than adult females) were 6 (+1), 8 (+2), 7, and 12 (+1) in 2000, 2001, 2002, and 2003, respectively. Compiling the data from 2000 to 2004, the proportion of adult males that stayed in the same OMU per year was 62% (Table 4). This proportion increases to 81% when we exclude disappearances, as this may have included cases of lost ear tags. The proportion for subadult males was 0%, and the proportion was 23% (28% excluding disappearances) for juvenile and adolescent males combined. When we calculated the rates of transfer for females, we excluded those cases where leader males were shot dead. The proportion that stayed in the same OMU was 63% (71% excluding disappearances), and 33% (36%) for juvenile females.
Five adult females who did not show clear membership to any particular OMU were either observed to belong to an OMU in the following year or to disappear (Table 4). Of seven juvenile females without clear membership to OMUs, three continued to have unclear membership in the following year. One of five adult males without clear membership joined an OMU in the following year. Of 29 juvenile males + adolescent males + subadult males who showed unclear membership, 10 joined OMUs, 11 continued to be unclear, and 8 had disappeared by the following year.
Types of OMU transfer
Two leader males were shot dead on September 7, 2000. One was the leader male of the OMU(2K-1). This OMU had five adult females who subsequently transferred into four other OMUs. These females did not show any tendency to gather in a single OMU in the following year. The second OMU whose leader was shot was OMU(2K-2), but it did not have any marked adult females. The leader male of OMU(2K3-9) was shot dead on August 31, 2003. This OMU had three marked adult females (O:Y:F:A:, Y:B:F:A:, NO:B:F:A:YOUNG), and one young adult male with a wire around his neck, NO:NO:M:A:PLASTIC. Within a few days, the female Y:B:F:A: joined an OMU with six adult females, two of which were marked, O:NO:F:A:, NO:G:F:A:. The other two marked females and the young adult male joined an OMU without marked members that included seven adult females. The four marked individuals of the OMU(2K3-9), were found in four different OMUs in the following year. The loss of the leader males resulted in complete dispersion of OMU members in subsequent years. Cases of dispersal of all OMU members were found in only two other instances, OMU(2K-3), OMU(2K1-15), though we do not know if this was precipitated by the deaths of leader males. To examine the dispersal patterns of OMU members, the collapse of an OMU or the partial desertion of OMU members, we need more than two marked members in an OMU. The numbers of OMUs which had more than four marked individuals were 3, 4, 6, and 7, in 2000, 2001, 2002, and 2003, respectively, which was 20 OMU years in total. We excluded the first transfer of OMUs directly after the killing of leader males, 2 OMU years. The relevant OMUs were then 18 OMU years. Two of the 18 OMU years broke down for unknown reasons. Sixty-four adult female years were calculated for the 18 OMUs. We excluded eight females that had disappeared, perhaps due to loss of ear tags, and did not count them as cases of transfer. So the females of concern numbered 56 (=64–8) female years. 35 female years stayed in the same OMUs, so the ratio that remained was 62% (=35/56). Transferred females numbered 7 female years, excluding OMU breakdown, so the transfer ratio was 13% (=7/56). One female was ambiguous in terms of which OMU she belonged to. Departure of females from an OMU without the collapse of OMUs are 14%[(7+1)/56]. In two cases where OMUs were divided, seven females moved to daughter OMUs other than that of the leader males, the ratio being 13% (=7/56). When we add 13% of transfer because of OMU division to the ratio of departure from OMU without collapse, the ratio of departure (=transfer) is 27%[(7+1+7)/56.
Extraordinarily large OMUs
We also found extraordinarily large OMUs. OMU(2K2-2) consisted of one adult male and seven adult females (four of them were marked) in 2002, and this OMU became a huge group, OMU(2K3-3), accepting many new members in 2003. The OMU(2K3-3) consisted of one adult male and 11 adult females of whom eight were tagged. It also included eight marked juveniles and one marked subadult male. Many of the new marked members were individuals that had been tagged in the preceding year. The peculiarity of this OMU is that it included many juvenile and adolescent individuals that don’t usually exhibit clear membership in OMUs. The huge OMU(2K3-3) had divided into three OMUs by 2004. The leader of OMU(2K3-3) formed an OMU kept one of his marked females, and added two marked females from other OMUs in 2004. Daughter OMU(2K4-3), with three marked females from OMU(2K3-3), was invaded by the leader male of OMU(2K3-8) that had kept one of his two original females but had lost the other. The other daughter, OMU(2K4-4), included two marked adult females from OMU(2K3-3) and the leader male that was the marked former subadult male of the same OMU, adding two more adult females from another OMU.
The large OMUs observed with ten or more adult females were OMU(2K1-1), OMU(2K1-13), OMU(K1-15), OMU(2K2-4), OMU(2K2-10), OMU(2K2-11), and the abovementioned OMU(2K3-3). Among these seven OMUs, OMU(2K1-1) included nine marked individuals, and a part of OMU(2K1-1) behaved like a subgroup in the following year, 2002 (see in details below). OMU(2K1-15) is the abovementioned case of OMU collapse in 2003. The leader male of OMU(2K2-10) was shot dead in 2004. OMU(2K3-12), which had nine adult females, divided into three OMUs in 2004.
Pair OMUs
We found OMUs that exhibited bonds among males. OMU(2K3-13) had one marked adult leader male and one marked subadult male in 2003. The subadult male acquired one adult female and formed an OMU in the next year, while the leader male kept his OMU. These two OMUs frequently moved together in 2004 (referred as “PAIR OMUs 1” hereafter). OMU(2K2-1) and SUB-OMU shared four marked adult females and one marked five-year-old female of OMU(2K1-1), respectively. They could have been two OMUs, but they moved together and the former leader male always behaved dominantly (“PAIR OMUs 2”). OMU(2K4-1) and OMU(2K4-16) also moved together and their boundaries were not apparent when the two OMUs were sitting adjacent (“PAIR OMUs 3”). More information on these two OMUs will be given in the following section.
Band
Although groupings which arrived at the water site at the same time could be defined as a band, we adopt a different definition where groupings of various sizes, temporary groupings, arrived at the water site of the dam, as the distance between the park (the major feeding site) and the dam was only 2.5–3 km (Fig. 1). We must, however, know the association ratio between individuals or between OMUs to be able to determine whether or not a grouping is temporary. Were we to obtain a clear-cut cluster in a cluster analysis of association rates between individuals, we could consider that cluster a band. If the membership of a cluster is clear, and if the cluster shows unity against other members, for example at times of conflict, we could assume that the cluster might be a clan. The membership of males in a clan should not change much over time, as males do not leave their natal clan (Sigg et al. 1982). The ratio of association between individuals was calculated as memberships in groups coming to the dam water site, and cluster analysis was conducted using the STATISTICA, WPGMA method (Fig. 2). As cluster analysis was conducted only among adult and subadult individuals in 2003, some of the individuals in 2002 (Fig. 2) had no corresponding data points in 2003 (Fig. 3). Similarly, because 86 individuals were newly marked in January 2003, some had no corresponding data point in the figure for 2002. Individuals that do not appear in figures for 2002 and 2003 are indicated in each figure.

Cluster analysis of the association of individuals in groups coming together at the dam water site in 2002. Individuals are identifiable by ear tags (see details in “Methods”). Crossed individuals were not found in 2003

Cluster analysis of individuals in 2003. Crossed individuals were not found in 2002. The clusters to which individuals had belonged to in 2002 are indicated by the letters A, B, C, and D at the bottom of each tree bar
We identified four clusters, A, B, C, and D, in 2002 (Fig. 2). The individuals highlighted by thick bars in cluster D, though not clearly separated in the analysis, were most distantly related to the others. Behaviorally, they were clearly distinct from the others in observations, since they were found in a group that did not visit the park with any other marked animals. They belonged to a band of about 70 individuals, and stayed around the dam. As they did not visit the park and behaved hesitantly around the other baboons, they were rarely captured, so the ratio of marked individuals was extremely low, with only two tagged individuals and two with other identifiable markers. The same kind of cluster analysis was conducted for associations in the park. Individuals found in the Park on the same day were regarded as belonging to the same grouping and association ratios between individuals were calculated. The clusters were similar for both the dam and park data, except for the D cluster.
We performed a cluster analysis of the association of individuals that came to the dam water site in 2003 (Fig. 3). The individuals common to the 2002 data (Fig. 2) are highlighted by the letter of the cluster in 2002 at the bottom of the tree bars. D band disappeared from the study area. Members of the A, B, and C bands in 2002 were mixed in the clusters of 2003. B band members were closely associated into something like a clan in 2002. However, two of its members, a young adult male, R:R:M:A, and another subadult male, W:G:M:S:, formed new OMUs upon gathering females from the A band in 2002. These two OMUs had a rather close relationship.
As the number of marked individuals increased, it became difficult to make the same kind of cluster analysis based on individual baboons in 2004. We conducted cluster analysis of associations between OMUs that came together in a group at the dam water site. Although the resulting figures are not shown, the clusters do not show any consistent correlation with the clusters of 2002 or 2003. We found strong associations for two pairs of OMUs (PAIR OMUs 1 and PAIR OMUs 3).
Relationships with neighboring populations
Case 1:
We found a female with a red square tag on her right ear (SQR:NO:F:A:) on September 16 and 17, 2003, though the female was never observed again during the 38 days of observation. The marked female belonged to one of the two OMUs, which visited from the west, beyond the home range of the study group. The female, SQR:NO:F:A:, must be one of the three females on which we placed red square ear tags on the right ears in 2000. They were not observed in the study period of 2001 and 2002. This case indicates that another neighboring population exists, and a part of this joins the study population on rare occasions.
Case 2:
The D band of 2002 disappeared in 2003 and was never observed again.
Case 3:
Two OMUs, “PAIR OMUs 3,” included four and five marked individuals that were captured in January 2003, respectively. These marked individuals did not come to the study site during the 44 days of observation in the summer of 2003. The two OMUs were present throughout the study period of 2004 and they moved together. They always sat adjacent among many other OMUs at the water site.
Discussion
OMU
According to a report from Ethiopia (Sigg et al. 1982), the transfer of females from an OMU occurred at the same time as OMU breakdown resulting from the leader male being ousted. Independent female transfer rarely occurs and more than 70% of adult females stayed in the same OMU for more than three years. We recalculated the data from Sigg et al. (1982) for 1971–1977 (Fig. 5 of that work). Changes in OMUs between two consecutive years during the seven years of their study can be calculated; changes from 1971 to 1972, from 1972 to 1973, ..., and from 1976 to 1977. The data totaled 40 OMU years for 113 adult female years. Female transfer was observed 18 times where OMU breakdown occurred in six OMUs with 15 cases of transfer, and independent adult female transfer occurred only three times. The other seven adult females disappeared and six of these disappearances occurred when the OMU broke down. These six females possibly transferred to less well-known bands. We can estimate that 24 (18 + 6) adult females transferred in total. The proportion that remained in an OMU as estimated by the ratio of adult females/year is 0.78 [(113 − 24 − 1)/(113 − 1)]. The ratio of independent transfer of adult females was 0.026[3/112].
In our study group, adult female members of an OMU were observed to transfer comparatively often. If we exclude the cases in which leader males were shot dead, the ratio of adult females remaining in the same OMU was 63% (Table 4). This figure goes up to 71% if we exclude females that disappeared, as some of these disappearances were undoubtedly caused by loss of ear tags. Sigg et al. (1982) stated that “70% of observed bonds lasted three years or more” (bonds between males and females). This figure, 70% over three years, is the equivalent of 88.7% per year. However, the present fractions remaining in an OMU/year, 63%, should be compared with the recalculated Ethiopian ratio of 78% given above. Saudi OMUs, therefore, are more unstable than Ethiopian ones. Furthermore, the type of transfer between OMUs was different between the two populations. The ratio of female transfer without collapse of OMUs was 13%, which is much higher than that of Ethiopia, at 3%. When OMUs collapsed in Ethiopia, the leader males lost all of their adult females as defeated leaders (Sigg et al. 1982). In our cases of division of OMUs, the leader males still kept some of his females, adding more adult females from other OMUs. Females in daughter OMUs other than those of the leader males are assumed to be partly responsible for their transfer. When we add 13% of transfer due to OMU division to the ratio of independent transfer, the ratio of transfer because of loosely organized OMUs is 25%[(7 + 7)/56]. Thus, the OMU in the Saudi Arabian commensal group is looser and exhibits less stability than the OMU in Ethiopia.
The paternity of infants in OMUs was checked using microsatellite DNA, and many infants were found to have been sired by males outside of their OMU (Yamane et al. 2003). This result is expected given the frequent transfer of females between OMUs.
The OMUs that became huge and then divided into daughter OMUs seem to share characteristics in common with an Arsi gelada OMU that had many adult males (Mori et al. 2003). The gelada OMU, C5, had many adult males, adolescents and juveniles and an adult female. The OMU was characteristic of an all-male group, yet it also seemed to function as a recipient group for individuals that overflowed from ordinary OMUs. Though the huge OMUs of our observations had only one adult male, they do have some common characteristics. When the movements of females increased and females moved out of OMUs, this huge OMU of loose integrity seemed to function as a repository for these females. In this loose OMU, even juveniles were accepted, as in the gelada case. We found many juveniles in the large OMU(2K3-3). This result supports the preliminary observation of Kummer et al. (1985), that Saudi females behave more independently than Ethiopian ones.
There is, however, no difference between Saudi Arabian and Ethiopian hamadryas societies in the sense that the OMU exists in both cases. Adult females of both populations transfer between OMUs. Juvenile and adolescent females of Saudi populations showed only loose membership with OMUs. These observations suggest that female philopatry based on an OMU does not exist in either the Saudi or the Ethiopian populations.
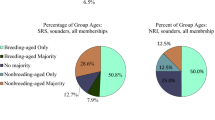
The number of adult females in an OMU in the study group was larger than that observed in natural populations. The peak distributions in the numbers of adult females in OMUs in wild groups from Ethiopia and Saudi Arabia are two to one (Biquand et al. 1992b; Kummer et al. 1981), while the peak in our study group is five (Fig. 4). This might have resulted in looser associations of females in an OMU. Certain members of OMUs were sometimes observed to move a little further away from others, but they moved in the same group to travel. The larger number of adult females in an OMU is probably influenced by the biased adult sex ratio (4.78) in this population. The ratio is much higher than any previously reported, 1.80 in Ethiopia, 2.87 in Eritrea, 2.18–2.51 in Saudi Arabia (Zinner et al. 2001b). A biased adult sex ratio can result in a species where a very different age of maturation occurs between the sexes (Ohsawa and Dunbar 1984). Another cause could be a tendency towards selective killing of adult and subadult males by local people. The conspicuously biased adult sex ratio cannot be explained solely by food conditions. It is worth noting that a fundamental social characteristic of this population, whereby young males can easily acquire females, is affected by human influences.

Frequency distributions of the number of adult females in a unit for each group. The data for Ethiopia and Saudi Arabia wild groups were taken from Table V of Biquand et al. (1992b). The data for Ethiopia were taken at Erer-Gota in 1961 and 1974 (Kummer et al. 1981). Following the method of Biquand et al., the frequency (%) of a female number of larger than four was summed for the Saudi data
Band
We obtained different results between 2002 and 2003. We could observe relatively clear groupings in 2002. The D band containing around 70 members came to the dam, but not to the park. It had disappeared from the study site by the next year. Furthermore, the B band sometimes fought with the A band at the water site of the dam and in the park. Young adult males and subadult males of B band actively and cooperatively participated in face-to-face confrontation between the two groups. These observations indicate the existence of some kind of group membership, especially among B band members. However, members of A and B together with C were mixed in the clusterings of 2003. This suggests that some kind of loose grouping can occur among OMUs but that it does not continue beyond a year.
We presumed that the B band might actually be a clan, as its males seemed so closely knit. The B band seemed to fit Abegglen’s statement that “these groupings, which we called “clans,” rested and traveled together within a band” (Abegglen 1984). Furthermore, some members of the B band continued to exhibit close relationships after two years. Two pairs, (R:R:M:A: and W:G:M:6:) and (SQR:NO:M:A: and NO:Y:M:Y:), still existed in 2003, though the pairings were separated into different clusters. A similarly close relationship was observed among young males in the case of “PAIR OMUs 1.” Abegglen (1984) considers that the kidnapping behavior in which young males took care of infants is the basis for the development of bonds between leader and follower males, and there should be a slight age difference between bonded males. This seemed to fit for R:R:M:A: and W:G:M:6: or LY:LY:M:S:. With these results, we can say that young males may maintain close ties for at least two years, but that such ties do not develop into a clan. Furthermore, two young males of the B band, W:Y:M:Y: and G:W:M:S:, disappeared in 2003. We could confirm some tendency towards male bonding among young males. We can say that behavior patterns that can sustain a clan were present but that the clan itself was not confirmed in the commensal group in Saudi Arabia.
The park was not only a huge feeding site with much discarded food (Fig. 1), but also a dangerous place where people chased them with stones, so the baboons collected into a large group to visit there. When a number of bands of different origins use a larger feeding site, and when they use the same sleeping/water site, it may be difficult to distinguish between bands and troops. A large group size (50–220) of hamadryas baboons has been observed in Ethiopia, and their main food was doum palm nuts (Swedell 2002a). Large groups of 25–800 at sleeping cliffs and 7–550 in feeding groups are known from Eritrea (Zinner et al. 2001a), though group size was not clearly correlated with a food resource. The band sizes of Erer-Gota ranged between 52 and 90 (Abegglen 1984), and the troop size at Cone rock was 236 (Sigg et al. 1982). There might be some difference in the social levels of clan and band in larger groups in other study sites.
Local population
The disappearance of the D band or some adolescent and subadult males suggests the dispersion of these individuals to neighboring populations. Furthermore, two OMUs from outside of the dam site population temporarily joined the dam site group. Many individuals marked in one year were lost in the next but then reappeared in two OMUs in the following year. These findings indicate that the dam site group is not a closed system and that it interacted with neighboring populations. Emigration of males and OMUs beyond the home range does not accord with expectations of male philopatry. These findings indicate that OMUs have some independence and they move out of and come back to an area, ranging over a much wider area. This kind of regional fluidity of OMUs has some similarity with the gelada multi-level social structure (Ohsawa 1979; Mori et al. 1999, 2003).
Development of the clan system
Though the movement of OMUs between areas does not conform to male philopatry and the clan structure, pairs of OMUs showed “brotherly bonding,” which is a characteristic of the clan. The question is: why don’t these “brotherly bondings” continue long enough to develop into a clan? When some of the female members of an OMU independently move out of an OMU, it is possible for young bachelor males to form new OMUs with these breakaway females. This seems to be a major reason for not forming the male alliance of a “clan” to defend females as a scarce resource. We did not observe any typical “initial unit” of young males and juvenile females in our study group, whereas it is an important process for young males in Ethiopia (Sigg et al. 1982). Male philopatry based on clans seems nonexistent in the study group, and the disappearance of young males also argues against it. As male philopatry is a means to acquire females, if conditions are such that females can easily be obtained, then male philopatry could disappear.
Our results, however, might not be common in wild groups in Saudi Arabia, as a change in population structure during the commensal situation might have affected the above social relationships. Changes in feeding conditions could alter the very basic social structure of hamadryas baboons, the clan, and male philopatry. Male social behavior in OMUs with the same level of hybrid index in different groups were much affected by differences in group social structure (Bergman and Beehner 2004). Thus, the consideration of behavioral change as a consequence of group structure is paramount when investigating the emergence of the hamadryas social system.
References
Abegglen JJ (1984) On socialization in hamadryas baboons. A field study. Associated University Press, Cranbury, NJ
Bergman T, Beehner J (2004) Social system of a hybrid baboon group (Papio anubis × P. hamadryas). Int J Primatol 25(6):1313–1330
Biquand S, Biquqnd-Cuyot V, Boug A, Gautier J-P (1992a) The distribution of Papio hamadryas in Saudi Arabia: ecological correlates and human influence. Int J Primatol 13(3):223–243
Biquand S, Biquqnd-Cuyot V, Boug A, Gautier J-P (1992b) Group composition in wild and commensal hamadryas baboons: a comparative study in Saudi Arabia. Int J Primatol 13(5):533–543
Boug A, Biquand S, Biquand-Guyot, Kamal K (1994) The response of commensal hamadryas baboons to seasonal reduction in food provisioning. Rev Ecol (Terre et Vie) 49:307–319
Kummer H (1968) Social organization of hamadryas baboons. Karger, Basel
Kummer H, Banaja AA, Abo-Khatwa AN, Ghandour AM (1981) Mammals of Saudi Arabia. Primates. A survey of hamadryas baboons in Saudi Arabia. In: Wittmer W, Büttiker W (eds) Fauna of Saudi Arabia 3. Pro Entomologia/Natural History Museum, Basel, pp 441–471
Kummer H, Banaja AA, Abo-Khatwa AN, Ghandour AM (1985) Differences in social behavior between Ethiopian and Arabian hamadryas baboons. Folia Primatol 45:1–8
Mori A, Iwamoto T, Bekele A (1999) Sociological and demographic characteristics of a recently found Arsi gelada population in Ethiopia. Primates 40:365–381
Mori A, Gurja B, Iwaomoto T (2003) Changes in unit structures and infanticide observed in Arsi geladas. Primates 44:217–223
Nagel U (1973) A comparison of anubis baboons, hamadryas baboons and their hybrids at a species border in Ethiopia. Folia Primatol 19:104–163
Ohsawa H (1979) Herd dynamics. In: Kawai M (eds) Ecological and sociological studies of gelada baboons. Karger, Basel, pp 47–80
Ohsawa H, Dunbar RIM (1984) Variation in the demographic structure and dynamics of gelada populations. Behav Ecol Sociobiol 15:231–240
Pusey AE, Packer C (1987) Dispersal and philopatry. In: Smuts BB, Cheney DL, Seyfarth RM, Wrangham RW, Struhsaker TT (eds) Primate societies. The University of Chicago Press, Chicago, IL
Shotake T (1981) Population genetical study of natural hybridization between Papio anubis and P. hamadryas. Primates 22:285–308
Sigg H, Stolba A, Abegglen J-J, Dasser V (1982) Life history of hamadryas baboons: physical development, infant mortality, reproductive parameters and family relationships. Primates 23(4):473–487
Sugawara K (1979) Sociological study of wild group of hybrid baboons between Papio anubis and P. hamadryas in the Awash Valley Ethiopia. Primates 20:21–56
Sugawara K (1982) Sociological comparison between two wild groups of anubis-hamadryas hybrid baboons. Afr Study Monogr 2:73–131
Sugawara K (1988) Ethological study of the social behavior of hybrid baboons between Papio anubis and P. hamadryas in free-ranging groups. Primates 29(4):429–448
Swedell L (2002a). Ranging behavior, group size and behavioral flexibility in Ethiopian hamadryas baboons (Papio hamadryas hamadryas). Folia Primatol 73:95–103
Swedell L (2002b) Affiliation among females in wild hamadryas baboons (Papio hamadryas hamadrys). Int J Primatol 23:1205–1226
Wrangham R (1980) An ecological model of female-bonded primate groups. Behaviour 75:262–300
Yamane A, Shotake T, Mori A, Boug AI, Iwamoto T (2003) Extra-unit paternity of hamadryas baboons (Papio hamadryas) in Saudi Arabia. Ethol Ecol Evol 15:379–387
Zinner D, Pelaez F, Torkler F (2001a) Distribution and habitat associations of baboons (Paspio hamadryas) in central Eritrea. Int J Primatol 22:397–413
Zinner D, Pelaez F, Torkler F (2001b) Group composition and adult sex-ratio of hamadryas baboons (Papio hamadryas hamadryas) in central Eritrea. Int J Primatol 22:415–430
Acknowledgements
We thank the National Commission for Wildlife Conservation and Development (NCWCD) in Riyadh and the National Wildlife Research Center (NWRC) in Taif, Kingdom of Saudi Arabia, for giving us the opportunity to participate in this joint project. We are grateful to the staff of NWRC for their kind support, especially to Mr. Abdulrahman Khoja, the administrative director of NWRC. We thank Dr. Vanessa Hayes for comments and English correction of the manuscript. This study was partially funded by Grants-in-Aid for Scientific Research (No 14405005) from the Japan Society for the Promotion of Science (JSPS) and Grants-in-Aid for Scientific Research (No10041174) from the Ministry of Science, Sports, and Culture (Monbusho), Japan.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Mori, A., Yamane, A., Sugiura, H. et al. A study on the social structure and dispersal patterns of hamadryas baboons living in a commensal group at Taif, Saudi Arabia. Primates 48, 179–189 (2007). https://doi.org/10.1007/s10329-007-0045-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-007-0045-2