
Abstract
The efficiency of the indirect selection for wood density using the Pilodyn in Cryptomeria japonica was studied by comparing the Pilodyn penetration (PP) depth and the direct measurement of wood density in three test sites. The influence of the genotype by environment (G × E) interaction of wood density was estimated using the Pilodyn with common 12 C. japonica clones in 10 test sites in Kanto breeding region in Japan. The PP depth was highly correlated with wood density, and the genetic correlation between them was −0.88. The indirect selection using the Pilodyn realized 87% of the genetic gain obtained by the direct selection of wood density. The G × E interaction in PP depth was small. The ratio of the variance component of the G × E interaction to that of genotype was only 0.096 in the PP depth, whereas it was 0.700 in tree height and 1.410 in diameter at breast height. These findings indicate that the Pilodyn is useful for the genetic improvement of wood density in Cryptomeria japonica. The small G × E interaction in wood density estimated using the Pilodyn indicates that the relative clonal performance in wood density is stable among diverse environments in Kanto breeding region in Japan.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wood density is an important target trait in tree breeding programs because it is strongly correlated with other important wood properties, such as timber strength. Furthermore, wood density is an important determinant of the biomass accumulation capacity of each tree or genotype, which strongly affects forest carbon sequestration performance. Wood density is known to be under strong genetic control (Zobel and Jett 1995).
In tree breeding programs, a large number of measurements are required for the accurate selection of genotypes with superior performance in target traits and for the estimation of genetic parameters. The direct measurement of wood density is costly and time-consuming due to the need to prepare several specimens, in addition to harvesting trees or sampling with an increment borer. Thus, easier and inexpensive methods to evaluate the wood density of standing trees with minimal damage are required in the field of tree breeding. The Pilodyn, a tool originally developed to test for wood decay, is often used for the evaluation of wood density. The Pilodyn injects a pin into the wood with a constant force, and the depth of the penetration of the pin is measured. The measurement can be done very quickly, and damage to trees is minimal. There are many reports on the correlation between wood density and its indirect measurement based on the Pilodyn penetration (PP) depth of standing trees. Cown (1978) reported that the Pilodyn is a rapid method of measurement that provides a correlation coefficient of −0.96 with wood density on a clonal mean basis in Pinus radiata. Taylor (1981) also reported a high correlation coefficient (−0.81) between PP depth and wood density using 35 individual Pinus taeda. Hall (1988) reported correlation coefficients between the Pilodyn results and wood density of −0.48 in Picea mariana, −0.62 in Picea glauca and −0.48 in Larix laricina. In Japanese species, Yamashita et al. (2007) reported a correlation coefficient of −0.82 between penetration depth and the wood density of wood blocks from Cryptomeria japonica. Based on family means, the correlation coefficients are −0.79 in Cryptomeria japonica (Fukatsu et al. 2006) and −0.78 in Larix kaempferi (Fukatsu et al. 2007). Iki et al. (2009) reported a correlation coefficient of −0.769 in clonal mean basis in Abies sachalinensis. These reports, however, concentrated only on the estimation of correlation coefficients. A genetic correlation between the Pilodyn results and wood density is necessary for the selection of genetically superior trees using selection criteria such as best linear unbiased predictions or selection indices based on multiple traits (Falconer and Mackay 1996; White and Hodge 1989). Furthermore, the efficiency of the indirect selection of wood density using the Pilodyn must be evaluated based on the degree of the loss in genetic gain due to indirect measurement for tree breeding of Cryptomeria japonica.
The size of the genotype by environment (G × E) interaction is also an important consideration in tree breeding programs because of the difficulty in controlling environmental factors at planting sites (Shelbourne 1972). The relative performances of genotypes often vary with testing environments when there is a large G × E interaction. Two methods are used to reduce the effect of the G × E interaction in forest tree breeding: (1) selecting families that perform well across a wide range of conditions; and (2) regionalizing the breeding areas to sub-districts to decrease the extent of the G × E interaction. In Japan, such regionalization was first done based on climatic conditions in early stages of the forest tree breeding program. There are some reports on the G × E interaction in growth characteristics in Japanese main forestry species (Kurinobu 1984; Nishimura and Tajima 1993; Yamada and Miyaura 2002), and recently re-regionalization was suggested based on the height growth ranking in clonal tests of Cryptomeria japonica in the Kanto breeding region in Japan (Miura et al. 2005). The selection of the next generation is being performed not only by growth traits but also by wood properties. Evaluation of the G × E interaction in wood properties and comparison with that of growth traits is therefore important for the Japanese tree breeding program. There are few reports on the G × E interaction in wood density in Japan (Fujisawa et al. 1993; Akutsu et al. 2008). The G × E interaction for wood density in the Honshu main island of Japan has not yet been reported. The cause of the deficit in research of the G × E interaction is partly due to the cost of measuring wood properties. If the efficiency of the Pilodyn for estimating wood density is confirmed in Cryptomeria japonica, evaluation of the G × E interaction for wood density is achievable using the Pilodyn.
In this paper, we evaluate the efficiency of indirect selection for wood density in Cryptomeria japonica using the Pilodyn. In addition, we assess the extent of the G × E interaction for wood density in Cryptomeria japonica using the Pilodyn.
Materials and methods
Estimation of wood density using the Pilodyn
Clonal tests
Three clonal test sites of Cryptomeria japonica (sugi) used in this study were established in 1974 in the Kita-Kanto breeding district of the Kanto breeding region in Japan (sites 404, 405, and 450 in Fig. 1; Table 1). The field design of the three clonal tests was a randomized block design with three blocks. Clones were planted row plots of 10–45 ramets per block depending on the sites. These sites were composed of clones selected mainly from the Kita-Kanto breeding district and others from the other three districts of the Kanto breeding region.

The test sites used in this study. The numbers show test site codes in the Forest Tree Breeding Center, Forestry and Forest Product Research Institute. The test site numbers within the oval outlines are the test sites used in the analysis for genotype by environment interaction. The shaded area shows the Kanto breeding region and degree of shading shows different breeding districts
Measurement of wood density, diameter, and height
For the measurement of wood density, we sampled discs from 31 clones at site No. 404, 12 clones at site No. 405 and 15 clones at site No. 450, at 1.2 m above the ground. The discs were obtained from the thinning of the clonal test sites. The selection of thinned trees were planned to keep the stem density of the test sites would be suitable for the forest management in the future. The survival rates were different between clones and the harvested trees were not always suitable for the sampling for density measurement. Therefore, the number of trees that were thinned and sampled was different between plots. One to six individuals (mean 2.8) were sampled per plot. Diameters at the breast height (DBH) and tree height were measured on the sampled trees. The mean number of common clones between sites was 4.6. We analyzed each site individually because of the small numbers of common clones across the sites. In total, 332 individuals were sampled. Test specimens (3 cm tangential, 4 cm axial, and arbitrary length in the radial direction depending on the diameter of each tree) were cut from the heartwood and sapwood from two directions in the minor diameter axis. The wood density (basic density) of the specimens was measured from the water saturated volume and the oven-dried weight of the specimens. The density of sapwood (sapwood density) and that of heartwood (heartwood density) were calculated by averaging the values of two radii as representatives of two wood types of an individual tree. Disc density, representing the density of a whole disc sampled from breast heights of each tree, was calculated for each individual as an area-based weighted mean of the wood density of heartwood and sapwood. Heritability and genetic correlations with growth traits of the wood density of these materials were reported by Tamura et al. (2006).
Measurement of Pilodyn penetration depth
Pilodyn penetration depth was measured at the same plot, on the different individuals from the individuals sampled for the measurement of wood density. Three individuals were measured for each plot. The ages at the time of measurement differed between PP and wood density (Table 1). PP was measured using a Pilodyn 6J Forest (PROCEQ, Zurich, Switzerland) with a 2.5-mm diameter pin without removing the bark, from two directions at right angles to the direction of the slope, two PP readings per one direction. The mean value obtained from four Pilodyn shots was used as the PP for each individual tree.
Estimation of variance components
The variance components were calculated based on the following model:
where Y ijk is the value of an individual k of a clone j in a block i; μ is the general mean; B i is the fixed effect of a block i; c j is the random effect of a clone j; p ij is the random plot error existing in a clone j in a block i; and e ijk is the random error of an individual k in a clone j in a block i.
All the variance components for the random effects in this paper were calculated based on each equation using the restricted maximum likelihood (REML) method with the SAS VARCOMP procedure (SAS Institute Inc.).
Estimation of heritability and genetic correlation
The broad sense heritability of the clonal mean and individuals for each trait were calculated using the following formulas:
where h 2c and h 2i represent the broad sense heritability of the clonal mean and the individuals, respectively; σ 2c is the clonal variance; σ 2p is the variance of plot error; σ 2e is the variance of error; t is the number of blocks; and b is the number of the individuals in each plot. Clonal covariance components between trait x and trait y were estimated using the following formula (White and Hodge 1989; Xiang et al. 2003):
where Covc(x,y) is the clonal covariance between trait x and trait y; σ 2c(x+y) is the clonal variance of the sum value of trait x and trait y; σ 2c(x) is the clonal variance of the trait x; and σ 2c(y) is the clonal variance of trait y. The genetic correlation between two traits (x, y) was calculated as follows:
where r g(x,y) is the genetic correlation between two traits.
Genetic gain by the direct selection and the indirect selection
To evaluate the efficiency of the indirect selection for wood density using Pilodyn, the genetic gains by the direct and indirect selections were estimated. The genetic gain by the direct selection, as the response of selection for trait x by the same trait x was calculated as follows:
where ΔG x is the genetic gain of trait x; i is the intensity of selection; h c(x) is the square root of the clonal heritability for trait x; and σ c(x) is standard deviation of the clonal effect. Genetic gain as the response of indirect selection for trait x by trait y was estimated using the following formula:
where ΔG x(y) is the genetic gain of trait x by the selection of y, and h c(y) is the clonal heritability for trait y. Relative genetic gain was calculated as the genetic gain divided by the mean value of the original population (site mean) for each site. In this paper, we used 1 for i, which corresponds to selecting 37% of the upper portion (better clones from the original population) (Falconer and Mackay 1996).
Analysis of the G × E interaction in wood density
Clonal tests
To estimate the G × E interaction, we used 10 clonal test sites composed of 12 common clones selected from the Kanto breeding region. The locations and characteristics of the clonal test sites are shown in Fig. 1 and Table 1. The investigated clones are listed in Table 2. The Kanto breeding region is composed of the Metropolis of Tokyo and 12 prefectures. The 12 clones investigated in this study were originally selected from the 12 prefectures in the Kanto breeding region. There are a few untested clones in some sites; the mean number of clone per sites was 11.5. The field designs of sites were as described in “Estimation of wood density using the Pilodyn”.
Measurement of Pilodyn penetration depth, diameter, and height
PP was measured using the same method used in “Estimation of wood density using the Pilodyn”. Basically, three to six trees (5.2 trees on average) were measured for PP in each plot, depending on the sites. DBHs and tree heights of all surviving and undamaged trees were measured (25.4 trees per plot on average). In a few plots, where survival rates were unusually low, we could measure only one or two trees per plot for the three traits. Stand ages at time of the measurements differed between the PP and growth traits (Table 1). Inter-site phenotypic correlations for all combinations of the 10 sites were calculated based on the clonal means of each trial.
Analysis of the variance component
The model used for the analysis of variance components for each trait was:
where Y ijk is the plot mean for a clone k in a block j in a site i; μ is the general mean; E i is the fixed effect of a site i; B ij is the fixed effect of a block j in a site i; c k is the random effect of a clone k; ce ik is the interaction effect between a clone k and a site i (G × E interaction); and e ijk is the random error of a clone k in a block j of a site i. The difference in stand age at the time of the measurements between sites (Table 1) was incorporated into the difference in the environment between sites in our analysis because the difference in age between sites was not large. The variance components in Eq. 8 were calculated for two groups of test sites; all 10 test sites investigated, and the 8 test sites belonging to the Taiheiyo breeding distinct (Table 1) proposed by Miura et al. (2005). The genetic correlations between traits were calculated using Eq. 4. The ratio of the variance of the G × E interaction to clonal variance (GEI/G) was calculated for each trait as an index of the extent of the G × E interaction (Shelbourne 1972).
Results
Selection of wood density using the Pilodyn
Clear correlations were observed between PP and wood density in the clonal mean of each site (Fig. 2). PP shows a higher correlation with sapwood density (r = −0.848 on average for three test sites) than heartwood density (r = −0.699). The PP-disc density correlation was intermediate (r = −0.837) between PP-heartwood density and PP-sapwood density. The broad sense heritability of individuals (h 2i ) of both PP (0.71) and disc density (0.76) was higher than that of growth traits (0.49 for height and 0.33 for DBH) (Table 3). Genetic correlations between PP and wood densities showed high values (Table 3). The highest genetic correlation was observed between sapwood density and PP. The standard deviations of genetic correlations between sites were very small between PP and wood density traits.

Relationship between Pilodyn penetration depth and wood density. The correlation between Pilodyn penetration depth and sapwood density (a), heartwood density (b), and disc density (c). The least square mean (Littell et al. 1991) of each clone of each test site was plotted. Different symbols show the samples from different test sites. PP Pilodyn penetration depth. n = 31 in site 404, 12 in site 405, 15 in site 450
The genetic gain in disc density was 0.0219 g/cm3 for an average of three sites by the direct selection of the top 37% of the clones (i = 1) (Fig. 3). The relative genetic gain was 7.47% in disc density. Indirect selection by the PP shows the genetic gain of 0.0191 g/cm3 in disc density by the same selection intensity. Indirect selection showed a loss of genetic gain of −12.8%. The loss of genetic gain by the indirect selection using the PP data was largest (−19.7%) in heartwood density and smallest (−9.45%) in sapwood density.

Genetic gain in wood density by the direct selection and by the indirect selection using the Pilodyn. The predicted genetic gain for the selection of the top 37% clones (i = 1). Black bar and white bar indicate the results from the direct selection and the indirect selection using the Pilodyn, respectively. DS Sapwood density, DH heartwood density, DD disc density
The G × E interaction in the wood density estimated by the Pilodyn
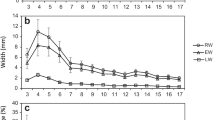
Broad-sense heritabilities based on the average value of the plots were 0.797 for the Pilodyn, 0.150 in DBH, and 0.182 in height, across the 10 test sites. The compositions of the variance components are shown in Fig. 4. The ratios of the G × E interaction variance to clonal variance (GEI/G) were 0.096 for the Pilodyn, 0.700 for the height and 1.420 for the DBH. The correlation coefficients between each test site for each trait and genetic correlation between traits is shown in Table 4. The strongest correlation was observed in PP (r = 0.888 on average). Low genetic correlations were observed between PP and height (−0.093) and between PP and DBH (0.262).

The proportion of variance components. The proportion of clonal variance, G × E interaction variance and error variance to total variance is shown for each investigated trait. All 10 test sites were used. PP Pilodyn penetration value, DBH diameter at breast height, H height
When the GEI/G across all the test sites and the GEI/G across only the 8 test sites belonging to the Taiheiyo breeding distinct (shown in Table 1 as T) proposed by Miura et al. (2005) were compared, the GEI/G decreased from 0.700 to 0.244 in height and from 1.420 to 0.630 in DBH by the regionalization, whereas the GEI/G in the PP changed only slightly from 0.096 to 0.105.
Discussion
The findings of the present study indicates a high correlation between wood density and PP in Cryptomeria japonica. The correlation was stronger for sapwood density than for heartwood density, which was consistent with the fact that the Pilodyn pin penetrated into only the outside (only a few centimeters from the bark; Table 2) of the trunk of a tree when PP was measured on a standing tree. The correlation between PP and either heartwood density or disc density was, nevertheless, moderately high. Several studies reported a high correlation between PP and wood density (Cown 1978; Taylor 1981; Hall 1988; Yamashita et al. 2007; Fukatsu et al. 2007; Iki et al. 2009). Our findings also indicate a high correlation between PP and wood density consistent with the previous reports. The small loss of genetic gain (−12.8%) by the indirect selection of disc density using the Pilodyn indicates that the Pilodyn measurement is effective for the selection of clones with higher wood density. We conclude that we can improve wood density via the indirect selection using the Pilodyn in Cryptomeria japonica. In addition, the great efficiency for the estimation of wood density, which has been revealed by the high correlation with wood density, and the rapidity of the measurement of PP depth indicate that the Pilodyn enable the evaluation the G × E interaction in wood density.
The G × E interaction was small in PP among the sites investigated (Fig. 4; Table 4), suggesting that the interaction was also small in wood density because the Pilodyn was confirmed to be efficient for the estimation of wood density in this study. Reports on the G × E interaction in wood density are limited, especially in Japan. Fujisawa et al. (1993) reported a small interaction Cryptomeria japonica clones and test sites in Kyushu Island. Akutsu et al. (2008) reported that the G × E interaction was significant in wood density, and suggested that regionalization of seed distribution zones was necessary for Abies sachalinensis in Hokkaido Island, Japan. In other countries, Lima et al. (2000) reported that rank correlations between sites were all significant in Eucalyptus hybrids using 4 test sites. McKeand et al. (1997) showed that the variance in the G × E interaction is relatively small for wood density compared with growth traits in Pinus taeda. Osorio et al. (2001) and Baltunis et al. (2007) also reported that the G × E interaction for wood density is small in Eucalyptus grandis in Colombia and in Pinus radiata in Australia, respectively. Our result was similar to the previous reports except for that by Akutsu et al. (2008).
Miura et al. (2005) reported that the Kanto breeding region in Japan should be divided into two breeding districts based on the analysis of height growth using the test sites, including the sites used in this paper. The G × E interaction in PP was small compared with growth traits in Cryptomeria japonica in the Kanto breeding region as revealed in this research. Shelbourne (1972) stated that, as an approximate rule of thumb, the effect of the G × E interaction would be serious for selection when GEI/G reaches 0.5 or more. The regionalization of the breeding region is more important for growth traits than wood density, considering the extent of the G × E interaction and GEI/G for each trait in this area. When the regionalization proposed by Miura et al. (2005) was considered, the GEI/G was greatly decreased in growth traits, whereas GEI/G was stable in the Pilodyn measurements. This finding also indicates that the regionalization of breeding region is relatively more important for growth traits than wood density. The target traits in the breeding of Cryptomeria japonica are multiple, including growth traits, wood quality, resistance to pests, etc. From our results, when regionalization of a breeding region is done based on the growth traits, the genetic improvement of wood density can proceed without problem with the G × E interaction. The low genetic correlations between PP and growth traits (Table 4) indicates that simultaneous genetic improvement of both traits is possible.
Thus, the findings of the present study indicate the efficiency of the Pilodyn measurements for the indirect estimation of wood density, and the usefulness of the Pilodyn for the selection of clones with higher wood density. The small G × E interaction in wood density estimated by the Pilodyn indicates that the relative performances of clones in wood density are stable among diverse environments in Kanto breeding region, and that the selection of clones with a higher wood density is practicable according to the regionalized breeding area based on the growth characteristics of the Cryptomeria japonica in the Kanto breeding region in Japan.
References
Akutsu H, Kita K, Uchiyama K, Kuromaru M (2008) Genetic variation and genotype-environment interaction in annual ring structure in progeny test of plus tree of Sakhalin Fir (Abies sachalinensis). J Jpn For Soc 90:137–144 (in Japanese)
Baltunis BS, Wu HX, Powell MB (2007) Inheritance of density, microfibril angle, and modulus of elasticity in juvenile wood of Pinus radiata at two locations in Australia. Can J For Res 37:2164–2174
Cown DJ (1978) Comparison of the Pilodyn and torsiometer methods for the rapid assessment of wood density in living trees. N Z J For Sci 8:384–391
Falconer DS, Mackay TFC (1996) Introduction to quantitative genetics, 4th edn. Pearson, Harlow
Fujisawa Y, Ohta S, Tajima M (1993) Wood characteristics and genetic variations in sugi (Cryptomeria japonica) II. Variation in growth ring components among plus-trees clones and test stands. Mokuzai Gakkaishi 39:875–882
Fukatsu E, Higashihara T, Nakada R (2006) Analysis of variation and heritability of wood density in families by controlled mating in Cryptomeria japonica. Kanto J For Res 58:83–86 (in Japanese)
Fukatsu E, Nakada R, Tsubomura M, Kubota M (2007) Genetic parameters for traits related to carbon-sequestration from a full-diallel mating design in Japanese larch (Larix kaempferi). In: Perron M (ed) Proceedings of Larix 2007: international symposium of the IUFRO working group S2.02.07. Ministère des Ressources naturelles et de la Faune, Québec, pp 89–93
Hall JP (1988) Using the Pilodyn tester in selecting plus trees. North J Appl For 5:170–171
Iki T, Tamura A, Nishioka N, Abe M, Iizuka K (2009) Longitudinal changes of basic density and non-destructive quality evaluation using the Pilodyn in todomatsu (Abies sachalinensis) plus tree clones. Mokuzai Gakkaishi 55:18–28 (in Japanese)
Kurinobu S (1984) A methodological study of the analysis of progeny trial plantations of Japanese larch. Bull For Tree Breed Inst 2:1–60 (in Japanese)
Lima JT, Breese MC, Cahalan CM (2000) Genotype-environment interaction in wood basic density of Eucalyptus clones. Wood Sci Technol 34:197–206
Littell RC, Freund RJ, Spector PC (1991) SAS system for the linear models, third edition. SAS Institute, Cary
McKeand SE, Eriksson G, Roberds JH (1997) Genotype by environment interaction for index traits that combine growth and wood density in loblolly pine. Theor Appl Genet 94:1015–1022
Miura M, Kubota M, Nomura T, Kurinobu S (2005) Investigation of regionalization in the Kanto breeding region based on data gained from the clonal tests on 20 years-old sugi (Cryptomeria japonica). J Jpn For Soc 87:233–240 (in Japanese)
Nishimura K, Tajima M (1993) Classification of tree breeding districts in the Kyushu region by the growth of sugi (Cryptomeria japonica D. Don) plus trees. J Jpn For Soc 75:493–500 (in Japanese)
Osorio LF, White TL, Huber DA (2001) Age trends of heritabilities and genotype-by-environment interactions for growth traits and wood density from clonal trials of Eucalyptus grandis Hill ex Maiden. Silvae Genet 50:108–117
Shelbourne CJA (1972) Genotype-environment interactions: its study and its implications in forest tree improvement. In: IUFRO genetics-SABRAO joint symposia B-1, Tokyo, pp 1–28
Tamura A, Kurinobu S, Fukatsu E, Iizuka K (2006) An investigation on the allocation of selection weight on growth and wood basic density to maximize carbon storage in the stem of sugi (Cryptomeria japonica D. Don) plus-tree clones. J Jpn For Soc 88:15–20 (in Japanese)
Taylor FW (1981) Rapid-determination of southern pine specific gravity with a Pilodyn tester. For Sci 27:59–61
White TL, Hodge GR (1989) Predicting breeding values with applications in forest tree improvement. Kluwer, Dordrecht
Xiang B, Li BL, McKeand S (2003) Genetic gain and selection efficiency of loblolly pine in three geographic regions. For Sci 49:196–208
Yamada H, Miyaura T (2002) Relationships between stand productivity and height growth for clonal lines and open-pollinated progenies of sugi (Cryptomeria japonica D. Don) plus trees selected from Kansai forest tree breeding region. J Jpn For Soc 84:50–53 (in Japanese)
Yamashita K, Okada N, Fujiwara T (2007) Use of the Pilodyn for estimating basic density and its applicability to density-based classifying of Cryptomeria japonica green logs. Mokuzai Gakkaishi 53:72–81 (in Japanese)
Zobel BJ, Jett JB (1995) Genetics of wood production. Springer, Berlin
Acknowledgments
The clonal test sites used in this study were established and maintained by the National Forest of Japan in collaboration with Forest Tree Breeding Center, Forest and Forest Product Research Institute. We thank the members of these organizations for their effort. We are especially grateful to Dr. Masahiro Miura, Forest Tree Breeding Center, for his useful advice regarding this study.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Fukatsu, E., Tamura, A., Takahashi, M. et al. Efficiency of the indirect selection and the evaluation of the genotype by environment interaction using Pilodyn for the genetic improvement of wood density in Cryptomeria japonica . J For Res 16, 128–135 (2011). https://doi.org/10.1007/s10310-010-0217-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10310-010-0217-6