
Abstract
Accumulation of proline is trusted to be an adaptive response of plants against drought stress, and exogenous application of nitric oxide (NO) enhances proline accumulation in Cu-treated algae. In order to investigate whether NO works as a necessary signaling molecule in drought-induced proline accumulation in rice leaves, effects of drought stress on endogenous NO content and proline accumulation were studied in rice leaves, using sodium nitroprusside (SNP, a NO donor) and 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (cPTIO, a NO scavenger). The results showed that drought treatment increased both endogenous NO and proline contents in rice leaves, while foliar spray of various concentrations of SNP failed to induce proline accumulation in the leaves of well-watered rice and foliar spray of cPTIO failed to inhibit proline accumulation in the leaves of drought-stressed rice. These results indicate that increase of endogenous NO is dispensable for proline accumulation in the leaves of rice under drought stress. Further studies indicate that exogenous application of NO alleviates drought-induced water loss and ion leakage by decreasing transpiration rate of rice leaves.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
As one of the most pivotal factors limiting crop production in many regions of the world, drought stress causes several seriously deleterious effects on plant metabolic processes including water relations, nutrient uptake and metabolism, and photosynthetic assimilates (Zhang and Kirkham 1994; Shinozaki et al. 2003). Plants have evolved a wide series of defense mechanisms to increase tolerance capacity during water deficit and other osmotic stresses, such as inducing stomatal closure and decreasing transpiration (García-Mata and Lamattina 2001). A strategy that may confer drought tolerance is the activation of antioxidative enzymes and accumulation of antioxidants (Loggini et al. 1999; Fu and Huang 2001; Luna et al. 2005). Another strategy is the rapid accumulation of compatible, low-molecular-weight osmolytes, such as sugar alcohols, special amino acids, and Gly-betaine (Munns 2002).
As a crucial gaseous signaling molecule in plants, nitric oxide (NO) plays significant roles in modulating several physiological and biochemical functions in different plant tissues, from germination to flowering, and until senescence (Lamattina et al. 2003; Crawford and Guo 2005). Recently, increasing instances suggest that NO is involved in the plant response to biotic and abiotic stresses such as pathogen, chilling, salt, heavy metals and drought, and a number of articles have reported the roles of exogenously applied NO in alleviating abiotic stresses in plants (Qiao and Fan 2008; Xiong et al. 2010). Studies suggest that exogenous application of NO donors might induce stomatal closure and enhance the adaptive responses against drought stress in Triticum aestivum (García-Mata and Lamattina 2001). Recently, experimental evidence indicates that exogenously applied NO alleviates osmotic stress by decreasing oxidative damage and stimulating proline accumulation in wheat (Tan et al. 2008). Moreover, exogenous application of NO enhanced drought tolerance in Cucumis sativus, Zea mays, and Oryza sativa are suggested primarily with increased antioxidants, which scavenge reactive oxygen species (ROS), improve cellular membranes stability and maintain photosynthesis and water status (Hao et al. 2008; Arasimowicz-Jelonek et al. 2009; Farooq et al. 2009). Recently, it is reported that NO dependence on nitric oxide synthase-like activity serves as a signaling component in the induction of protective responses and is associated with drought tolerance in maize seedlings (Hao et al. 2008). So far, exogenous application of NO to drought-stressed plants has facilitated obtaining enhanced plant growth and maintenance of a higher relative water content (RWC) (Tian and Lei 2006; Lei et al. 2007; Arasimowicz-Jelonek et al. 2009). However, reports on the effects of exogenous NO on drought stress is still limited, and current understanding of the molecular and physiological mechanisms of NO in alleviating drought stress is limited too.
Free proline is not only an osmoprotectant, but it might also eliminate ROS and promote the antioxidant ability, stabilize the structure of biological macromolecules, decrease cell activity and relieve the toxicity of NH4 + (Taylor 1996). Accumulation of proline is trusted to be an adaptive response of plants against environmental stresses such as heavy metal, high salinity, light-induced stress, and drought stress (Siripornadulsil et al. 2002; Zhang et al. 2008). Exogenously applied NO not only promotes free proline accumulation in annual ryegrass under chilling stress (Ma et al. 2005), but also improves leaf-free proline content in wheat and rice under drought stress (Lei et al. 2007; Tan et al. 2008; Farooq et al. 2009). A few authors report that exogenous application of NO enhances drought tolerance in plants is partly involved with the accumulation of free proline in plants (Tan et al. 2008; Farooq et al. 2009). In contrast, López-Carrión et al. (2008) show that exogenous application of NO decreases saline-induced proline accumulation in Brassica rapa. Until now, the relationships between endogenous NO increase and proline accumulation in rice leaves is still unclear. Moreover, the effects of drought stress on endogenous NO content in rice leaves require further researches too.
Recently, Zhang et al. (2008) reported that copper-induced proline synthesis was associated with NO generation in Chlamydomonas reinhardtii, while exogenous NO failed to induce proline accumulation in the absence of copper. In order to investigate whether drought-induced proline accumulation is associated with NO generation in rice leaves, this study aims to investigate the effects of drought stress on endogenous NO content and proline accumulation in rice leaves. We also will evaluate the effect of exogenous application or scavenging of NO on proline content in rice leaves. The outcome of this work may help to understand the mechanisms and regulatory pathways of NO in alleviating drought stress in crucial crops such as rice.
Materials and methods
Plant cultivation and drought treatment
The experiments were conducted at the experimental base of China National Rice Research Institute (CNRRI) in Fuyang city of China (30°03′N, 119°57′E). Healthy rice seeds (Oryza sativa L., cv. Nipponbare) were sterilized with 5% sodium hypochlorite for 15 min, and then the seeds were rinsed 3 times with sterilized water and incubated for 48 h at 37°C. Uniformly germinated seeds were transferred to a net tray floating on container filled with a half-strength Yoshida rice nutrient solution for 2 weeks; the nutrient solution was refreshed every 4 days. Then, three uniform rice seedlings were transplanted into a 12 l polyvinyl chloride pot (30 cm in diameter and 40 cm in height) filled with 15 kg submerged puddle gleyic stagnic anthrosol, and 1 g urea was mixed into each pot 1 day before transplanting. The soil was taken from the experimental paddy field at CNRRI farm that was plowed and harrowed 7 days ago, the pH (H2O) of the topsoil (0–15 cm) was 6.0, and the soil contained 28.6 g kg−1 organic matter, 1.88 g kg−1 total N, and 15.6 cmol kg−1 CEC. All experiments were conducted in a greenhouse with 30/24°C (day/night) thermoperiod, 80% relative humidity and natural photoperiod (about 14/10 h, day/night).
When the rice seedlings were 6 weeks old, drought was imposed by withholding water application and draining off water from holes in the bottom of the polyvinyl chloride pots, the duration of drought was 15 days.
Determination of soil water potential and water content
A negative pressure tensiometer (produced by Institute of Soil Science, Chinese Academy of Sciences) was inserted into the soil for 15 cm depth, and the soil water potential was recorded every day. Soil from 15 cm depth was dried for 24 h at 105°C and water content in the soil was directly determined using the difference in weight before and after drying a soil sample, the water content was recorded every day. Water content and water potential in the soil during drought duration were shown in Fig. 1.

Water content (a) and water potential (b) in the soil during drought treatment. The value is the average of 5 repeats each, vertical bar represents SE
Chemicals and treatment
Sodium nitroprusside (SNP; Fluka, Germany) was used as NO donor, and 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (cPTIO; Sigma, St Louis, MO, USA) was used as NO scavenger. Uniform 6-weeks-old rice plants were used in the experiments, a half of the plants were drought stressed by withholding water application and by draining off water from holes in the bottom of polyvinyl chloride pots, and another half of the plants were well watered as control. From the first day of drought treatment, each plant was sprayed with 50 ml of SNP, cPTIO or SNP + cPTIO (0, 10, 50, 100, 200 and 500 μM) every 2 days. Plants were sprayed with a concentration of 100 μM SNP or/and cPTIO mainly, the wide range of 0–500 μM was only used for concentration-dependent proline measurement.
Determination of NO content
NO determination was performed by fluorometric approach following the method of Zhao et al. (2009) with slight modification. For NO visualization in rice leaf, 2 cm segments of the penultimate leaf were excised and incubated with 2.5 μM NO-specific fluorescent probe 4-amino-5-methylamino-2′,7′-difluorofluorescein diacetate (DAF-FM DA; Invitrogen, Carlsbad, CA, USA) for 30 min. DAF-FM DA was prepared in HEPES–NaOH buffer (pH 7.5), therefore, incubated leaves were washed 3 times with the same buffer for 10 min each. Leaf segments were then soaked in glycerol: HEPES–NaOH buffer (1:1 v/v) and mounted in the same medium for examination with a confocal laser scanning microscope system (LSM 510; Zeiss, Oberkochen, Germany), the fluorescence was imaged with excitation and emission wavelengths of 488 and 515 nm, respectively. The three-dimensional reconstructed image of the leaf was used to calculate the relative fluorescence intensity, and the fluorescence intensity was expressed in color level on a scale ranging from 0 to 255.
Determination of leaf relative water content
To determine RWC, fresh rice leaves were weighed immediately to get fresh weight (FW), then these leaves were soaked in deionized water for 2 h and saturated weight (SW) was determined, these leaves were dried for 24 h at 75°C to determine dry weight (DW). RWC was calculated as:
Determination of leaf water potential
A water potential system (PSYPRO; Wescor Inc., Logan, UT, USA) was used for leaf water potential measurement. For each treatment, eight rice plants were chosen randomly and the penultimate leaves were used for measurement.
Determination of leaf ion leakage
After treatment, 1 g fresh rice leaves were cut into 25-mm2 pieces and engrossed in a test tube with 12 ml deionized water for 2 h at 25°C, after the incubation, the electrical conductivity of solution (EC1) was determined with a conductivity meter (DDA-11A; Shanghai Hongyi Instrument Co. Ltd., Shanghai, China). Samples were then heated at 80°C for 2 h in their effusates and cooled to 25°C, and the electrical conductivity (EC2) was measured again in the bathing solution. Ion leakage was calculated as the ratio between EC1 and EC2.
Determination of leaf proline content
Proline content was determined following the method of Bates et al. (1973) with slight modification. 0.25 g fresh rice leaves were frozen with liquid N2 and then ground into powder, 5 ml of 3% aqueous sulphosalicyclic acid was added into the powder. The homogenate was centrifuged at 13,000g for 10 min at 4°C, 2 ml supernatant was mixed with 2 ml acid-ninhydrin and 2 ml glacial acetic acid in a test tube. The mixture was heated in a water bath at 100°C for 1 h, then the reaction mixture was extracted with 4 ml toluene and the toluene containing chromophore was aspirated, cooled to 25°C and absorbance was measured at 520 nm with a UV/vis spectrophotometer (Lambda 25; PerkinElmer, MA, USA).
Determination of transpiration
Transpiration assay was done with a portable photosynthesis system (LI-6400, LICOR, Lincoln, NB, USA). All the measurements were taken at a constant airflow rate at 500 μmol s−1 and saturation irradiance with incident photosynthetic photo flux density (PPED) of 1,200 μmol m−2 s−1. The ambient CO2 concentration was approximately 400 cm3 m−3 and the temperature was 30°C. Data were recorded hour after foliar spray of water, SNP, cPTIO or SNP + cPTIO.
Statistical analysis
Each experiment was repeated 3 times independently, values were expressed as mean ± SE. Data were analyzed by analysis of variance (ANOVA) and difference test (LSD) was employed to determine the differences among the treatments at P = 0.05 level.
Results
Effects of drought, SNP and cPTIO treatment on NO content in rice leaf
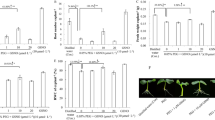
The results of our pre-experiments showed that 100 μM SNP or cPTIO had no obvious promoting or inhibiting effect on the growth performance of well-watered rice, and furthermore, 100 μM SNP had the most significant alleviation effect on growth performance of drought-treated rice and a concentration of 100 μM cPTIO had been widely used as a NO scavenger in many plants. Therefore, a concentration of 100 μM SNP or/and cPTIO was used in subsequent experiment mainly. In order to monitor the effect of drought stress on endogenous NO content in rice leaves accurately, NO content was detected visually with the NO-specific fluorescent probe DAF-FM DA in a time-course manner (Figs. 2, 3). In the penultimate leaf of well-watered rice, the fluorescence intensity of NO was faint and had no obvious variation in relation to duration (Fig. 2). However, in the drought-stressed rice, the fluorescence intensity of NO in penultimate rice leaf increased significantly with the duration of drought. The peak level of fluorescence intensity was observed at 9 days of drought, with the intensity being approximately 3-fold of the control. Following that period, the level of fluorescence intensity declined gradually (Fig. 3). These observations reveal that drought stress significantly increase endogenous NO content in rice leaves, with time-dependent changes.

Effects of foliar spray of 100 μM SNP or/and cPTIO on NO content in the leaves of well-watered rice. a Visualization of NO-specific fluorescence in the leaves of well-watered rice. b Time-course of NO-specific fluorescence density in the leaves of well-watered rice. The value is the average of 5 leaves each, vertical bar represents SE of each treatment. Asterisk means significant difference compared with control at the same day

Effects of foliar spray of 100 μM SNP or/and cPTIO on NO content in the leaves of drought-treated rice. a Visualization of NO-specific fluorescence in the leaves of drought-treated rice. b Time-course of NO-specific fluorescence density in the leaves of drought-treated rice. The value is the average of 5 leaves each, vertical bar represents SE of each treatment. Asterisk means significant difference compared with control at the same day
As shown in Figs. 2 and 3, the effects of foliar spray of SNP or/and cPTIO on endogenous NO in rice leaves were investigated, foliar spray of 100 μM SNP significantly increased NO content in the leaves of both well-watered and drought-treated rice. In contrast, foliar spray of 100 μM cPTIO significantly decreased NO content in the leaves of both well-watered and drought-treated rice. Simultaneously foliar spray of 100 μM SNP and 100 μM cPTIO slightly increased NO content in the leaves of well-watered rice. However, simultaneously foliar spray of 100 μM SNP and 100 μM cPTIO inhibited drought-induced NO increase in the leaves of drought-treated rice, the NO content maintained at a steady level.
Effects of drought stress and NO on proline accumulation in rice leaf
To understand the pattern of drought-induced proline accumulation in rice leaves, an initial time-course experiment was performed (Fig. 4). In the leaves of well-watered rice, the proline level had no obvious change with the duration (Fig. 4a). However, in the leaves of drought-stressed rice, there was a rapid accumulation of proline in the time-course experiment. The proline level increased by 5-fold after 12 days of drought stress, and following that period, the proline level decreased gradually (Fig. 4b).

Effects of foliar spray of 100 μM SNP or/and cPTIO on proline content in the leaves of rice. a In the leaves of well-watered rice. b In the leaves of drought-treated rice. The value is the average of 8 leaves each, vertical bar represents SE of each treatment. Asterisk means significant difference compared with control at the same day
In order to investigate the role of NO in drought-induced accumulation of proline in rice leaves, an exogenous NO donor SNP and an exogenous NO scavenger cPTIO were applied in the study (Fig. 4). Interestingly, foliar spray of 100 μM SNP had no significant effect on proline level in the leaves of well-watered rice, and the proline content remained unchanged with the duration (Fig. 4a). Similarly, foliar spray of 100 μM cPTIO had no effect on proline level in the leaves of well-watered rice. At the same time, foliar spray of 100 μM SNP also had no effect on the proline content in the leaves of well-watered rice (Fig. 4a). Foliar spray of 100 μM cPTIO also failed to decrease proline accumulation in the leaves of drought-stressed rice (Fig. 4b). Figure 4 also showed that simultaneous spray of 100 μM SNP and 100 μM cPTIO had no effect on proline level in the leaves of well-watered and drought-stressed rice. These results reveal that exogenous application of NO possible has no effect on proline accumulation in the leaves of both well-watered and drought-stressed rice. These results indicate that increase of endogenous NO has no relationship with proline accumulation in the leaves of both well-watered and drought-stressed rice.
To exclude the possibility that the concentrations of 100 μM SNP or (and) 100 μM cPTIO were not suitable to affect proline accumulation in rice leaves, concentration-course studies of SNP (Fig. 5a) and cPTIO (Fig. 5b) also were performed. These results showed that treatments with 10, 50, 100, 200 and 500 μM SNP or cPTIO hardly affected proline levels in the leaves of well-watered and drought-stressed rice, as compared with the controls. Taken together, our results demonstrate that neither exogenously applied NO nor endogenous NO affect proline accumulation in the leaves of rice under drought-stressed or well-watered condition.

Effects of different concentrations of foliar spray of SNP or cPTIO on proline content in the leaves of well-watered and drought-stressed rice. a Treatment with 0, 10, 50, 100, 200 and 500 μM SNP on proline content in rice leaves. b Treatment with 0, 10, 50, 100, 200 and 500 μM cPTIO on proline content in rice leaves. The proline content was determined after 12 days drought treatment. The value is the average of 8 leaves each, vertical bar represents SE of each treatment. Different letters indicate significant differences at P = 0.05
Effect of NO in alleviating drought-caused water loss and ion leakage
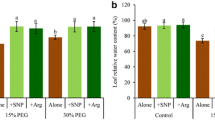
Farooq et al. (2009) reported that exogenous application of NO significantly increased free proline content in the leaves of rice under drought stress and then enhanced rice drought tolerance. In contrast, our results indicate that although drought induces both proline and NO accumulations in rice leaves, exogenous application of NO hardly affected proline accumulation in the leaves of well-watered and drought-stressed rice. In order to evaluate the effect of exogenously applied NO in enhancing rice drought tolerance, SNP and cPTIO were foliar sprayed to investigate the effect of NO on water loss and ion leakage in rice leaves under drought stress. Figure 6a shows that foliar spray of 100 μM SNP, 100 μM cPTIO or 100 μM SNP + cPTIO hardly had effect on the RWC in the leaves of well-watered rice, while foliar spray of 100 μM SNP significantly alleviated drought-induced decrease of RWC in the leaves of drought-stressed rice (Fig. 6b). Figure 6c shows that foliar spray of 100 μM SNP, 100 μM cPTIO or 100 μM SNP + cPTIO hardly had effect on the water potential in the leaves of well-watered rice, while foliar spray of 100 μM SNP significantly alleviated drought-induced decrease of water potential in the leaves rice (Fig. 6d). Similarly, foliar spray of SNP, cPTIO or SNP + cPTIO hardly had effect on the ion leakage in the leaves of well-watered rice (Fig. 6e), while foliar spray of 100 μM SNP significantly alleviated drought-induced increase of ion leakage in the leaves of rice. All of these results demonstrate that applying or scavenging NO hardly affect water content or ion leakage in the leaves of well-watered rice, while exogenous application of NO alleviates water loss and ion leakage in the leaves of drought-stressed rice.

Effects of foliar spray of 100 μM SNP or/and cPTIO on relative water content, water potential and relative ion leakage in the leaves of well-watered and drought-stressed rice. a Relative water content in the leaves of well-watered rice. b Relative water content in the leaves of drought-treated rice. c Water potential in the leaves of well-watered rice. d Water potential in the leaves of drought-treated rice. e Relative ion leakage in the leaves of well-watered rice. f Relative ion leakage in the leaves of drought-treated rice. The assays were performed after 12 days drought treatment. The value is the average of 8 leaves each, vertical bar represents SE of each treatment. Asterisk means significant difference compared with control at the same day
Effect of NO on transpiration rate of rice leaves
A possible effect of NO on transpiration rate was assayed in well-watered and drought-stressed rice (Fig. 7). Exogenous application of NO donor, SNP, significantly inhibited transpiration rate in the well-watered rice, but NO scavenger cPTIO hardly affected transpiration rate in the well-watered rice. In the drought-stressed rice, transpiration rate decreased obviously as the duration, foliar application of SNP aggravated drought-induced decrease of transpiration rate insignificantly. In contrast, foliar application of cPTIO also hardly affected transpiration rate in the drought-stressed rice.

Effects of foliar spray of 100 μM SNP or/and cPTIO on transpiration rate of rice. a well-watered rice. b Drought-treated rice. The value is the average of 8 leaves each, vertical bar represents SE of each treatment. Asterisk means significant difference compared with control at the same day
Discussion
Until now, the effect of drought stress on endogenous NO content in rice leaves remains unclear. In this study, we provided the first evidence that drought stress significantly increased endogenous NO content in rice leaves, with time-dependent changes (Figs. 2, 3). Simultaneously, we observed that drought stress induced a rapid proline accumulation in rice leaves (Fig. 4), and the facts were identical to previous reports on many other plants. Recently, Zhang et al. (2008) reported that copper-induced proline synthesis was associated with NO generation in C. reinhardtii. Based on these results, one might speculate that NO works as a significant signaling molecule in drought-induced proline accumulation in rice leaves. In order to prove this hypothesis, another two evidences are required. First, exogenous application of NO could induce proline accumulation in the leaves of well-watered rice. Secondly, scavenging of NO could inhibit proline accumulation in the leaves of drought-stressed rice. However, in the present study, foliar spray of SNP failed to induce proline accumulation in the leaves of well-watered rice and foliar spray of cPTIO failed to inhibit drought-induced proline accumulation too.
According to the facts that treatment with SNP promoted proline accumulation and treatment with cPTIO inhibited SNP-induced proline accumulation in Cu-treated C. reinhardtii, Zhang et al. (2008) concluded that NO was probably playing important roles in regulating the algae proline accumulation and tolerance to Cu toxicity. However, similar to our observations, exogenous application of SNP hardly affected proline content in C. reinhardtii in absence of Cu (Zhang et al. 2008). These results excluded the possibility that NO works as signaling molecule downstream of drought in drought-induced proline accumulation in rice leaves. Δ1-pyrroline-5-carboxylate synthetase (P5CS) is the rate-limiting key enzyme in the synthesis rout of proline, and its expression and enzymatic activity are controlled by proline levels as well as by environmental stimuli such as salinity, drought, chilling and darkness (Alia and Matysik 2001; Ábrahám et al. 2003). Zhang et al. (2008) reported that treatment with the SNP induced an additional increase in the activity of P5CS and up regulated expression of P5CS in Cu-treated algae, while exogenous application of NO hardly affected the activity of P5CS or expression of P5CS in the algae in the absence of Cu. These results also demonstrate that NO is not directly related with proline content or P5CS activity, the expression of P5CS is not directly regulated by NO.
We demonstrate that treatment with SNP fails to induce proline accumulation in rice and Zhang et al. (2008) demonstrate that treatment with SNP has no effect on proline content in algae without Cu stress. We suppose an explanation might attribute to the observation that treatment with SNP promoted proline accumulation in Cu-treated algae in the study of Zhang et al. (2008). It is reported that proline accumulation in Cu-treated algae exhibits a concentration-dependent change with Cu (Zhang et al. 2008), and it is possible that treatment with SNP increased proline accumulation in Cu-treated algae by enhancing Cu absorption. In higher plant rice, we have demonstrated that exogenous application of SNP in nutrient solution increase Cd accumulation in rice roots (Xiong et al. 2009). Recently, Besson-Bard et al. (2009) demonstrate that NO promotes Cd accumulation in the roots of Arabidopsis. Therefore, we suppose that exogenous application of NO induced an additional increase of proline in Cu-treated algae by increasing Cu accumulation. Of course, this hypothesis requires further verification.
In our study, altered concentrations of exogenous SNP were independent from proline accumulation at both well-watered and drought-stressed conditions in rice leaves. However, this was contrary to the published reports in wheat (Tan et al. 2008) and in fine grain aromatic rice (Farooq et al. 2009). We suggest that these opposite findings on SNP-induced proline accumulation could be explained by the different plant species (or varieties) used, different drought level, different age of the plants and different time of drought treatment. In addition, in the paper of Tan et al. (2008), Proline content was determined and expressed as μmol g−1 fresh mass. As we know, PEG (or drought) treatment causes serious loss of water content in plant leaves, so it is more credible to determine and express proline content with dry weight.
Many evidences demonstrate that drought-induced proline accumulation is regulated by ABA-dependent signaling pathway (Hare et al. 1999), osmotic stress induces ABA accumulation and then regulates the expression of P5CS gene (Xiong et al. 2001). In Arabidopsis, ABA induces NO synthesis in guard cells, with a concomitant response of stomatal closure (Desikan et al. 2002). Our present study demonstrates that NO is not directly related with proline accumulation in drought-stressed rice. Therefore, we also exclude the possibility that NO plays a role downstream of ABA in drought-induced proline accumulation in rice leaves.
Although exogenous application of NO fails to induce proline accumulation in well-watered or drought-stressed rice (Fig. 4), it does alleviate drought-induced water loss and ion leakage in rice leaves (Fig. 6), and these findings are consistent with previous studies (García-Mata and Lamattina 2001; Farooq et al. 2009). Foliar spray of SNP decreased transpiration rate in well-watered rice in present study (Fig. 7), and the result is consistent with the study of García-Mata and Lamattina (2001).
In conclusion, although our studies demonstrate that drought stress induces a simultaneous accumulation of NO and proline, NO is dispensable for drought-induced proline accumulation in rice leaves. Foliar spray of SNP increases endogenous NO content, induces stomatal closure, decreases transpiration rate and enhances the adaptive responses against drought stress in rice.
Abbreviations
- ABA:
-
Abscisic acid
- CEC:
-
Cation exchange capacity
- cPTIO:
-
2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide
- DAF-FM DA:
-
4-Amino-5-methylamino-2′,7′-difluorofluorescein diacetate
- DW:
-
Dry weight
- FW:
-
Fresh weight
- NO:
-
Nitric oxide
- P5CS:
-
Δ1-Pyrroline-5-carboxylate synthetase
- PPED:
-
Photosynthetic photo flux density
- SNP:
-
Sodium nitroprusside
- SW:
-
Saturated weight
- ROS:
-
Reactive oxygen species
- RWC:
-
Relative water content
References
Ábrahám E, Rigó G, Székely G, Nagy R, Koncz C, Szabados L (2003) Light-dependent induction of proline biosynthesis by abscisic acid and salt stress is inhibited by brassinosteroid in Arabidopsis. Plant Mol Biol 51:363–372
Alia MP, Matysik J (2001) Effect of proline on the production of singlet oxygen. Amino Acids 21:191–203
Arasimowicz-Jelonek M, Floryszak-Wieczorek J, Kubiś J (2009) Involvement of nitric oxide in water stress-induced responses of cucumber roots. Plant Sci 177:682–690
Bates LS, Waldren RP, Tear ID (1973) Rapid determination of free proline for water stress studies. Plant Soil 39:205–207
Besson-Bard A, Gravot A, Richaud P, Auroy P, Duc C, Gaymard F, Taconnat L, Renou J, Pugin A, Wendehenne D (2009) Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake. Plant Physiol 149:1302–1315
Crawford NM, Guo FQ (2005) New insights into nitric oxide metabolism and regulatory functions. Trends Plant Sci 10:195–200
Desikan R, Griffiths R, Hancock J, Neill S (2002) A new role for an old enzyme: nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. PNAS 99:16314–16318
Farooq M, Basra SMA, Wahid A, Rehman H (2009) Exogenously applied nitric oxide enhances the drought tolerance in fine grain aromatic rice (Oryza sativa L.). J Agron Crop Sci 195:254–261
Fu J, Huang B (2001) Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ Exp Bot 45:105–114
García-Mata C, Lamattina L (2001) Nitric oxide induces stomatal closure and enhances the adaptive plant responses against drought stress. Plant Physiol 126:1196–1204
Hao G, Xing Y, Zhang J (2008) Role of nitric oxide dependence on nitric oxide synthase-like activity in the water stress signaling of maize seedling. J Integr Plant Biol 50:435–442
Hare PD, Cress WA, van Staden J (1999) Proline synthesis and degradation: a model for elucidating stress-related signal transduction. J Exp Bot 50:413–434
Lamattina L, García-Mata C, Graziano M, Pagnussat G (2003) Nitrate oxide: the versatility of an extensive signal molecule. Ann Rev Plant Biol 54:109–136
Lei Y, Yin C, Ren J, Li C (2007) Effect of osmotic stress and sodium nitroprusside pretreatment on proline metabolism of wheat seedlings. Biol Plant 51:386–390
Loggini B, Scartazza A, Brugnoli E, Navari-Izzo F (1999) Antioxidant defense system, pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physio 119:1091–1099
López-Carrión AI, Castellano R, Rosales MA, Ruiz JM, Romero L (2008) Role of nitric oxide under saline stress: implications on proline metabolism. Biol Plant 52:587–591
Luna CM, Pastori GM, Driscoll S, Groten K, Bernard S, Foyer CH (2005) Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J Exp Bot 56:417–423
Ma XL, Wei XH, Long RJ, Cui WJ, Wan YL (2005) Studies on mechanism of enhancing the chilling resistance of annual ryegrass by exogenous nitric oxide. Acta Ecol Sin 25:1269–1274
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell Environ 25:239–250
Qiao W, Fan L (2008) Nitric oxide signaling in plant responses to abiotic stresses. J Integr Plant Biol 50:1238–1246
Shinozaki K, Yamaguchi-Shinozaki K, Seki M (2003) Regulatory network of gene expression in the drought and cold stress responses. Curr Opin Plant Biol 6:410–417
Siripornadulsil S, Traina S, Verma DPS, Sayre RT (2002) Molecular mechanisms of proline-mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell 14:2837–2847
Tan J, Zhao H, Hong J, Han Y, Li H, Zhao W (2008) Effects of exogenous nitric oxide on photosynthesis, antioxidant capacity and proline accumulation in wheat seedlings subjected to osmotic stress. World J Agri Sci 4:307–313
Taylor GB (1996) Proline and water deficit. Ups, downs, ins and outs. Plant Cell 8:1221–1224
Tian X, Lei Y (2006) Nitric oxide treatment alleviates drought stress in wheat seedlings. Biol Plant 50:775–778
Xiong L, Ishitani M, Lee H, Zhu JK (2001) The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cofactor sulfurase and modulates cold stress and osmotic stress responsive gene expression. Plant Cell 13:2063–2083
Xiong J, An L, Lu H, Zhu C (2009) Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta 230:755–765
Xiong J, Fu G, Tao L, Zhu C (2010) Roles of nitric oxide in alleviating heavy metal toxicity in plants. Arch Biochem Biophys 497:13–20
Zhang J, Kirkham MB (1994) Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol 35:785–791
Zhang LP, Mehta SK, Liu ZP, Yang ZM (2008) Copper-induced proline synthesis is associated with nitric oxide generation in Chlamydomonas reinhardtii. Plant Cell Physiol 49:411–419
Zhao MG, Chen L, Zhang LL, Zhang WH (2009) Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol 151:755–767
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (30623006 and 30871473), China National Key Technology R&D Program in the 11th Five year Plan (NO. 2009BAK43B12), Specific Foundation of Commonweal Program (Agriculture) (NYHYZX07-001), Zhejiang Provincial Natural Science Foundation of China (Y3100090, Y3100060, Y3110040), Zhejiang Key Program Foundation (2008C22073, 2009C32048) and Sannong Wufang Program of Zhejiang Province (SN200806).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Xiong, J., Zhang, L., Fu, G. et al. Drought-induced proline accumulation is uninvolved with increased nitric oxide, which alleviates drought stress by decreasing transpiration in rice. J Plant Res 125, 155–164 (2012). https://doi.org/10.1007/s10265-011-0417-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-011-0417-y