
Abstract
The Markermeer is a large and shallow man-made freshwater lake in the Netherlands, characterized by its high turbidity. As part of a study aiming to mitigate this high turbidity, we studied the water–bed exchange processes of the lake’s muddy bed. The upper centimeter’s–decimeter’s of the lake bed sediments mainly consists of soft anoxic mud. Recent measurements have proved the existence of a thin oxic layer on top of this soft anoxic mud. This oxic layer, which is much easier to be eroded than the anoxic mud, is believed to be related with Markermeer’s high-turbidity levels. Our hypothesis is that the thin oxic layer develops from the anoxic mud, enhanced by bioturbation. Actually, we will demonstrate that it is the bioturbated state of the bed that increases its erodability, and not the oxidation state of the sediments. In particular, we will refer to bioturbation caused by meiobenthic fauna. The objective of this study is therefore to determine the influence of the development of the thin oxic layer on the water–bed exchange processes, as well as to establish the role of bioturbation on those processes. This is done by quantifying the erosion rate as a function of bed shear stresses, and at different stages of the development of the oxic layer. Our experiments show that bioturbation increases the rate at which Markermeer sediments are eroded by almost an order of magnitude. The short-term fine sediment dynamics in Markermeer are found to be driven by the complex and highly dynamic interactions between physics, chemistry, and biology. Finally, the long-term fine sediment dynamics are driven by the erosion of the historical deposits in the lake’s bed, which is only possible after bioturbation, and which leads to an increase of the stock of sediments in the lake’s muddy bed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The Markermeer is a large man-made freshwater lake located in the center of the Netherlands. Together with the northern IJsselmeer it is one of the largest natural freshwater reservoirs of Europe. This area is known as the IJsselmeer Region. The average suspended sediment concentration in the Markermeer is about 50 mg/l near the surface. During storms, the sediment concentration increases to a level of about 100 mg/l or more (Van Kessel et al. 2008). It is assumed that the turbidity in the Markermeer started to increase after the lake’s closure in 1975. However, there is no clear increasing trend in turbidity between 1982 and today (Vijverberg et al. 2011). During the last decades, the lake has experienced a decrease in ecological values (Noordhuis and Houwing 2003; van Eerden and van Rijn 2003). The reasons for this ecological deterioration are still uncertain. However, fine sediments are considered to be an important stress factor for the ecosystem of the lake. Furthermore, water quality problems are also related to sediment composition and transport in the Markermeer (Van Duin 1992). Therefore, fine sediments in the system seem to be a key factor in explaining the negative trends in the ecosystem over the last decades. As part of a study that aims to mitigate Markermeer’s high turbidity, we studied the water–bed exchange processes of the lake’s muddy bed.
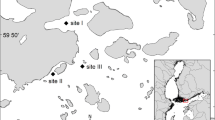
Lake Markermeer did not exist before the twentieth century. The IJsselmeer Region was then the Zuiderzee, a shallow inlet from the North Sea of about 5,000 km2. During the Zuiderzee era, when the whole region was still a large estuary, there was a net landward fine sediment flux, caused by tide and estuarine circulation. Thick layers of clay and loam were deposited as a result of this flux, and given certain favorable local conditions (e.g., sheltered areas). Then, in the twentieth century, the Zuiderzee works took place, and the morphology of the region changed entirely. Figure 1 shows the current configuration of the IJsselmeer Region. Markermeer was created at the head of the old sea inlet, with finer bottom sediments and smaller depths than the northern IJsselmeer. The closure and the disconnection from the sea led to changes in the hydrodynamics of the system. The dike separating the lakes, known as the Houbtrijdijk, does not allow fine sediments to be transported out of Markermeer.

The IJsselmeer region. The dikes are indicated in red. The dike separating the IJsselmeer Region and the North Sea was finished in 1932. The dike separating the Markermeer and the IJsselmeer was finished in 1975
The Markermeer is a shallow lake, with a mean water depth of 3.6 m. About 90 % of the lake has a water depth between 2 and 5 m (Vijverberg 2008). The total surface of water, including Lake IJmeer, is 691 km2 (Coops et al. 2007). The volume of stored water is about 2.5 109 m3 (Van Duin 1992). The residence time ranges between 6 and 18 months (Vijverberg 2011). The large-scale flow pattern in Markermeer is mainly dominated by wind, inducing horizontal circulations (Vijverberg 2011). These circulations may have opposite directions in the surface and near the bottom, which results in a complex 3D flow pattern (Vijberverg 2011). Different large-scale circulation patterns can occur depending on wind direction (van Kessel et al. 2008). This water circulation is responsible for the fine sediment dispersion over the system. Wind also generates waves, which, together with currents, induce bed shear stresses that may resuspend sediments from the bed of the lake. The relationship between wind speed and suspended solids in the Markermeer is shown in Fig. 2. Wind-induced waves may have a more important effect on re-suspension of sediments from the bed than wind-induced currents in the Markermeer (Royal Haskoning and Delft Hydraulics 2006).

Relationship between wind speed and suspended solids in Markermeer for temperatures above and below 15 °C (Noordhuis 2010). Note that temperature is most likely a proxy for winter conditions—the larger values in wintertime are likely induced by memory effects of the large Markermeer sedimentary system
Figure 3 shows a schematic illustration of the sediment layers in the bed of the Markermeer. The upper sediment layer in the bed is a very thin oxic layer. It is well known that thin oxic layers on a mud surface exert a pronounced influence on the exchange of substances across the mud–water interface (Mortimer 1942). In fact, erosion and deposition of the upper oxic mud layer dominates the suspended sediment concentration in the water column (Vijverberg 2011). Thus, it is crucial to quantify the erodability of this oxic layer, as well as to establish the mechanisms behind it. A thicker soft anoxic mud layer is present underneath the oxic layer. This anoxic mud is mobilized during storm events only, while erosion of the oxic layer can occur already at low wind speeds (e.g., Bft 3) (Vijverberg 2011). When the oxic layer is completely eroded from the bed, a new, easily erodible oxic layer will develop from the more stable anoxic layer. Our main hypothesis is that the mechanism behind the development of the new oxic layer and consequent increase in erodability is bioturbation. Diffusion of oxygen into the anoxic layer is of course required for the oxidation of the sediments, inducing the yellowish color. However, the increased erodability of the oxic layer cannot be explained by diffusion of oxygen into the sediments. Thus, we hypothesize that bioturbation further enhances the development of the oxic layer, as well as increases its erodability. The aim of the study described in this paper is to test this hypothesis.

Schematic illustration of sediment layers in the bed of Markermeer. The main processes affecting the dynamics of these layers are also sketched in this Figure
Bioturbation includes the processes of feeding, burrowing, and locomotory activities of sediment-dwelling benthos (Fisher and Lick 1980), and severely affects sediment dynamics (Le Hir et al. 2007). Bioturbators form an important subgroup within benthic biota (Knaapen et al. 2003), which can attain high densities in soft mud substrata (Pearson and Rosenberg 1978). Previous researchers have measured the effect of bioturbation in the erodibility of fine sediments (Rhoads 1970; Willows et al. 1998; Widdows et al. 1998 and 2000; Amaro et al. 2007). Most of them used an annular flume and characterized this erodability through the increase in turbidity levels in the water column. Others have studied the feeding mode and feeding rates of bioturbating fauna, and have characterized their effect on the stratigraphy of the sediment deposits (Davis 1974; Fisher and Lick 1980; van de Bund 1994). We have measured the effect of bioturbation by meiobenthic fauna on the erodability of sediments, but our approach focuses on quantifying the erosion rate as a function of bed shear stresses. These erosion rates were measured at several times within the development of the thin oxic layer.
In this paper, we present the results of three groups of experiments: groups I, II, and III. Each group of experiments aims to address a particular research question. In group I, we compare the erodibility of the oxic layer in a defaunated sample with the erodibility of the oxic layer in a bioturbated sample. Note that an oxic layer is expected to develop in the defaunated sample due to diffusion of oxygen, while development of the oxic layer in the bioturbated sample is expected to be enhanced by the increase in porosity by bioturbation. In group I, bioturbation was caused by all meiobenthic bioturbators found in a fresh sample of Markermeer sediments. The research questions to be answered with group I are the following: (1) Is bioturbation indeed an important driver of the physico-chemical water–bed exchange processes in the Markermeer, (2) Does oxidation of the sediments increase their erodability or is bioturbation responsible for the increase in erodability of the bed, and (3) Does the increase in erodability by bioturbation (partly) explain the high turbidity of the Markermeer. In groups II and III, we study the bioturbation effect of Tubifex only. Our sampling revealed that Tubifex is an important meiobenthic bioturbator in the Markermeer. At some locations, it was not the most abundant, but it was present at all the locations we analyzed. In group II, we studied the effect of bioturbation by Tubifex in two soft anoxic mud samples: a surficial sample and a 10-cm deep sample. The research questions to be answered with group II are as follows: (1) What is the specific effect of Tubifex in the bioturbation process and (2) Has the degree of consolidation of the bed cause an impact in the effect of bioturbation. Furthermore, group II aims to provide additional information to contribute in answering research question (3) from group I. Finally, in group III, we study the effect of bioturbation by Tubifex in the Zuiderzee deposits. The research question to be answered with group III is (1) If bioturbation of the Zuiderzee deposits is possible, and, if so, what are the consequences of this for the long-term sediment transport processes. The last question will help in understanding the long-term sediment transport processes is the lake, which are currently unknown.
2 Methods
Cylindrical bed cores were collected from the Markermeer. The sampler was designed and constructed at Delft University of Technology. The diameter of the cylinders is 0.1 m. Most of the cores collected contained a thin oxic layer on top of the soft anoxic mud, and with some Zuiderzee deposits at the bottom of the core. The experimental treatment used in our experiments is illustrated in the sequences shown in Fig. 4. The upper sequence in Fig. 4 illustrate the treatment applied for all bioturbation experiments: firstly, we removed the thin oxic layer from the surface of the soft anoxic mud. As a result, the surface of the soft anoxic mud became smooth and unbioturbated. The soft anoxic mud was then carefully extruded from the sampling cylinder and placed in another cylinder. The latter was designed to fit into the erosion device, with 0.1-m diameter as well. The approximate height of the extruded soft anoxic mud was 0.1 m. Secondly, an initial erosion experiment was performed to the anoxic mud sample, to characterize the erodibility of the not yet bioturbated sample. Thirdly, individuals of Markermeer meiobenthic bioturbators were added to the sample, aiming to enhance the development of an oxic layer. In group I, we added all the meiobenthic bioturbators we found in a fresh sample. In groups II and III, we added Tubifex only. All the meiofauna was obtained by sieving fresh and oxic mud through 250- and 500-μm sieves. The cylinders were kept in a small chamber, in which temperature and light conditions were controlled to mimic field conditions. We chose 6 °C and no light exposure. Incubated for n days, the upper part of the sample oxidized and became bioturbated, forming a thin oxic layer on top of the soft anoxic mud, upon which, fourthly, an erosion experiment was executed. The erodibility of an n-days-old thin oxic layer was thus characterized. After an erosion experiment was finished, the thin oxic layer was removed, the meiofauna in the thin oxic layer sieved and stored, and the process was repeated from the first step. We performed erosion experiments at n = 2, 4, 6, and 8 days after the beginning of the thin oxic layer development and bioturbation (the n of each step is therefore 2 days higher than for the preceding step). We will refer to these experiments as days 2, 4, 6, and 8, respectively. The lower sequence in Fig. 4 illustrate the treatment applied to the defaunated sample in group I. In this case, the sample was first defaunated by sterilization with gamma rays at the Nuclear Reactor Institute Delft. This gamma ray treatment ensured the absence of fauna in the sample. Bioturbators were never added to this defaunated sample. The experiments in group I were executed in the following order: days 4, 2, 8 and 6. In groups II and III, the sequence was as follows: days 2, 4, 6, and 8. Table 1 presents a summary of all the experiments performed for all groups, specifying name, bioturbator type, and bed material type for each of the experiments. Note that for Group IIb, we used a soft anoxic mud sample obtained by removing the upper 0.1 m of the core, aiming to test bioturbation in a more consolidated bed.

Upper sequence experimental treatment used in all samples where we studied the effect of bioturbation. Lower sequence experimental treatment used in the defaunated sample
To the bioturbated sample in group I, we added a specified number of individuals of meiofauna to the first erosion experiments. The survivors of the first experiment were added to the second one, etc. In groups II and III, 11 individuals of Tubifex were added at the beginning of every experiment.
The erosion experiments were performed with an UMCES-Gust Erosion Microcosm System (Gust 1989 and 1990). This microcosm was calibrated at Deltares, using sand of various diameters and Shield/van Rijn’s erosion diagram (van Rijn 1984). The resulting calibration curve can be seen in Fig. 5. The following nine bed shear stresses were applied at every erosion experiment on Markermeer samples: 0.05, 0.1, 0.2, 0.3, 0.4, 0.5, 0.6, 0.7, and 0.8 Pa. Every bed shear stress lasted for 60 s. A so-called OSLIM turbidity meter was installed at the suction outlet of the microcosm, with which the turbidity of the out-flowing water was measured. The turbidity meter was calibrated with samples at different concentrations of the sediments to be studied. Eq. 1 shows how the eroded mass was calculated from the turbidity data:

Calibration curve of the UMCES-Gust Erosion Microcosm System. The error bars were calculated by using D10 and D90 (instead of D50) for the calculation of the critical shear stress according to Shields/van Rijn (van Rijn 1984)
where E is the eroded mass (in grams), c is the sediment concentration (in grams per liter) in the out-flowing water, Q is the discharge (in liters per second) of water through the suction outlet, and Δt (in seconds) is the interval between two measurements of the turbidity meter. The erosion rate (in grams per square meter per second) was then calculated by dividing the eroded mass by the surface of the microcosm section, and by the number of seconds within each bed shear step. Thus, we obtained one erosion rate per applied bed shear stress.
3 Results and analysis
3.1 Group I experiments
In this group of experiments, we studied the difference in erodibility between a defaunated sample and a bioturbated sample. The type of sediments studied was soft anoxic mud in both cases. Several meiobenthic bioturbators fed and burrowed in the bioturbated sample. These species were collected from a fresh sample, and consisted of Hydrobia ulvae, Chironomid larvae, Nematoda, Ostracoda, and Tubifex. The most abundant were Ostracoda and Tubifex. Figure 6 shows the density of each of the species present in the fresh oxic layer, as well as those that were found after every erosion experiment. The density of animals after experiment day 6 was the lowest. The oxic layer thickness in the bioturbated sample was 1 mm after 2 days, 1 mm after 4 days, 2 mm after 6 days, and 2.5 mm after 8 days. The thickness of the oxic layer in the defaunated sample was 1 mm, remaining constant thereafter. The oxic layer in the defaunated sample was formed by diffusion of oxygen only. Figure 7a shows the erosion rate as a function of the bed shear stress for the bioturbated sample. The rate at which a 2-day-old oxic layer erodes is higher than the erosion rate of the anoxic mud from which the oxic material developed. This holds for six out of the nine bed shear stresses applied. Please note that the erosion rate under 0.8 Pa is smaller than the erosion rate under 0.5 Pa. We elaborate on this inconsistency in the last chapter of this paper. All erosion rates of the oxic layers are higher than the erosion rate of the anoxic mud from which they developed. The erosion rate of a 4-day-old oxic layer is higher than the erosion rate of a 2-day-old oxic layer for four out of the nine bed shear stresses, while these rates are almost similar for the other five. However, the erosion rate of a 6-day-old oxic layer is higher than the erosion rate of a 4-day-old oxic layer only for the 0.8 Pa test. This is probably caused by a high mortality of meiofauna during the day 6 experiment. Finally, the erosion rate of an 8-day-old oxic layer is higher than the erosion rate of a 4-day-old oxic layer for the highest three bed shear stresses, while similar for the other bed shear stresses.

Density of each specie present in the field oxic layer, as well as after each of the erosion experiments within group I

Comparison between erodibility of a bioturbated sample and erodibility of a defaunated sample (group I). The left panel shows the erodibility of a bioturbated sample as function of bed shear stress, and as a function of the time since the bioturbation process started. The right panel shows the erodibility of a defaunated sample as a function of bed shear stress, and as a function of time since the oxidation process started
Figure 7b shows the erosion rate as a function of the bed shear stress for the defaunated sample. An oxic layer did develop on the defaunated sample as well, but did not exceed 1-mm thickness. The erosion rate was always between 0 and 0.1 (in grams per square meter per second) at each bed shear stress, and did not show any evolution over time. Furthermore, the erosion rate of the anoxic layer was higher than the erosion rate of the oxic layer for all the defaunated experiments. Thus, oxidation of the anoxic layer by diffusion of oxygen does not increase the erodibility of the bed in the range of bed shear stresses we studied, but decreases it.
Figure 8a shows the cumulative eroded mass for all the experiments on bioturbated samples, as well as for all the anoxic samples from which they developed. The total eroded mass (TEM) from a 2-day-old oxic layer is higher than the TEM from the anoxic layer from which the oxic layer developed. Actually, the TEM from any of the oxic layers is higher than the TEM from the anoxic layer from which they developed. The TEM from a 4-day-old oxic layer is higher than the TEM from a 2-day-old oxic layer. However, the TEM from a 6-day-old oxic layer is smaller than the TEM from a 4-day-old oxic layer, but still higher than the TEM from a 2-day-old oxic layer. Finally, the TEM from an 8-day-old oxic layer is the highest amongst the observations. Figure 8b shows the cumulative eroded mass for all the experiments in the defaunated sample. All TEM from oxic layers in Fig. 8b are smaller than the TEM from the anoxic layers. The critical shear stress for erosion can be defined as the stress at which the erosion curves in Fig. 8a deviates from zero. According to that criterion, there is not a clear effect of bioturbation on the critical shear stress for erosion. Figure 9 gives an overview of the lower range of erosion rates. Erosion rates in the range 10−3 to 10−2 g/m2s occur at 0.05 Pa.

Cumulative eroded mass (group I). The left panel shows the cumulative eroded mass of the bioturbated sample. The right panel shows the cumulative eroded mass of the defaunated sample

Erodibility of a bioturbated sample as function of bed shear stress
3.2 Group II experiments
In this group of experiments, we studied the individual effect of Tubifex on the erodibility of Markermeer sediments. This group is composed of two sub-experiments. For group IIa, we used a surficial soft anoxic mud sample, while for group IIb, we used a soft anoxic mud sample obtained by removing the upper 0.1 m of the core. Both samples were extruded from the same sediment core. The oxic layer thickness in group IIa was 1.2 mm after 2 days, 1.5 mm after 4 days, 2 mm after 6 days, and 2.5 mm after 8 days. The oxic layer thickness in group IIb was 1 mm after 2 days, 1 mm after 4 days, 1.5 mm after 6 days, and 2 mm after 8 days. Figure 10 shows the erosion as a function of bed shear stress in group II, as well as the cumulative eroded mass for all the experiments in group II. Figure 10a, b refers to group IIa, while Fig. 10c, d refers to group IIb. Figure 10a, c shows that erosion rates increase with increasing bioturbation time. However, there are some deviations from that trend: the 8-day-old oxic layer in Fig. 10a and the 6-day-old oxic layer in Fig. 10c. The erosion rates of these layers deviate from the general pattern. For group I, the highest erosion rates were found at the highest bed shear stresses. This is not the case for group II. Most of the highest erosion rates in Fig. 10a were found at bed shear stresses of 0.3 and 0.4 Pa. Thus, the bed shear stresses producing the highest erosion rates in group I are not producing the highest erosion rates in group II. We believe that this discrepancy between groups I and II is related to the way through which different species of meiofauna affects the bed characteristics: the effect of Tubifex only is probably different from the effect of the group of species in group I.

Erosion rate as a function of bed shear stress, and cumulative eroded mass (group II). a Erodibility of the surficial anoxic sample as function of bed shear stress and as a function of the time since the bioturbation process started. b Cumulative eroded mass as a function of experiment time and as a function of the time since the bioturbation process started. c Erodibility of the 0.1-m deep anoxic sample as a function of bed shear stress and as a function of time since the bioturbation process started. d Cumulative eroded mass as a function of experiment time and as a function of the time since the bioturbation process started
Figure 10b, d show the cumulative eroded mass in group II. Again, the trend is clear: the longer the bioturbation time, the higher the TEM. The 8-day-old oxic layer in Fig. 10b and 6-day-old oxic layer in Fig. 10d are exceptions.
3.3 Group III experiments
In Group III, we studied the effect of Tubifex on the erodibility of the Zuiderzee deposits. We presume that bioturbation of the Zuiderzee deposits is only possible when oxygen is available at its surface, and therefore when this surface is not covered by anoxic mud. The oxic layer thickness in the sample was 1 mm after 2 days, 1.5 mm after 4 days, 1.5 mm after 6 days, and 2 mm after 8 days. Figure 11a shows the erosion rate as a function of bed shear stress in group III, while Fig. 11b shows cumulative eroded mass for all the experiments in group III. The erosion rate increases as a function of bioturbation time at most of the bed shear stresses. The highest increase in erosion rate over time can be observed at 0.3 Pa. Please note that the vertical scale in Fig. 11a is twice the vertical scale of Figs. 7 and 10a, c. The vertical scale in Fig. 11b is also three times larger than the vertical scale in Figs. 8 and 10b, d. Figure 11b shows that the TEM increases as a function of the bioturbation time as well. The TEM of a non-bioturbated Zuiderzee bed is never higher than 0.02 g. This is the lowest TEM measured among all the experiments presented in this paper. However, after 8 days of bioturbation, the TEM on a Zuiderzee bed is in the range of the TEM of a non-consolidated anoxic bed. The critical shear stress for erosion of a non-bioturbated Zuiderzee bed cannot be clearly identified in the range between 0 and 0.8 Pa, while all the bioturbated Zuiderzee beds showed a critical shear stress for erosion of 0.2 to 0.3 Pa.

Erosion rate as a function of bed shear stress and cumulative eroded mass for group III
4 Discussion
In the defaunated sample, unstirred by bioturbation, a thin oxic layer develops within a day, due to the diffusion of oxygen into the sediments. The thickness of this oxic layer is approximately 1 mm, and remains constant over time because of oxygen consumption within the bed. With bioturbation, the bed is disrupted by channels, burrows, and pelletization, increasing the bed’s water content. Oxygen can therefore penetrate further, and thus, the oxic layer thickness increases over time, well beyond the thickness of the oxic layer in the defaunated sample. The strength of the thin oxic layer in the defaunated sample is slightly higher than the strength of the anoxic mud from which the thin oxic layer developed. The observed small increase of bed strength upon oxidation of sediments in the defaunated sample can be explained by a cementation effect caused by the formation of Fe+++, which may form rust (Fe2O3), bog-ore (Fe(OH)3), and/or various forms of FeOOH (Dniker et al. 2003; Ivanov and Chu 2008). With bioturbation, erodibility increases as a function of bioturbation time, due to disturbance of the bed, but not because of changes in chemical sediment properties. Actually, in the case of bioturbation, there should also be iron oxide formation, since bioturbation also causes oxidation. However, the bioturbation disturb the sediment network continuously in a shorter time scale, so that reinforcement by iron oxides is not possible. Therefore, it has been demonstrated that bioturbation drives the physico-chemical water–bed exchange processes, increasing erodability of the bed and oxic layer thickness as a function of bioturbation time. Oxidation of the sediments in the bed itself does not increase the erodability of the bed under the range of bed shear stresses we studied, and, therefore, bioturbation must be the only driver of the increased erodability of the bed. Furthermore, there are also important consequences to the system to be highlighted: without biota effects, significant erosion of the bed may only occur at bed shear stresses much higher than 0.8 Pa. How much higher cannot be deduced from the experiments presented in this paper. Wind speeds ranging from 10 to 13 m/s (depending on wind direction) would be needed to reach a 0.8-Pa bed shear stress in the middle of the lake (van Kessel et al. 2008). Wind speeds exceeding 10 m/s were recorded 11 % of the time during 2010 in the middle of the Markermeer. In that way, we would have erosion of the bed only 11 % of the time, whereas the lake is actually characterized by a highly turbid state. Therefore, bioturbation is an important mechanism behind the high turbidity in Markermeer.
The erodibility of the group IIa-bioturbated oxic layers is higher than the erodibility of the group IIb-bioturbated oxic layers. Furthermore, the erodibility of the group IIa anoxic layers is higher than the erodibility of the group IIb anoxic layers. This is attributed to the higher degree of consolidation of the sample in group IIb. However, the total eroded mass from the 8-day-old oxic layer in group IIb, is higher than the total eroded mass from a 4-day-old oxic layer in group IIa. Thus, enough bioturbation time can increase the erodibility of a bed up to the erodibility characteristic of a less consolidated bed. We have observed two ways in which bioturbation disturbs the bed. These are related with the type of meiobenthic fauna present in the bed. Montserrat (2008) uses a classification of benthic fauna as a function of their impact on the benthic region. He distinguishes tube-building fauna, surface-disrupting species, surface pelletizers and biodiffusing bivalves. The first type of disturbance observed in our experiments is caused by surface disrupters and biodiffusing bivalves. Ostracoda is a biodiffusing bivalve, and H. ulvae is a surface disrupter. These species move around the sediment-water interface, increasing the water content of the bed and therefore decreasing its strength. This results in weakening of the bed. The second type of disturbance observed is caused by surface pelletizers. According to the descriptions by Fisher et al. (1980) and Davis (1974), Tubifex can be classified as a surface pelletizer. This is supported by our visual observations of the bioturbated samples in groups II and III experiments where fecal pellets were found on the surface of the sediments, while tubes or other types of disruptions were not. Figure 12 shows a picture of the bed before erosion experiment day 6 in group IIb, from which we can observe several large clusters of pellets deposited on top of the mother bed. Some clusters of pellets can be seen near the curved wall of the microcosm in Fig. 12. Tubifex ingest sediment from the substratum and expel this material at the sediment-water interface (Fisher et al. 1980). This result in the mixing and subduction of sediments: a layer of sediment above the zone of peak feeding moves downward as a fairly discrete unit (Fisher et al. 1980). Therefore, the surface of a sediment bed in which Tubifex is active may be completely pelletized after a certain time, as described by Rhoads (1970), who analyzed the development of fecal-rich surfaces under bioturbators influence. The pelletization time varies with meiofauna density and temperature. This type of bioturbation has important implications for the erodibility of the bed. The Tubifex worms rework the fine sediments deeper into the bed, forming loose aggregates on the bed surface. These pellets are more easily erodible than the undisturbed bed sediments. Some Tubifex individuals are likely to be more effective than others, producing larger/more pellets than other individuals. Thus, a pelletized bed consists of a distribution of pellet-size classes. Each size class will be eroded at different eroding forces. This is in line with the work of van Prooijen and Winterwerp (2010), who suggested that some parts of cohesive sediment beds are more likely to erode than others, and due to, e.g., difference in sizes and bonding properties.

Pelletized bed before performing erosion day 6 experiment in group IIb. Large clusters of pellets can be observed on the bed, especially close to the curved wall of the microcosm
As discussed above, we have observed that during some experiments, erosion rates decrease with increasing bed shear stress, as shown in Figs. 10a, c; 11a; and 7a. This decrease in erodibility is to be attributed to starvation of erodible sediment. There are two possible causes for such starvation. The first possible explanation is a vertical gradient in strength within the bed, induced by physico-chemical processes such as consolidation. However, this gradient is likely to be destroyed by the burrowing meiofauna. The second explanation is a depletion of sediments available for erosion. These sediments may either originate from a region of the bed weakened by surface disrupters, or may consist of fecal pellets deposited on the surface. In groups II and III, starvation of erodible sediments occurred during all experiments. In group I, such starvation occurred in two out of four erosion experiments. These observations are in agreement with Amos (1992), who defined an erosion type Ia, associated with the erosion of fecal pellets in a highly active benthic environment. This type of erosion was characterized by a steep increase of erosion rate after an increase in bed shear stress, followed by a smooth decrease over the next 3 min. Hence, the pelletization of the bed and subsequent erosion rate are a function of the number of bioturbating individuals and their efficiency, and of the time available to build up the pellets on the bed, prior to removal by erosion. A third factor is the degradation of pellets. Sediment ingested by Tubifex is packaged into a mucus (McCall 1979), which breaks down with time, resulting in degradation of the fecal pellet. McCall (1979) studied the pellet degradation rate, and found that the pellets broke down within 2 to 20 days, depending on temperature and mechanical stirring. Thus, the number of pellets available for erosion is determined by the complex balance between production of pellets and breakdown of the mucus. In our work, we did not study the behavior of degrading pellets, but it is an important factor to be taken into account to predict availability of pellets for erosion. The specific role of Tubifex in the bioturbation process has been defined.
Next to erosion of the bed by pick-up of pellets, we observed erosion of the bioturbated bed during days 6 and 8 in the group I experiment, as shown in Fig. 7a. In that case, erosion rates continued to increase with increasing bed shear stress. From our data, it is difficult to assess the distribution of eroded material originating from pellets or from the bioturbated bed. However, we anticipate this distinction can be made with the use of a simple mathematical model of the bed response to bioturbation, the parameters of which can be calibrated against our observations.
Tubifex individuals were found in 10 out of the 12 samples we analyzed. In fact, they were the dominant species in nine samples, and the only present species in four samples. However, we cannot be certain that Tubifex is the dominant species for the bioturbation in the Markermeer without further data on the biota and their relative effects. Nevertheless, it was found that Tubifex is abundant in organic rich waters because of lack of competition and abundant food supply, in conjunction with a high tolerance for reduced oxygen conditions (Chapman 2001). As all available information and literature suggests that Tubifex should indeed be abundant in the Markermeer, erosion of pellets by the flow is likely an important erosion mechanism in the Markermeer. These observations suggest that the current short-term sediment dynamics are characterized by a continuous cycle of pelletization, surface disruption, erosion, flocculation, deposition, consolidation, reduction, and oxidation, all subject to the wind forcing exerted on the sediment surface through flow and wave action. All together, these phenomena form a highly dynamic system driven by the interaction of physics, chemistry, and biology. A sketch of that system is given in Fig. 13. The fate of sediments in the current state of Markermeer is determined by the complex interactions between all the phenomena in Fig. 13, and long-term predictions on the sediment dynamics should be based on an understanding of all these phenomena and their interactions.

Sketch of the highly dynamic system driven by the interaction of physics, chemistry, and biology. All together, these processes determines Markermeer sediment dynamics
Erodibility of the bed under large bed shear stresses is not addressed in this paper. However, Vijverberg (2011) found that the soft anoxic mud can be mobilized under storm conditions only. This is relevant information for the Markermeer sediment dynamics as well. Tubifex can harvest in anoxic sediments, as long as part of their body remains in contact with oxygen. Hence, also part of the anoxic layer is subject to bioturbation, losing its strength. It is this weaker material that can be mobilized under storm conditions, as observed by Vijverberg et al. (2011).
Our results have further important implications for the fine sediments dynamics in the Markermeer. Currently, it is hypothesized that Markermeer’s soft anoxic mud stems from the Zuiderzee deposits. This hypothesis was already postulated by van Duin (1992), who suggested that the “IJsselmeer deposits” partly originate from sediments from the river IJssel and partly from erosion of the more ancient Zuiderzee deposits. Without bioturbation, the erosion rate of the Zuiderzee deposits varies from 0.001 to 0.01 g/(m2 s), for bed shear stresses of 0.2 to 0.8 Pa. However, when bioturbated, erosion rates reach values characteristic for a soft anoxic bed. In particular, we measured erosion rates up to 0.2 g/(m2s) under 0.3 Pa after 8 days of bioturbation by Tubifex. This is one order of magnitude higher than without bioturbation. In practice, this implies that Zuiderzee deposits become erodible only after bioturbation. However, bioturbation of the Zuiderzee deposits is only possible when their surface is exposed to oxygen, activating the meiofauna in the sedimentary deposits. Therefore, an inference of these findings is that the soft anoxic mud protects the Zuiderzee deposits from bioturbation, hence from further erosion. This explains why the total amount of soft sediment in the Markermeer is still fairly limited, even though the current hydrodynamic conditions prevail already for more than three decades.
Let us now evaluate the possibility that the soft anoxic mud actually stems from bioturbation-associated erosion of the Zuiderzee deposits. Let’s also assume no soft anoxic mud layer present on the bed of the lake before the closure of the Houtribdijk. In that case, bioturbation of the Zuiderzee deposits was possible over the entire Markermeer bed. The bioturbated material would be eroded from the bed, as shown in our experiments, and transported over the lake. Next, these sediments settle in the bed again, in the form of oxic sediments. These sediments accumulate in specific places of the lake, and after achieving a certain thickness, reduction of the sediments due to the action of bacteria will occur. This creates a (soft) anoxic mud, covering the Zuiderzee deposits and protecting it from further bioturbation and erosion.
We can use the data from our measurements to estimate the yearly production rate of soft anoxic mud, and therefore, the decrease in Zuiderzee deposits surface subjected to bioturbation. Let’s assume that all sediments eroded from the Zuiderzee deposits will contribute to the stock of soft anoxic mud. We measured a TEM from the Zuiderzee deposits of 0.2 g after 8 days of bioturbation in group III, experiment day 8. If we assume all pellets on the bed were eroded during the erosion experiment, and if we assume that all erosion was in the form of pick-up of pellets, then 0.2 g of pellets were produced in 8 days of bioturbation. The density of Tubifex in Group III was 1,400 individuals/m2. The density of Tubifex in the 10 fresh samples we analyzed was 2,177 ± 883 individuals/m2. Thus, 0.31 g of Zuiderzee deposits would be eroded in the field, per microcosm surface, every 8 days, assuming a linear effect of biota density on the production of pellets. This means an erosion of about 5 g/(m2day) and therefore of about 2 kg/(m2year). Given dry bed densities of the soft anoxic mud of 200 to 300 g/l, this leads to an upper estimate for the production of soft anoxic mud of 0.9 cm per year all over the lake, and a lower estimate of 0.6 cm per year all over the lake. This would imply that the 10-cm soft anoxic mud should have been formed within a period of 10–15 years after closure of the Houtribdijk. This estimate would only hold for the first years after the closure of the Houtribdijk, when the entire surface of the Zuiderzee deposits would still be exposed to bioturbation. However, a large part of the surface of the Zuiderzee deposits became covered by the soft anoxic mud within a few years.
Another factor not yet considered is temperature. Davis (1974) found large variations in the amount of sediment pelletized by Tubifex. In most cases, the pelletization rate was found to increase with increasing temperature. The pelletization rates reported by Davis (1974), as well as the current values measured for the soft mud and Zuiderzee deposits, are presented in Fig. 14. The currently measured weight of pelletized sediments over 8 days, at a Tubifex density of 1,400 individuals/m2, has been scaled to milliliter per worm per year. The pelletization rate from the soft mud layer is well in agreement with the results reported by Davis 1974. However, the pelletization rate from the Zuiderzee deposits is much smaller than any other pelletization rate. This must be caused by the virgin state of these deposits, e.g., their high degree of consolidation.

Relationship between temperature and yearly defecation rate reported by Davis (1974), as well as the relationship we measured on the Zuiderzee deposit and on the soft Markermeer mud
Figure 14 suggests that our estimations on pellets production rates from the Zuiderzee deposits, as discussed above, are not representative of the actual pelletized material over a year, since a variation in temperature would lead to a variation in pelletization rates. Therefore, two important factors influencing bioturbation and erosion of the Zuiderzee deposits are percentage of coverage of Zuiderzee deposits and temperature. We have used these two factors for refining our estimations with a very simple model of the production of soft anoxic mud since the closure of the Houtribdijk. The water temperature in Markermeer varies between 0 and 20 °C over the year. We have modeled water temperature variation in Markermeer as: 10sin(t-π/2) + 10, which leads to 0 °C at 1 January and 20 °C at 2 July. We assume that the pelletization rate varies linearly with temperature. The total daily production of soft anoxic mud is a function of the exposed Zuiderzee deposits surface. Therefore, we have established a bimodal coverage function: the surface of the Zuiderzee deposits can be either covered by a 10-cm thick soft anoxic mud layer, or completely exposed. Thicknesses of the soft anoxic mud layer higher or smaller than 10 cm are not considered in our approach. The result of these calculations is shown in Fig. 15. Half of the surface of the Zuiderzee deposits was covered already 5 years after the closure of the Houtribdijk. In about 10 years, three quarters of the surface of the Zuiderzee deposits were covered by soft anoxic mud. Note that we did not take into account neither hydrodynamics nor consolidation processes. In reality, the hydrodynamics of the lake would not allow the 10-cm thick deposits to be created in the shallow areas, while thicknesses much higher than 10 cm can be achieved in deep regions of the lake. Vijverberg 2011 reported that there is not a clear increasing trend of turbidity since 1982, though variations in turbidity over the years depended mostly on climatology. This is consistent with our analysis, since according to our computations the coverage of the Zuiderzee deposits was approximately 60 % in 1982. The current coverage is estimated around 70 %, suggesting that the coverage remains constant, while the total amount of soft sediment continues to grow. This increase in stock of soft sediments is rather slow, since a large part of the currently exposed Zuiderzee deposits surface is sandy bed.

Evolution of Zuiderzee deposits surface exposed to bioturbation since the closure of the Houtribdijk. The larger the exposed surface, the larger the production of soft anoxic mud over the lake. Hydrodynamics and consolidation processes have not been considered for this calculation
Next to the corroboration of the hypothesis and research questions, the following physics-related facts can be concluded from the results of our experiments as well: (1) Meiobenthic bioturbators increases the rate at which Markermeer sediments are eroded up to a factor of four at bed shear stresses of 0.2 to 0.8 Pa. (2) The action of meiobenthic bioturbators do not affect the critical shear stress for erosion of the soft anoxic layer, while it does affect the critical shear stress for erosion of the Zuiderzee deposits. (3) The erosion rate increases with bioturbation time in 15 out of the 18 bioturbation experiments presented in this paper. Thus, the higher the bioturbation time, the higher the increase in erosion rate.
5 Conclusions
Our study shows that bioturbation by meiobenthic fauna drives the development of an oxic layer and the subsequent increase in erodability in the fine sediment bed of the Markermeer. Diffusion of oxygen is responsible for the oxidation of the sediments in the oxic layer, but bioturbation plays a major role in the exchange of oxygen with the water column, further increasing the depth at which oxygen can penetrate into the bed because of the increased porosity caused by the tunneling and burrowing of the meiofauna. Furthermore, the oxidation state of the sediments does not increase the erodability of the bed itself, but it is bioturbation alone that produces this increase in erodability. On the other hand, a certain oxygen level is needed to activate the meiofauna in the sediments.
The current results have important implications for the Markermeer. Bioturbation of the anoxic layer is creating a continuous input of erodible material, which is responsible of the high turbidity in Markermeer. Without the effect of biota, the erodability of the bed would be smaller, leading to re-suspension of sediments only under storm events. The specific role in the bioturbation process of Tubifex in comparison with other type of fauna was also evaluated in this paper, and through studying the way in which fauna modifies the bed. We observed two ways through which meiobenthic bioturbators can increase the erodibility of the bed. Firstly by mechanical disruption of the bed by meiobenthic bioturbators, leading to increase in porosity and water content, and a decrease in bed strength and secondly, by pelletization of the bed. Some species of meiobenthic bioturbators (e.g., Tubifex) can pelletize the bed. These pellets are easily erodible, in the form of a pick-up of pellets. The effect of this pelletization on the erodability is only temporal.
In summary, the short-term sediment dynamics in Markermeer are driven by the complex interactions between physics, chemistry, and biology. On the other hand, the long-term sediment dynamics are dominated by the increase in stock of anoxic layer. The latter is linked with bioturbation-associated erosion of the Zuiderzee deposits, as well as with the percentage of Zuiderzee deposits surface that is exposed to bioturbation. It is suggested here that the stock of anoxic layer increased rapidly during the first years after the closure of the lake, and due to a sequence of processes that was initiated by bioturbation of the Zuiderzee deposits. Meiobenthic bioturbators play therefore a major role in the long-term and short-term fine sediment dynamics in Markermeer.
References
Amaro TPF, Duineveld CA, Bergman MJN, Witbaard R, Scheffer M (2007) The consequences of changes in abundance of Callianassa subterranea and Amphiura filiformis on sediment erosion at the Frisian Front (south-eastern North Sea). Hydrobiologia 589:273–285
Amos CL, Daborn GR, Christian HA, Atkinson A, Robertson A (1992) In situ erosion measurements on fine-grained sediments from the Bay of Fundy. Mar Geol 108:175–196
Chapman P (2001) Utility and relevance of aquatic Oligochaetes in Ecological Risk Assessment. Hydrobiologia 463(149–169):2001
Coops H, Kerkum FCM, van der Berg MS, van Splunder I (2007) Submerged macrophyte vegetation and the European Water Framework Directive: assessment of status and trends in shallow, alkaline lakes in the Netherlands. Hydrobiologia 584:395–402
Davis RB (1974) Stratigraphic effects of tubificids in profundal lake sediments. Mar Eco Prog Ser 19(1974):466–488
Dniker SW, Rhoton FE, Torrent J, Smeck NE, Lal R (2003) Iron (hydr)oxide crystallinity effects on soil aggregation. Soil Sci Soc Am J 67:606–611
Fisher JB, Lick WJ (1980) Vertical mixing of lake sediments by tubified Oligochaetes. J Geophys Res 85:3997–4006
Gust, G (1989) Merhod and apparatus to generate precisely defined wall shearing stresses. US Patent 4884892 A, 5 Dec 1989
Gust, G. (1990) Method of generating precisely defined wall shearing stresses. US Patent 4973165 A, 27 Nov 1990.
Ivanov V, Chu J (2008) Applications of microorganisms to geotechnical engineering forbioclogging and biocementation of soil in situ. Rev Environ Sci Biotechnol 7:139–153
Knaapen MAF, Holzhauer H, Hulscher SJMH, Baptist MJ, and de Vries MB, van Ledden M (2003) On the modelling of biological effects on morphology in estuaries and seas. In: Third IAHR Symposium on River, Coastal and Estuarine Morphodynamics, RCEM 2003, 1–5 September 2003, Barcelona, Spain
Le Hir P, Monbet Y, Orvain F (2007) Sediment erodability in sediment transport modelling: can we account for biota effects. Cont Shelf Res 27(8):1116–1142
McCall PL (1979) The effects of deposit feeding Oligochaetes on particle size and settling velocity of Lake Erie sediments. J Sediment Res 49(3):813–818
Montserrat F, van Colen C, Degraer S, Ysebaert T, Herman PHJ (2008) Benthic community-mediated sediments dynamics. Mar Ecol Prog Ser 372:43–59
Mortimer CH (1942) The exchange of dissolved substances between mud and water in lakes. The Journal of Ecology 30(1):147–201
Noordhuis R (2010) Ecosysteem IJsselmeergebied: nog altijd onderweg. Trends en ontwikkelingen in water en natuur van het Natte Hart van. Rijkswaterstaat Waterdienst, Nederland
Noordhuis R, Houwing EJ (2003) Afname van de Driehoeksmossel in het Markermeer. RIZA rapport 2003.016
Pearson TH, Rosenberg R (1978) Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr Mar Biol Annu Rev 16:229–311
Rhoads DC, Young DK (1970) The influence of deposit-feeding organisms on sediment stability and community trophic structure. Journal of Marine Research 28(2):150–178
Royal Haskoning & Delft Hydraulics (2006), Verdiepingsslag en maatregelen slibproblematiek Markermeer – Analyse kennisleemten en inventarisatie maatregelen, authors Van Ledden et al., Royal Haskoning and WL Delft Hydraulics.
van de Bund WJ, Goedkoop W, Johnson RK (1994) Effects of deposit-feeder activity on bacterial production and abundance in profundal lake sediment. J N Am Benthol Soc 13(4):532–539
van Duin E H S (1992) Sediment transport, light and algal growth in the Markermeer—a two dimensional water quality for a shallow lake. Ph.D. thesis, Wageningen University.
van Eerden M, van Rijn S (2003) Redistribution of the Cormorant population in the Ijsselmeer area. CRGB 5:33–37
van Kessel T, de Boer G, Boderie P (2008) Calibration suspended sediment model Markermeer. Deltares report Q 4612
Van Prooijen BC, Winterwerp JC (2010) A stochastic formulation for erosion of cohesive sediments. J Geophys Res 115(C1) doi:10.1029/2008JC005189
van Rijn L (1984) Sediment transport. Part I: bed load transport. J Hydraul Eng 110(10):1431–1456
Vijverberg T (2008) Mud dynamics in the Markermeer. MSc-thesis Delft University of Technology. Delft, The Netherlands
Vijverberg T, Winterwerp JC, Aarninkhof SGJ, Drost H (2011) Fine sediment dynamics in shallow lakes and implication for design in hydraulic works. Ocean Dyn 61:187–202
Widdows J, Brinsley MD, Bowley N, Barrett C (1998) A benthic annular flume for in situ measurement of suspension feeding/biodeposition rates and erosion potential of intertidal cohesive sediments. Estuarine. Coastal Shelf Sci 46(1):27–38
Widdows J, Brinsley MD, Salkeld PN, Lucas CH (2000) Influence of biota on spatial and temporal variation in sediment erodability and material flux ona tidal flat (Westernschelde, The Netherlands). Mar Ecol Prog Ser 194:23–37
Willows R, widdows J, Wood R (1998) Influence of an infaunal bivalve on the erosion of an intertidal cohesive sediment: a flume and modelling study. Limnol Oceanogr 43(6):1132–1343
Acknowledgments
This research is part of the innovation program, Building with Nature, which ran from 2008 till the end of 2012. It is funded from different sources, among which the Subsidieregeling Innovatieketen Water (SIW), sponsored by the Netherlands Ministry of Infrastructure and Environment, and contributions of the participants to the Ecoshape consortium. The program receives co-funding from the European Fund for Regional Development and the Municipality of Dordrecht. The authors thank the anonymous reviewers of this paper, for thoroughly reading the manuscript, providing valuable comments, and very useful feedback. Moreover, the authors thank Peter Herman, Gerard Kruse, Bram van Prooijen, and Ruurd Noordhuis for the support and guidance on this paper and related work, and are grateful to Mr. Marinus Hom for helping to defaunate Markermeer samples through sterilization with gamma rays in the Nuclear Reactor Institute Delft.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Qing He
This article is part of the Topical Collection on the 11th International Conference on Cohesive Sediment Transport
Rights and permissions
About this article
Cite this article
de Lucas Pardo, M.A., Bakker, M., van Kessel, T. et al. Erodibility of soft freshwater sediments in Markermeer: the role of bioturbation by meiobenthic fauna. Ocean Dynamics 63, 1137–1150 (2013). https://doi.org/10.1007/s10236-013-0650-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10236-013-0650-0