
Abstract
Behavioural interactions between alien invasive fishes and native fishes is considered one of the drivers of native fish decline. However, there are few experimental studies on their behavioural interactions at the individual level. In this study, we investigated the behavioural interactions between the threatened native killifish Valencia letourneuxi, and the alien invasive Eastern mosquitofish Gambusia holbrooki, thought to induce the killifish’s rapid population decline. The aim was to assess aggressive behaviours, disruption in activity, stress levels, boldness, forging efficiency and shelter use by the native fish in the presence of the mosquitofish. Interspecies interactions were assessed in a three-tiered experimental setup, i.e. in an empty arena, in the presence of an artificial cover, and during feeding, using two opponents at each trial. The behaviours recorded and assessed were nips, following, approach, moving/immobile, top/bottom, erratic movement, cover use, feeding and latency to exit. The results showed that the mosquitofish was bolder, followed the native species, spent significantly less time under the cover and consumed food, as opposed to no following by the native species, significantly higher cover use and time immobile, as avoidance behaviours, and zero feeding in the presence of the mosquitofish. More significantly, the native species received direct aggression by the mosquitofish, as opposed to no aggression at all exhibited by it. Future research needs, as well as the conservation implications of our findings are briefly discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Alien invasive fish species are considered to be one of the main factors responsible for the decline of native freshwater fish species, worldwide and in the Mediterranean. However, it is often difficult to assess their exact impact, since this is often concurrent to habitat degradation (Maceda-Veiga 2013; Reid et al. 2019). Nevertheless, the theory that invasive species are the main drivers of native species decline has been supported by case studies showing the abundance of invasive species being the best single predictor of native species’ decline, with habitat degradation playing a lesser role, as in the Hermoso et al. (2011) and the Light and Marchetti (2006) studies. These indicate that freshwater fish assemblages in Mediterranean-type rivers globally may be relatively resistant to habitat perturbations, but are highly sensitive to biotic interactions with invasive species.
Alien fish species exert their negative impacts on native species through a series of mechanisms, such as predation, direct aggression, interference competition and competition for resources, such as for shelter, spawning sites and food, as well as on time investment for parental care (for a review see Leunda 2010; Ribeiro and Leunda 2012; Franco et al. 2023; Olivares-Rubio et al. 2023). Several studies on native–alien interactions in cichlids, salmonids and gobies have shown that established non-native fish exhibit higher levels of aggression than native fish (Fausch 1988; Blanchet et al. 2007; Bergstrom and Mensinger 2009, Lorenz et al. 2011, but see Archundia and Arce 2019). Aggression may lead to reduced access to resources for the native species, slowdown in their growth and reproductive disruption, ultimately leading to native fish displacement (Lopez 2017; Keller and Brown 2008; Champneys et al. 2021).
The Eastern mosquitofish, Gambusia holbrooki, an extremely successful invader now very abundant worldwide and in many Mediterranean countries, as well as its congener, the Western mosquitofish, Gambusia affinis, have been implicated in the decline of native fish species through predation, interference competition and/or exploitation competition (Pyke 2005, 2008; Caiola and de Sostoa 2005). Similarly, the decline of the two Valenciidae killifish species living in Greece (Valencia letourneuxi and Valencia robertae), i.e. fragmented, rapidly declining extant populations, is attributed to the combined degradation of their environment and the competition with G. holbrooki; a recent study has shown that killifish abundance is significantly lower and female killifish condition poorer in co-occurrence to the Eastern mosquitofish (Kalogianni et al. 2010, 2014; 2019). More specifically, Kalogianni et al. (2022) demonstrated, with the application of the Living Planet Index (LPI), a population decline of V. letourneuxi by 97.7% and of V. robertae by 91% in the period 2005–2018. However, the exact mechanisms of the impacts of the alien mosquitofish, especially in the form of agonistic interactions between the mosquitofish and the two killifishes, are unknown.
The aim of this study was to assess the agonistic interactions (in an experimental setting) between the Eastern mosquitofish G. holbrooki and the native threatened killifish V. letourneuxi in an empty arena, in the presence of artificial cover and in the presence of food using paired, naïve opponents; cover use and food consumption can indirectly be influenced by aggression. We also assessed non-agonistic interactions (i.e. boldness) that may also be important for this interspecific competition (e.g. boldness can affect foraging decisions, and/or cover use and may vary between species). This setting may provide useful insights into the behavioural interactions between a native and an alien invasive freshwater fish species during their “first encounter” in an aquatic ecosystem.
Materials and methods
Subjects, housing and acclimation
Killifish and mosquitofish subjects of both sexes were collected from localities where the species do not co-occur (Kalogianni et al. 2022) to ensure that neither had previous experience of encounters prior to the experiment, and therefore to remove the possible effect of learning and memory (Grabowska et al. 2016). Killifish were collected from a stream in Corfu Island in May 2019 (45 individuals) with an EFKO electrofishing DC unit (Honda 7 kVA generator, 150 m cable, 1.5 m anode pole, 6 A DC output, voltage range 300–600 V). Mosquitofish were collected from a small pond near Athens, Attica, in September 2019 (60 individuals) with dip nets. In the field, we recorded the following physicochemical parameters for the subsequent acclimation of the subjects in the laboratory: conductivity (μS/cm), pH, and temperature (°C).
Killifish were transferred to HCMR laboratories in individual 500 ml plastic containers with oxygen supply, while mosquitofish were transferred into a 30 L container with oxygen supply, due to the close proximity of the collection site. No mortalities were recorded during fish transfer. After acclimatization, killifish and mosquitofish were placed in separate 250 and 180 L tanks with water maintained at temperature 20 °C, pH 8.16 and conductivity 1000 µS/cm (V. letourneuxi stocking density 0.220 g/L; G. holbrooki stocking density 0.236 g/L). Artificial lighting with two T5 daylight 39W bulbs was provided for 10 h per day. A canister filter with a maximum circulation of 1150 L/h was used. Fish were provided with live Ceratophyllum sp. floating plants for shelter, while a thin layer of natural gravel was placed at the bottom of the tanks to create a naturalistic environment.
Fish were fed with commercial flakes and granules (Vipan and Vipagran, respectively, Sera GmbH, Germany) once a day ad libitum. Feeding with flakes was reduced gradually until 2 weeks prior to the experiments conducted in March 2020, when fish were fed only with granules that were used during the subsequent experiment (Stage 2, see below). At the same time, each housing tank was separated into two compartments with a perforated Plexiglas division and fish placed in one of the two compartments. After each experiment, used subjects were placed in the other compartment to monitor for mortalities and signs of post-experimental stress.
Experimental setup
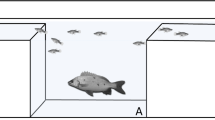
The experimental setup consisted of an aquarium (arena) measuring 36 cm length × 22 cm width × 28 cm height), where two transparent Plexiglas sliding dividers were added to create three equal-sized compartments (two side compartments and a central one, dimensions 12 × 22 × 28 cm, Fig. 1). Sliding dividers allowed the exchange of visual and chemical cues, prior to the onset of each experiment, between each pair of subjects (Raymond et al. 2015; Archundia and Arce 2019). A thin layer of gravel substrate was added at the bottom to reduce fish stress. Except of the front glass, all other sides of the aquarium were covered with opaque sheets, to facilitate recording with a camera (Logitech HD Webcam C270), while minimizing disturbance of the fish by external visual cues. The camera was placed approx. 1 m from the front glass, on which the horizontal midline was marked to facilitate the recording of fish position on the vertical axis.

Schematic drawing of the experimental arena
Fish were not fed for 24 h prior to each experiment and until after the completion of their trials. Prior to the main experiments, six pilot trials were conducted to standardize the experimental procedure.
Experimental protocol
29 trials involving 29 adult female mosquitofish and 29 adult killifish (15 females and 14 males) in pairs (i.e. 29 pairs of opponents) were conducted. Since male mosquitofish were much smaller than either female or male killifish, only female mosquitofish were used, thus reducing (though not zeroing) the effect of size. Experiments were conducted during daytime (10 am–4 pm) under natural lighting conditions (sunlight coming from a window). Experiments consisted of three stages, assessing the interactions of a pair of opponents in (a) an empty arena (Stage 1) (b) in the presence of artificial cover (Stage 2) and (c) in the presence of food (Stage 3) (following Carmona-Catot et al. 2013 and Archundia and Arce 2019 with modifications).
All fish were used once to avoid behavioural responses associated with learning, i.e. to avoid habituation or sensitization to the opponent. The physicochemical conditions of the water in the experimental arena remained constant and similar to those in the holding tanks. At the end of each trial, water in the experimental tank was renewed (50% dechlorinated tap water at 20 °C, 25% water from the mosquitofish holding aquarium and 25% water from the killifish holding aquarium, water depth 20 cm) to control for olfactory cues present in the tank from the previous experiment. The artificial cover was also rinsed thoroughly to remove any chemical cues from the previous subjects used.
At the beginning of each experiment, a mosquitofish and a killifish subject were carefully placed in the two outer compartments of the experimental tank and were allowed to acclimatize for 60 min before each trial. Placement in the outer compartments (left or right) alternated by species and killifish sex in successive trials. After acclimatization, the video recording commenced, with each trial lasting in total 55 min, and consisting of three stages, conducted sequentially. In Stage 1, the two dividers were gently raised simultaneously by pulling a string (to minimize stress), and the interaction of the two opponents in the empty arena was recorded for 15 min. Fish were then carefully returned to the two outer compartments and remained there for a 5-min interval, during which an artificial cover was placed in the central compartment for Stage 2. In Stage 2, the dividers were again lifted and fish were allowed to interact for 15 min. The artificial cover provided was an 8 × 2 cm plastic plant tied to a pebble with a nylon string that was placed approximately 4 cm from the bottom of the arena. The behaviour of the two species was again recorded for 15 min. Fish were once more returned to the two outer compartments and remained there for a 5-min interval. In Stage 3, the dividers were again lifted and the fish were allowed to enter the central compartment where a small portion of measured food granules (10 granules, mean weight ~ 25 mg) had been previously added (this food was provided to the fish already in their holding aquaria and readily consumed by the fish for a 3-week period prior to the experiments). After the end of the experiment, fish were weighted to the nearest 0.01 (with an Ohaus SC-2020 Scout scale).
The following behaviours captured in the video recording, either as number of events (n) or as duration of the given behaviour in seconds (s), were analysed: nips, i.e. bites involving physical contact between the mouth of a subject and the body of its opponent (n); following, i.e. a subject following its opponent at a distance equal or less of one body length (both opponents moving, s); approach, i.e. a subject approaching its opponent at a distance equal or less of one body length (opponent immobile, n); moving/immobile, i.e. a subject swimming or immobile respectively, for more than 2 s since both species move with small, abrupt swimming movements (s); top/bottom, i.e. a subject positioning itself at the top/bottom half of the arena (more than half of the body crossing the line, s) and erratic movement, i.e. non-linear stress-related swimming (n). Besides these behaviours that were assessed from pooling data at all three stages of each trial, we also assessed in Stage 2, cover use, i.e. at least half of the body length of a subject under the artificial cover (s) for more than 2 s, and in Stage 3, feeding, i.e. number of attempts to consume a food granule, for an indication of food competition (n). Finally, Latency to exit, i.e. time to exit (s) the side compartment was assessed as a measure of boldness (when fish exited the compartment in an erratic bout, they were excluded from this analysis).
Data processing and statistical analysis
The behaviour of each subject per stage was determined separately using the JWatcher. The observer scored behaviour for each species separately, thus it was not possible to record data blind. Data were exported and tested statistically with SPSS v25 (IBM Inc., USA). For overall behavioural analysis (data of three stages pooled, total duration 45 min) of “Nips”, “Following”, “Approach”, “Immobile” and “Erratic”, two trials (trial 16 and trial 20) were exempt, due to technical problems encountered during these trials (in Stage 2 and Stage 3 respectively). For “Latency to exit”, six trials (13, 15, 18, 19, 21, 25) were exempted from the overall behavioural analysis due to equipment failure or to subjects leaving their compartment in an erratic state. For single stage analyses (i.e. “Cover use” and Feeding”; duration: 15 min), trial 16 was exempted for cover use analysis and trial 20 for feeding analysis for the technical problems mentioned above. Data were assessed for normality using with the Shapiro–Wilk test and for homoscedasticity with Levene’s test. Body mass data were analysed with the paired t test. Since the behavioural data were non-normal and heavily skewed, non-parametric tests were used to assess differences between killifish and mosquitofish behaviour. We used paired tests since the subjects were tested in pairs in each trial, and thus killifish and mosquitofish data were not independent. We used paired Wilcoxon Signed Ranks tests for measures in which the paired differences followed a symmetric distribution (“Latency to exit”, “Approach”, “Immobile”, “Time bottom”, “Cover use”), and paired Sign tests for all other measures (“Nips”, “Following”, “Erratic”, “Darting”, “Feeding”). To examine the effect of stage (time, acknowledging the two factors are tightly linked), and since sphericity and normality were not satisfied, we used a non-parametric test for longitudinal data provided by the nparLD package (Noguchi et al. 2012) in R (v4.0.3, R Core Team 2020). This package provides a Wald-type test statistic with its associated degrees of freedom and p values for the effect of stage (time) within the mosquitofish group, and stage (time) and sex within the killifish group. Violin plots were created in R (v4.0.3, R Core Team, 2020). Medians are illustrated by thick black lines, the interquartile range (IQR) is shown within the boxes, whiskers represent data points within 1.5 × IQR and dots are outliers. All data are included as electronic supplemental material.
Results
Mosquitofish body mass (0.50 ± 0.28 g, all female) was significantly lower compared to killifish body mass (1.26 ± 0.49 g, paired t test: t = − 7.144, df = 28, p < 0.001, n = 29). Mosquitofish (female) subjects paired with female killifish had significantly lower body mass when compared with their female killifish opponents, (paired t test: t = − 6543, df = 15, p < 0.001, n = 16). Mosquitofish (female) subjects paired with male killifish had also significantly lower body mass than their male killifish opponents (paired t test: t = − 5265, df = 12, p < 0.001, n = 13).
Mosquitofish exhibited significantly lower latency to exit compared to the killifish paired Wilcoxon signed ranks test, z = − 2.616, p = 0.009, n = 23, Fig. 2).

Latency to exit (s) of the non-native mosquitofish and of the native killifish
Mosquitofish attempted nips towards their opponent were significantly higher compared to the killifish (paired sign test, standardized z = − 2.667, exact p = 0.004 n = 27, Fig. 3a). Nine mosquitofish subjects (33.3%) attempted nips, with four subjects attempting multiple nips (outliers in Fig. 3b, values: 264, 78, 42, 11), while no killifish attempted to nip their opponent. Mosquitofish also followed their killifish opponents for a significantly longer time (paired sign test, standardized z = − 2.268, exact p = 0.016, n = 27, Fig. 3b). Twenty-six percent of the mosquitofish (seven subjects) followed their opponent, while no killifish followed their opponent. It should be noted here that five of the seven mosquitofish that followed their opponent have also attempted to nip their killifish opponent.

a Nips (n) by the non-native mosquitofish and by the native killifish; b following (s) by the non-native mosquitofish and by the native killifish
Mosquitofish also approached their opponents significantly more often compared to the killifish (paired Wilcoxon signed ranks test, z = − 2.594, p = 0.009, n = 27, Fig. 4a). Mosquitofish exhibited significantly fewer erratic movements compared to the killifish (paired sign test, standardized z = 2.345, exact p = 0.017, n = 27, Fig. 4b). They were also immobile for significantly less time compared to the killifish (paired Wilcoxon signed ranks test, z = − 3.075, p = 0.002, n = 27, Fig. 4c

a Approach (n) by the non-native mosquitofish and by the native killifish; b erratic movement (n) by the non-native mosquitofish and by the native killifish; c time immobile (s) of the non-native mosquitofish and of the native killifish
In Stage 2, mosquitofish used the artificial cover, when provided, significantly less compared to the killifish (paired Wilcoxon signed ranks test, z = 2.007, p = 0.045, n = 28, Fig. 5). In Stage 3, mosquitofish also performed feeding attempts compared to no feeding attempts by the killifish (paired sign test, standardized z = − 2.846, exact p = 0.002, n = 28, Fig. 6).

Cover use by the non-native mosquitofish and by the native killifish

Feeding of the non-native mosquitofish and of the native killifish
Finally, when examining the effect of stage (time), no significant effects of stage (time) were found in the case of the mosquitofish, on any of the behaviours tested. In the case of the killifish, stage (time) had a significant effect only on the number of approaches performed (Wald-type test, χ2 = 26.389, p < 0.001, df = 2), while the effect of sex was marginally non-significant (Wald-type test, χ2 = 3.599, p = 0.058, df = 1), and there was no significant effect of stage (time) and sex interaction (Wald-type test, χ2 = 1.302, p = 0.522, df = 2). Within sex, stage (time) had a significant effect on approaches in both the female (Wald-type test, χ2 = 12.441, p = 0.002, df = 2) and male group (Wald-type test, χ2 = 14.343, p < 0.001, df = 2). There were no other effects of stage (time) or sex on the rest of the measured behaviours.
Discussion
In this experimental study, we demonstrate that the native threatened killifish received aggression by the alien invasive Eastern mosquitofish, while exhibiting no aggression towards the mosquitofish. We also show that the native killifish spent more time under cover (using the artificial shelter when provided) and exhibited no feeding at all in the presence of the mosquitofish. Finally, the killifish was overall more hesitant to exit its starting compartment compared to the mosquitofish.
In the present study, the native killifish did not exhibit any direct aggression (nips) towards their mosquitofish opponents. In contrast, the alien invasive mosquitofish were aggressive towards their killifish opponents in several occasions. This conforms to studies showing the Eastern mosquitofish, as well as the Western mosquitofish G. affinis, exhibiting higher aggression than native species (e.g. in mesocosm studies on the agonistic interactions of the Eastern mosquitofish with native Iberian toothcarps, Aphanius iberus and Valencia hispanica (Rincón et al. 2002; Carmona-Catot et al. 2013), as well as on mosquitofish interactions with native galaxiid species (Rowe et al. 2007, for a review see Rowe et al. 2008), and with other native topminnows (Sutton et al. 2013). The aggression recorded in our experimental study using pairs of opponents could be higher in the natural environment, where killifish are likely to encounter mosquitofish at higher numbers, as mosquitofish occur at much higher densities than the killifish at most of their extant habitats (Kalogianni et al. 2010; 2019; 2022).Furthermore, several studies have shown a positive relationship between mosquitofish aggression and mosquitofish density (Rowe et al. 2008; Thompson et al. 2012). In contrast, Beatty et al. (2022), suggested that aggression was a factor of mosquitofish size, corroborating previous studies (Rincón et al. 2002; Mills et al. 2004). In our study, we used only female mosquitofish, typically much larger than males (Pyke 2005), to avoid the extreme mismatch of size between male mosquitofish and killifish subjects of both sexes, and observed high levels of aggression from the females towards their killifish opponents of both sexes. However, further examination of the precise effect of size and/or sex on killifish–mosquitofish interactions is warranted. Similarly, though the focus of our study was to investigate “first encounter” responses of the two species, a study on baseline behaviour of the two species in a similar three-tiered experiment could be useful for a more general behavioural assessment of the two species.
Fish receiving aggression in the form of nips translates to high metabolic cost, since body and fin damage results in loss of fitness, reduction in swimming performance, and thus in loss of access to resources when in contest with aliens or conspecifics (Archundia and Arce 2019; Carmona-Catot et al. 2013; Chifamba and Mauru 2017). Thus, aggression received by the mosquitofish could explain the poorer condition of female V. letourneuxi in co-occurrence with the mosquitofish (Kalogianni et al. 2019). Aggression-induced damage can also increase predation risk and susceptibility to disease (Carmona-Catot et al. 2013). Aggression, finally, may also lead to reduction in reproductive success, either through swimming performance reduction (Carmona-Catot et al. 2013) or through the discontinuation of courtship of the native fishes to engage instead in aggression towards the alien species, as has been shown for the related species Valencia hispanica (Rincón et al. 2002).
In our study, mosquitofish subjects followed their opponents, as opposed to no following observed in the killifish. This is maybe an exploratory behaviour, a form of inspection of a novel potential predator and assessment of its threat (Botham et al. 2006), as both mosquitofish and killifish subjects were naïve to each other. Alternatively, it may be a low cost effort to control/modify the opponent’s behaviour (Archundia and Arce 2019). In our study, a subset of the mosquitofish subjects followed their opponents for a much longer time and most of them also initiated a high number of aggressive nips, compared to the rest of the mosquitofish subjects, which suggests that following is a prelude to direct aggression (Carmona-Catot et al. 2013).
Ιn our study, there was a marked within-group variation in direct aggression and following by the mosquitofish, with only a subset of the mosquitofish attempting nips and/or following their opponent. This is in line with previous studies reporting that some individuals consistently carried out multiple attacks while others exhibited no aggression at all. (e.g. Magellan and García-Berthou 2016). This could be explained as a variation in the “personality of the invader” based on studies exploring animal personality or behavioural syndrome and its ecological consequences in the successful establishment of the invader in new environments (for fish see Groen et al. 2012; Winandy and Denoël 2015, for a broad overview, see Sih et al. 2004). An issue to consider would be to explore a possible correlation between boldness and aggression; however, the few mosquitofish attempting the higher number of nips were not the bolder fish (i.e. with the lowest latency to exit values). Alternatively, subtle differences in mosquitofish size (in relation to killifish size), or variation in their hunger levels, could at least partially explain this large variation in mosquitofish aggressiveness. However, our data do not support this hypothesis since there were no significant correlations between number of nips and body mass difference (between mosquitofish and killifish) or feeding attempts, for the subset of fish that nipped their opponent at least once.
In our study, time immobile and erratic movement were significantly higher in killifish subjects. This may indicate an avoidance behaviour characterized by high stress and perceived risk of aggression from the mosquitofish. Similarly, Mills et al. (2004) have shown that native chub were more stationary in the presence of the more aggressive mosquitofish. Also, Sutton et al. (2013) have shown that in experimental tanks without the provision of a cover, topminnows remained motionless to avoid attracting the attention of the mosquitofish. Our results also show a statistically significant difference in terms of the exit time of the two species at the beginning of each stage (latency to exit) with the native killifish exiting much later than the alien mosquitofish. This difference could be explained either as a sign of killifish stress in the presence of mosquitofish (similarly to time immobile) or as a sign of the higher tendency of the alien mosquitofish to explore new environments (Mills et al. 2004; Bisazza and Marin 1991) compared to an overall shyer killifish temperament.
In our experimental setting when an artificial cover was provided, in the form of an artificial plant in the centre of the aquarium, the killifish spent significantly more time under cover. This conforms to the findings of Mills et al. (2004) which showed that native chubs chose protected habitats (plants), in the presence of the dominant mosquitofish. Similarly, Sutton et al. (2013) have shown that, in the presence of the mosquitofish when shelter was provided, native fish used the shelter to minimize direct interactions with an aggressive species. Food intake was observed only by mosquitofish in our experiments. Similarly, but less profoundly, Rincon et al. (2002) showed that, in the presence of adult G. holbrooki, V. hispanica showed a significant reduction in the catch rates of live Daphnia spp; based on that observation, the authors postulated that interference competition, i.e. a reduction in the access of the native species to food resources, may be a main mechanism for V. hispancia’s population decline.
In conclusion, our findings suggest that the aggression of the non-native mosquitofish towards the native killifish may well be a direct cause of the decline of several of the native species’ historical populations through displacement (Kalogianni et al. 2010, 2019, 2022). Further research integrating biotic and abiotic factors in the study of the agonistic interactions between the two species is, however, required, to identify the modulatory effects of salinity and temperature, or the interplay thereof, on these interactions (Alcaraz et al. 2008; Carmona-Catot et al. 2013; Lorenz et al. 2016; Lopez et al. 2018), especially under current climate change scenarios. From a conservation point of view, habitat degradation coupled with water stress due to climate change will increase the encounters between the mosquitofish and threatened native fishes accelerating their decline. Thus, prevention of the spread of the mosquitofish in the last remaining habitats of the Corfu killifish that are still not compromised by the invasive mosquitofish, where we expect individual “first encounters” (Brodin et al. 2019; Magellan et al. 2019) similar to those demonstrated in our experimental setup, should be a managerial priority, as suggested elsewhere (Kalogianni et al. 2022).
Data availability
The datasets are available upon reasonable request.
References
Alcaraz C, Bisazza A, García-Berthou E (2008) Salinity mediates the competitive interactions between invasive mosquitofish and an endangered fish. Oecologia 155(1):205–213. https://doi.org/10.1007/s00442-007-0899-4
Archundia M, Arce E (2019) Fighting behaviour in native fish: the Mexican mojarra (Cichlasoma istlanum) wins when confronted with the non-native convict cichlid fish (Amatitlania nigrofasciata). J Ethol 37(1):67–73. https://doi.org/10.1007/s10164-018-0569-5
Beatty S J, Lear KO, Allen MG, Lymbery AJ, Tweedley JR, Morgan D L (2022) What factors influence fin‐nipping damage by the invasive Gambusia holbrooki (Poeciliidae) on native fishes in riverine systems?. Freshw Biol 67(2): 325–337
Bergstrom MA, Mensinger AF (2009) Interspecific resource competition between the invasive round goby and three native species: logperch, slimy sculpin, and spoonhead sculpin. Trans Am Fish Soc 138(5):1009–1017. https://doi.org/10.1577/T08-095
Bisazza A, Marin G (1991) Male size and female mate choice in the eastern mosquitofish (Gambusia holbrooki: Poeciliidae). Copeia. https://doi.org/10.2307/1446400
Blanchet S, Loot G, Grenouillet G, Brosse S (2007) Competitive interactions between native and exotic salmonids: a combined field and laboratory demonstration. Ecol Freshw Fish 16(2):133–143. https://doi.org/10.1111/j.1600-0633.2006.00205.x
Botham MS, Kerfoot CJ, Louca V, Krause J (2006) The effects of different predator species on antipredator behavior in the Trinidadian guppy, Poecilia. Naturwissenschaften 93:431–439. https://doi.org/10.1007/s00114-006-0131-0
Brodin T, Fogarty S, Sih A, Cote J (2019) Personality-dependent survival of the invasive mosquitofish: being social can be deadly. Aquat Invasions 14(3):465–477. https://doi.org/10.3391/ai.2019.14.3.06
Caiola N, De Sostoa A (2005) Possible reasons for the decline of two native toothcarps in the Iberian Peninsula: evidence of competition with the introduced Eastern mosquitofish. J Appl Ichthyol 21(4):358–363. https://doi.org/10.1111/j.1439-0426.2005.00684.x
Carmona-Catot G, Magellan K, García-Berthou E (2013) Temperature-specific competition between invasive mosquitofish and an endangered cyprinodontid fish. PLoS ONE 8(1):e54734. https://doi.org/10.1371/journal.pone.0054734
Champneys T, Genner MJ, Ioannou CC (2021) Invasive Nile tilapia dominates a threatened indigenous tilapia in competition over shelter. Hydrobiologia 848(16):3747–3762. https://doi.org/10.1007/s10750-020-04341-8
Chifamba PC, Mauru T (2017) Comparative aggression and dominance of Oreochromis niloticus (Linnaeus, 1758) and Oreochromis mortimeri (Trewavas, 1966) from paired contest in aquaria. Hydrobiologia 788(1):193–203. https://doi.org/10.1007/s10750-016-2997-y
Fausch KD (1988) Tests of competition between native and introduced salmonids in streams: what have we learned? Can J Fish Aquat Sci 45(12):2238–2246. https://doi.org/10.1139/f88-26
Franco M, Arce E, Mercado-Silva N, Córdoba-Aguilar A, Ramírez-Rodríguez R (2023) Invasive cichlids (Teleostei: Cichliformes) in the Amacuzac River, Mexico: Implications for the behavioral ecology of the native Mexican mojarra Amphilophus istlanus. Water Biol Secur 100182
Groen M, Sopinka NM, Marentette JR, Reddon AR, Brownscombe JW, Fox MG, Marsh-Rollo SE, Balshine S (2012) Is there a role for aggression in round goby invasion fronts? Behaviour 149(7):685–703. https://doi.org/10.1163/1568539X-00002998
Hermoso V, Clavero M, Blanco-Garrido F, Prenda J (2011) Invasive species and habitat degradation in Iberian streams: an analysis of their role in freshwater fish diversity loss. Ecol Appl 21(1):175–188. https://doi.org/10.1890/09-2011
Jermaczc GH, Coppde GH (2016) Interspecific competition for a shelter between non-native racer goby and native European bullhead under experimental conditions–effects of season, fish size and light conditions. Limnologica 56:30–38. https://doi.org/10.1016/j.limno.2015.11.004
Kalogianni E, Giakoumi S, Andriopoulou A, Chatzinikolaou Y (2010) Feeding ecology of the critically endangered Valencia letourneuxi (Valenciidae). Aquat Ecol 44(1):289–299. https://doi.org/10.1007/s10452-009-9253-8
Kalogianni E, Giakoumi S, Andriopoulou A, Chatzinikolaou Y (2014) Prey utilisation and trophic overlap between the non native mosquitofish and a native fish in two Mediterranean rivers. Mediterranean Mar Sci 15(2):287–301
Kalogianni E, Koutsikos N, Vardakas L, Giakoumi S, Chatzinikolaou Y, Oikonomou A (2019) Impacts of the alien mosquitofish on the abundance and condition of two Mediterranean native fish. Mediterr Mar Sci 20(4):727–735
Kalogianni E, Kapakos Y, Oikonomou A, Giakoumi S, Zimmerman B (2022) Dramatic decline of two freshwater killifishes, main anthropogenic drivers and appropriate conservation actions. J Nat Conserv 67:126191. https://doi.org/10.1016/j.jnc.2022.12619
Keller K, Brown C (2008) Behavioural interactions between the introduced plague minnow Gambusia holbrooki and the vulnerable native Australian ornate rainbowfish Rhadinocentrus ornatus, under experimental conditions. J Fish Biol 73(7):1714–1729. https://doi.org/10.1111/j.1095-8649.2008.02045.x
Leunda PM (2010) Impacts of non-native fishes on Iberian freshwater ichthyofauna: current knowledge and gaps. Aquat Invasions 5(3):239–262. https://doi.org/10.3391/ai.2010.5.3.03
Light T, Marchetti MP (2006) Distinguishing between invasions and habitat changes as drivers of biodiversity loss among California’s freshwater fishes. Conserv Biol 21:434–446. https://doi.org/10.1111/j.1523-1739.2006.00643.x
Lopez LK, Davis AR, Wong MY (2018) Behavioral interactions under multiple stressors: temperature and salinity mediate aggression between an invasive and a native. Biol Invasions 20(2):487–499. https://doi.org/10.1007/s10530-017-1552-8
Lopez LK (2017) The context-dependency of predator-prey and competitive interactions between the invasive eastern mosquitofish, Gambusia holbrooki, and native Australian freshwater fauna, Doctor of Philosophy thesis, School of Biological Sciences, University of Wollongong. https://ro.uow.edu.au/theses1/106. Accessed 30 June 2022
Lorenz OT, O’Connell MT, Schofield PJ (2011) Aggressive interactions between the invasive Rio Grande cichlid (Herichthys cyanoguttatus) and native bluegill (Lepomis macrochirus), with notes on redspotted sunfish (Lepomis miniatus). J Ethol 29:39–46
Lorenz OT, Riccobono SA, Smith P (2016) Effects of salinity on the survival and aggression of the invasive Rio Grande cichlid (Herichthys cyanoguttatus). Mar Freshw Behav Physiol 49(1):1–8
Maceda-Veiga A (2013) Towards the conservation of freshwater fish: Iberian Rivers as an example of threats and management practices. Rev Fish Biol Fish 23(1):1–22. https://doi.org/10.1007/s11160-012-9275-5
Magellan K, García-Berthou E (2016) Experimental evidence for the use of artificial refugia to mitigate the impacts of invasive Gambusia holbrooki on an endangered fish. Biol Invasions 18(3):873–882. https://doi.org/10.1007/s10530-016-1057-x
Magellan K, Alexander M, Deacon A, Wong M (2019) Behaviour and aquatic invasions in the 21st century: progress, trends and future research. Aquat Invasions 14(3):412–416. https://doi.org/10.3391/ai.2019.14.3.01
Mills MD, Rader RB, Belk MC (2004) Complex interactions between native and invasive fish: the simultaneous effects of multiple negative interactions. Oecologia 141(4):713–772. https://doi.org/10.1007/s00442-004-1695-z
Noguchi K, Gel YR, Brunner E, Konietschke F (2012) nparLD: an R software package for the nonparametric analysis of longitudinal data in factorial experiments. J Stat Softw. https://doi.org/10.18637/jss.v050.i12
Olivares-Rubio HF, Arce E, Burciaga LM, Franco M, Osorio-Beristain M (2024) Lead has different impacts on behavior of the native Mexican mojarra and invasive convict cichlid. Hydrobiologia, 851(1):147–159
Pyke GH (2005) A review of the biology of Gambusia affinis and G. holbrooki. Rev Fish Biol Fish 15(4):339–365. https://doi.org/10.1007/s11160-006-6394-x
Pyke GH (2008) Plague minnow or mosquito fish? A review of the biology and impacts of introduced Gambusia species. Ann Rev Ecol Evol Syst 39:171–191
Raymond WW, Albins MA, Pusack TJ (2015) Competitive interactions for shelter between invasive Pacific red lionfish and native Nassau grouper. Environ Biol Fishes 98(1):57–65. https://doi.org/10.1007/s10641-014-0236-9
Reid AJ, Carlson AK, Creed IF, Eliason EJ, Gell PA, Johnson PT, Cooke SJ (2019) Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol Rev 94(3):849–873
Ribeiro F, Leunda PM (2012) Non-native fish impacts on Mediterranean freshwater ecosystems: current knowledge and research needs. Fish Manage Ecol 19:142–156. https://doi.org/10.1111/j.1365-2400.2011.00842.x
Rincón PA, Correas AM, Morcillo F, Risueño P, Lobón-Cerviá J (2002) Interaction between the introduced eastern mosquitofish and two autochthonous Spanish toothcarps. J J Fish Biol 61(6):1560–1585. https://doi.org/10.1111/j.1095-8649.2002.tb02498.x
Rowe DK, Smith JP, Baker C (2007) Agonistic interactions between Gambusia affinis and Galaxias maculatus: implications for whitebait fisheries in New Zealand rivers. J Appl Ichthyol 23(6):668–674. https://doi.org/10.1111/j.1439-0426.2007.00912.x
Rowe DK, Moore A, Giorgetti A, Maclean C, Grace P, Wadhwa S, Cooke J (2008) Review of the impacts of gambusia, redfin perch, tench, roach, yellowfin goby and streaked goby in Australia
Sih A, Bell AM, Johnson JC, Ziemba RE (2004) Behavioral syndromes: an integrative overview. Q Rev Biol 79(3):241–277
Sutton TM, Zeiber RA, Fisher BE (2013) Agonistic behavioral interactions between introduced western mosquitofish and native topminnows. J Freshw Ecol 28(1):1–16. https://doi.org/10.1080/02705060.2012.688492
Thompson KA, Hill JE, Nico G (2012) Eastern mosquitofish resists invasion by nonindigenous poeciliids through agonistic behaviors. Biol Invasions 14(7):1515–1529. https://doi.org/10.1007/s10530-012-0176-2
Winandy L, Denoël M (2015) The aggressive personality of an introduced fish affects foraging behavior in a polymorphic newt. Behav Ecol 26(6):1528–1536. https://doi.org/10.1093/beheco/arv101
Acknowledgements
We wish to thank P. Kouraklis for his assistance in fish collection and transportation to HCMR and B. Zimmerman for English language help and for proof reading the ms. This work forms part of the PhD thesis of Y. Kapakos at the Department of Applied Hydrobiology, Agricultural University of Athens (AUA), Greece.
Funding
This study was conducted within the project DECAGON, funded by the A.G. Leventis Foundation, through the London Zoological Society.
Author information
Authors and Affiliations
Contributions
Yiannis Kapakos: conceptualization, methodology, software, formal analysis, investigation, writing, visualization. Ioannis Leris: conceptualization, methodology, software, formal analysis, writing, visualization. Brian Zimmerman: Nafsika Karakatsouli: supervision, writing. Eleni Kalogianni: conceptualization, methodology, resources, writing, supervision, project administration, funding acquisition.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare they have no conflict of interest.
Ethical approval
All experimental procedures were conducted according to the Greek law and conform to ASAB ethical guidelines. HCMR had secured all necessary permits for fish collection from the Greek Ministry of Environment, Energy and Climate Change (permit 9ZE24653Π-ΖΟ6, HCMR Research Ethics Committee under development during the implementation of this work and thus no specific licence to conduct the experiment was required). We conducted pilot tests before the main experiment to fine-tune the setup and procedure. During these tests, the observed level of aggression between the two species was not severe. In the main experiment, we used a long acclimation period (60 min) prior to each trial, and placed gravel on the bottom of the test tank, to reduce stress for both subjects. The experimenter was operating behind an opaque cover, and observed the trials remotely through a camera, to minimize disturbance. After each trial, used fish were placed in a separate compartment in their home tank for monitoring of their health and recovery. We did not observe any bleeding or scale loss during or after the trials, only minor fin damage was visible in some cases. Should there have been cases of severe aggression, the subjects would have been immediately separated using a stick (which was used to separate and guide them into their respective compartment at the end of each stage), and then returned to the used compartment of their housing tank for monitoring. After the conclusion of the experiments, no mortalities or other adverse effects were observed on the subjects, which were maintained in HCMR facilities for breeding purposes and future experimental use.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Kapakos, Y., Leris, I., Karakatsouli, N. et al. Behavioural interactions between a threatened native killifish and the alien invasive Eastern mosquitofish. J Ethol 42, 97–106 (2024). https://doi.org/10.1007/s10164-024-00807-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10164-024-00807-7