
Abstract
In many animals, investment in anti-predator traits can affect reproductive success. Conversely, males that invest more resources in mating success may have relatively fewer resources to devote to anti-predator traits, leading to increased predation risk. Although previous studies have reported a trade-off in investment between anti-predator traits and reproductive traits in male animals, few studies have specifically investigated the effects of anti-predator behavior on male reproductive traits. Many animals engage in death-feigning as an anti-predator behavior. Herein, we investigated the relationship between the death-feigning behavior and pre- and post-copulatory reproductive traits of male red flour beetles, Tribolium castaneum. In a previous study, we used artificial selection to establish T. castaneum strains with a genetically longer (L-strain) or shorter (S-strain) duration of death-feigning behavior. In the present study, we compared the attractiveness (a pre-copulatory trait) and paternity success (a post-copulatory trait) between L- and S-strain males. The results showed no significant difference in attractiveness or paternity success between the two strains. The results suggest that death-feigning behavior is not correlated with pre- or post-copulatory reproductive traits in male T. castaneum.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Both predation pressure and sexual selection exert evolutionary pressure on many animal species (e.g., Edmunds 1974; Blum and Blum 1979; Lima and Dill 1990; Sih 1992; Lima 1998). However, predation pressure is thought to be opposed by sexual selection (e.g., Magnhagen 1991; Andersson 1994; Hine et al. 2011; Sharma et al. 2012). This antagonism between natural and sexual selection has been reported in several previous studies. For example, in the wood tiger moth Parasemia plantaginis, males with more conspicuous warning coloration had increased survival rates but were less successful at mating than inconspicuous males (Nokelainen et al. 2012). This suggests that a trade-off exists between warning coloration and mating success because investment in warning coloration may decrease the resources for mating success in males. Furthermore, in the guppy Poecilia reticulata, males with conspicuous ornaments attracted more females but were at increased risk of predation due to their conspicuous ornamentation (e.g., Breden and Stoner 1987; Godin and McDonough 2003). Although previous studies have investigated the relationship between anti-predator and reproductive traits with a focus on conspicuous traits, such as coloration, few studies have investigated the effects of anti-predator behavior on reproductive traits in males.
Prey species often exhibit death-feigning behavior [thanatosis or tonic immobility (e.g., Miyatake et al. 2004; Ruxton et al. 2018)] in response to the threat of predation. Death-feigning behavior is observed in a wide range of animal taxa (reviewed in Humphreys and Ruxton 2018). In our previous study (Miyatake et al. 2004), two-way artificial selection of the duration of death-feigning was carried out in the red flour beetle, Tribolium castaneum, resulting in the establishment of two strains with a genetically longer (L-strain) or shorter (S-strain) duration of death-feigning behavior. L-strain beetles had a higher survival rate than S-strain beetles when encountering the jumping spider Hasarius adansoni, a model predator of T. castaneum (Miyatake et al. 2004). Moreover, L-strain males had lower mating success than S-strain males (Nakayama and Miyatake 2010a). These results suggest that a longer duration of death-feigning is adaptive for predator avoidance but costly in terms of the mating success of male T. castaneum. That is, males that invest more in death-feigning behavior might have a decreased investment in pre-copulatory reproductive traits. Attractiveness is an indicator of investment in pre-copulatory reproductive traits, and males investing more in attractiveness have increased mating success (Houde 1997; Zuk and Simmons 1997). Therefore, L-strain males may have a decreased investment in attractiveness compared with S-strain males, due to a greater investment in death-feigning behavior. In other words, S-strain males may be more attractive than L-strain males.
In polygamous species, mating opportunities do not ensure reproductive success for males. In addition to pre-copulatory male–male competition, sperm competition and other forms of post-copulatory male–male competition for egg fertilization (paternity success) are important for male reproductive success (e.g., Parker 1970; Eberhard 1996; Birkhead and Møller 1998; Simmons 2001). If males that invest more resources in anti-predator behavior decrease their investment in reproductive traits, they may have decreased paternity success as well as mating success. Therefore, males that invest more resources in anti-predator behavior may increase their investment in paternity success, but not mating success. Although investment in anti-predator behavior may affect pre- and post-copulatory reproductive traits in males, few studies have investigated this relationship (but see Wilber 1989; Jivoff 1997).
In the present study, we investigated the correlation between anti-predator behavior and reproductive traits in males using death-feigning strains (L- and S-strains) of Tribolium castaneum (Herbst, 1797). We compared the attractiveness, as an indicator of pre-copulatory reproductive traits, and paternity success, as an indicator of post-copulatory reproductive traits, of L- and S-strain males. In T. castaneum, females show a preference for males with high olfactory attractiveness, as indicated by the level of 4,8-dimethyldecanal (DMD) (Suzuki 1980; Bloch Quazi et al. 1998). We therefore tested the following two hypotheses. First, if L-strain males have a decreased investment in reproductive traits, they will produce less DMD and have a lower paternity success than S-strain males. Second, if there is a trade-off between pre- and post-copulatory reproductive traits, S-strain males will have higher levels of DMD, but therefore may have decreased resources for paternity success. On the other hand, if L-strain males decrease investment in DMD, they may have increased paternity success.
Materials and methods
Insect lines and artificial selection
The stock population of T. castaneum used in the present study has been reared in the laboratory at Okayama University for over 10 years. The beetles are fed a blend of whole wheat flour and brewer’s yeast at a 19:1 ratio. They are kept in an incubator (Sanyo, Tokyo) maintained at 25 °C under a 16-h light:8-h dark (lights on at 0700 hours, lights off at 2300 hours) cycle.
Artificial selection of duration of death-feigning behavior was carried out for over 40 generations (Miyatake et al. 2004; Matsumura and Miyatake 2018). Details of this selection procedure are described in Miyatake et al. (2004). In brief, 100 virgin males (14–21 days old) and 100 virgin females (14–21 days old) were chosen at random from the stock population (base strain) and placed into individual wells of 48-well plates with food. The next day, each beetle was placed on a ceramic dish, and its abdomen was stimulated with a fine stick. We defined the onset of death-feigning as the point when the beetle was observed to stiffen in response to this stimulation. We recorded the duration of feigning using a stopwatch. The ten males and ten females with the longest duration of death-feigning were collected and propagated as the L-strain, whereas the ten males and ten females with the shortest duration of death-feigning were collected and propagated as the S-strain. The offspring of each strain were reared in an incubator as described above. When the next generation of adults emerged, we measured the duration of death-feigning and selected those with the longest and shortest death-feigning durations in the same manner as before. We repeated this procedure for over 40 generations. We simultaneously created two replicates of each line in this selection experiment (LA, SA, LB, and SB).
In the present study, we measured the duration of death-feigning behavior in beetles of the L- and S-strains to assess the results of our artificial selection procedure. We chose 40 virgin males (21–35 days old) of the L- and S-strains at random and placed them into individual wells in a 48-well plate with food. On the next day, we measured the duration of death-feigning in these beetles by the method described above.
Attractiveness
L- and S-strain males were randomly chosen, and each beetle was placed in a glass vial (diameter, 20 mm; height, 50 mm) with food and a piece of filter paper (10 mm × 10 mm). These vials were maintained in an incubator, as described above, for 21 days. Previous studies have reported that this method allows olfactory attractants, including DMD, to adhere to the filter paper (Boake and Wade 1984; Boake 1985; Lewis and Austad 1994). After 21 days, filter papers with DMD were removed from the incubator and used for a test of male attractiveness. The filter paper was placed on the edge of a petri dish, and a randomly selected female of the T. castaneum base strain was positioned on the opposite side. We recorded the time from when the female was released in the petri dish until she touched the filter paper. The recorded time was defined as male attractiveness (i.e., shorter latency indicates higher attractiveness). The test was continued for 500 s. If the female did not touch the filter paper after this time, we recorded the time as 500 s. As a control treatment, the attractiveness test was also performed using filter paper subjected to the same incubation regime but without exposure to male beetles. All experiments were conducted in the laboratory at 25 °C between 1200 and 1800 hours.
Paternity success
To assess paternity success, we measured sperm defense (P1) and sperm offence (P2). P1 is the ability of a male’s sperm to enable paternity after a female mates with a second male. P2 indicates the ability of a second male’s sperm to enable paternity when a female has already mated with another male. In this experiment, we used a black mutant strain as a genetic marker. The black strain has a black body, which is semi-dominant over normal body color in T. castaneum (Sokoloff et al. 1960). The black strain has been used in many previous studies to investigate paternity success in T. castaneum (e.g., Schlager 1960; Lewis and Austad 1990, 1994; Edvardsson and Arnqvist 2000; Bernasconi and Keller 2001; Fedina and Lewis 2006, 2008; Lewis et al. 2012). In the test for P1, virgin males (14–21 days old) were randomly selected from the L- and S-strains, and each male was paired with a virgin base strain female (14–21 days old). After mating, the female was mated again with a virgin male (14–21 days old) of the black strain. The female was kept in a petri dish (diameter, 60 mm; height, 10 mm) with food for 1 week to allow oviposition. Offspring were reared in an incubator as described above. When the adults of the offspring emerged, we counted the number of beetles with a normal or black body color. Paternity success was defined as the proportion of offspring with normal coloration. In the test for P2, a base strain female was first mated with a black strain male and then with a L- or S-strain male. As in the first experiment, the female was allowed to oviposit in a petri dish, and offspring were reared in incubators. The colors of the offspring were recorded, and paternity success was calculated by the same method as before. All experiments were conducted in the laboratory at 25 °C.
Statistical analysis
For analysis of the duration of death-feigning behavior, we used a generalized linear mixed model (GLMM) with a log link function. In this analysis, GLMM was assumed to have a gamma distribution; strains were the fixed effect, and replicate lines the random effect. We performed a survival analysis with Cox proportional hazards regression and likelihood ratio test to analyze the results of the attractiveness test, as we had set an upper limit to the time data (500 s). In this analysis, the strain was the fixed effect, and the replicate line was the random effect. We also performed a post hoc Tukey’s honest significance difference (HSD) test. For analysis of paternity success, we used a GLMM with a binomial distribution, strain and mating order (P1 and P2) as the fixed effects, and replicate line as the random effect. Analyses for death-feigning and paternity were conducted in R (R Core Team 2017) version 3.4.3, and analysis for attractiveness was conducted in JMP version 12.2.0 (SAS Institute 2015).
Results
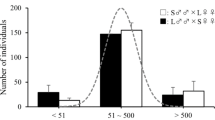
Males of the L-strain (n = 30) showed a significantly longer duration of death-feigning than those of the S-strain (n = 30) males (χ2= 235.42, p < 0.0001; Fig. 1; Table 1). In the attractiveness test, females showed a shorter response latency to touching filter paper containing DMD from males than to touching the control filter paper (n = 15) without DMD (Fig. 2). That is, females were attracted more by filter paper with DMD than by filter paper without DMD. There was a significant effect of treatments on attractiveness, and the post hoc test showed that the time to touching the filter paper by the females was significantly shorter in the L- (n = 34) and S-strains (n = 39) than in the control, but there was no significant difference between the L- and S-strains (Tukey’s HSD test: p > 0.05; Fig. 2, Table 1). In the test of paternity success, both the L- (P1, n = 56; P2, n = 207) and S-strains (P1, n = 55; P2, n = 211) had a significantly higher P2 than P1 (χ2 = 153.31, p < 0.0001), but there was no significant difference in paternity success between the two strains (χ2 = 0.27, p = 0.6049; Fig. 3, Table 1). The interaction between strain and mating order also had no significant effect (χ2 = 0.11, p = 0.7429; Fig. 3, Table 1).

Duration of death-feigning behavior in males. Black bars L-strain, white bars S-strain. Error bars indicate SE. Numbers in parentheses indicate sample size

Attractiveness (measured as time from release of female beetle to touching of filter paper) of males of L- and S-strains, including the replicate lines (a, b) and the control (filter paper without 4,8-dimethyldecanal). Shorter latency indicates higher attractiveness. Error bars indicate SE. Numbers in parentheses indicate sample size

Paternity success [P1 (a), P2 (b)] of male beetles. Black bars L-strain, white bars S-strain. Error bars indicate SE. Numbers in parentheses indicate sample size
Discussion
We investigated the effects of artificial selection of death-feigning duration on the pre- and post-copulatory traits of male T. castaneum. We found no significant difference in the attractiveness or paternity success of the L- and S-strains. These results suggest that investment in death-feigning behavior does not affect the reproductive traits of male red flour beetles.
In T. castaneum, males investing more in DMD production for olfactory attractiveness increased their mating success (Suzuki 1980; Bloch Quazi et al. 1998). We first considered the hypothesis that if males of the L-strain have decreased investment in reproductive traits, they would have decreased levels of DMD and decreased paternity success compared with the S-strain. However, our results revealed no significant difference in attractiveness between the L- and S-strains. A previous study showed that L-strain males had lower mating success than S-strain males (Nakayama and Miyatake 2010a). Furthermore, in the adzuki bean beetle Callosobruchus chinensis, males with a genetically determined longer duration of death-feigning had decreased mating success compared with males with a shorter duration of death-feigning (Nakayama and Miyatake 2010b). These studies suggested that males with more investment in death-feigning behavior had less investment in pre-copulatory reproductive traits. Therefore, the present finding raises the question of why S-strain males have higher mating success than L-strain males, despite no difference in olfactory attractiveness. In many animals, the activity level of males affects their encounters with females, and more active males often have higher mating success than inactive males (e.g., Parker 1978; Thornhill and Alcock 1983; Clutton-Brock and Parker 1992; Andersson 1994; Fromhage et al. 2016). A previous study found that S-strain T. castaneum were significantly more active than L-strain T. castaneum (Miyatake et al. 2008). The same difference between L- and S-strains has also been reported for Tribolium confusum (Nakayama et al. 2012) and C. chinensis (Ohno and Miyatake 2007). Mating success in T. castaneum may be affected by male activity level rather than attractiveness. Our results are unlikely due to errors in our methods, as we did find that both the L- and S-strains were significantly more attractive than individuals in the control treatment (without DMD). However, we did not measure the actual amount of DMD released by male beetles in this study, thus additional studies comparing the actual amount of DMD released by L- and S-strains are needed. Males of T. castaneum rub females with their legs during copulatory courtship (Eberhard 1996; Bloch Quazi et al. 1998), and we need additional research to compare the courtship behavior between the two strains.
We also considered a second hypothesis: if there is a trade-off between pre- and post-copulatory reproductive traits, S-strain males may have increased levels of DMD, while L-strain males may have increased paternity success. However, we found no difference in paternity success between the L- and S-strains. Many previous studies in animal species have reported that males with increased mating success have decreased paternity success (e.g. Parker 1970; Eberhard 1996; Birkhead and Møller 1998; Simmons 2001). That is, males investing more in pre-copulatory reproductive traits often decrease their resources for post-copulatory male–male competition for fertilization of eggs (i.e., sperm competition). In the present study, the two selected strains had the same level of attractiveness, with apparently similar levels of investment in paternity success. On the other hand, both strains had significantly higher values of P2 than P1, which concurs with the results of many previous studies in T. castaneum (e.g. Schlager 1960; Lewis and Austad 1990; Fedina and Lewis 2004). This indicates that there were no problems regarding the methods or results of this study.
Individuals that invest more in anti-predator traits often have fewer resources to invest in other traits. In fact, previous studies have reported a trade-off between predation avoidance and reproduction in males (e.g., Breden and Stoner 1987; Godin and McDonough 2003; Nokelainen et al. 2012). In the present study, we found that death-feigning behavior was not correlated with pre- or post-copulatory reproductive traits in males. However, a previous study reported higher mating success in males of the S-strain than the L-strain (Nakayama and Miyatake 2010a), suggesting a trade-off between death-feigning duration and reproductive success in male T. castaneum. This fitness cost of death-feigning behavior may lead to the maintenance of variation in death-feigning behavior within T. castaneum populations. The present study investigated the relationship between death-feigning behavior and reproductive success only in males, and additional studies are needed to investigate the effects of artificial selection of death-feigning duration on female reproductive traits.
References
Andersson M (1994) Monographs in behavior and ecology: sexual selection. Princeton University Press, Princeton
Bernasconi G, Keller L (2001) Female polyandry affects their sons’ reproductive success in the red flour beetle Tribolium castaneum. J Evol Biol 14:186–193
Birkhead TR, Møller AP (1998) Sperm competition and sexual selection. Academic Press, London
Bloch Quazi MC, Boake CRB, Lewis SM (1998) The femoral setiferous glands of Tribolium castaneum males and production of the pheromone 4,8-dimethyldecanal. Entomol Exp Appl 89:313–317
Blum MS, Blum NA (1979) Sexual selection and reproductive competition in insects. Academic Press, New York
Boake CRB (1985) Genetic consequences of mate choice: a quantitative genetic method for testing sexual selection theory. Science 227:1061–1063
Boake CRB, Wade MJ (1984) Populations of the red flour beetle Tribolium castaneum (Coleoptera: Tenebrionidae) differ in their sensitivity to aggregation pheromones. Environ Entomol 13:1182–1185
Breden F, Stoner G (1987) Male predation risk determines female preference in the Trinidad guppy. Nature 329:831–833
Clutton-Brock TH, Parker GA (1992) Potential reproductive rates and the operation of sexual selection. Quart Rev Biol 67:437–456
Eberhard WG (1996) Female control: sexual selection by cryptic female choice. Princeton University Press, Princeton
Edmunds M (1974) Defence in animals. Longman, London
Edvardsson M, Arnqvist G (2000) Copulatory courtship and cryptic female choice in red flour beetles Tribolium castaneum. Proc R Soc B 267:559–563
Fedina TY, Lewis SM (2004) Female influence over offspring paternity in the red flour beetle Tribolium castaneum. Proc R Soc B 271:1393–1399
Fedina TY, Lewis SM (2006) Proximal traits and mechanisms for biasing paternity in the red flour beetle Tribolium castaneum (Coleoptera: Tenebrionidae). Behav Ecol Sociobiol 60:844–853
Fedina TY, Lewis SM (2008) An integrative view of sexual selection in Tribolium flour beetles. Biol Rev 83:151–171
Fromhage L, Jennions M, Kokko H (2016) The evolution of sex roles in mate searching. Evolution 70:617–624
Godin JGJ, McDonough HE (2003) Predator preference for brightly colored males in the guppy: a viability cost for a sexually selected trait. Behav Ecol 14:194–200
Hine E, McGuigan K, Blows MW (2011) Natural selection stops the evolution of male attractiveness. Proc Natl Acad Sci USA 108:3659–3664
Houde AE (1997) Sex, color and mate choice in guppies. Princeton University Press, Princeton
Humphreys RK, Ruxton GD (2018) A review of thanatosis (death-feigning) as an anti-predator behavior. Behav Ecol Sociobiol 72:22
Jivoff P (1997) The relative roles of predation and sperm competition on the duration of the post-copulatory association between the sexes in the blue crab, Callinectes sapidus. Behav Ecol Sociobiol 40:175–185
Lewis SM, Austad SN (1990) Sources of intraspecific variation in sperm precedence in red flour beetles. Am Nat 135:351–359
Lewis SM, Austad SN (1994) Sexual selection in flour beetles: the relationship between sperm precedence and male olfactory attractiveness. Behav Ecol 5:223–224
Lewis SM, Tigreros N, Fedina T, Ming QL (2012) Genetic and nutritional effects on male traits and reproductive performance in Tribolium flour beetles. J Evol Biol 25:438–451
Lima SL (1998) Nonlethal effects in the ecology of predator-prey interactions. Bioscience 48:25–34
Lima SL, Dill LM (1990) Behavioral decisions made under the risk of predation: a review and prospectus. Can J Zool 68:619–640
Magnhagen C (1991) Predation risk as a cost of reproduction. Trends Ecol Evol 6:183–186
Matsumura K, Miyatake T (2018) Responses to relaxed and reverse selection in strains artificially selected for duration of death-feigning behavior in the red flour beetle, Tribolium castaneum. J Ethol 36:161–168
Miyatake T, Katayama K, Takeda Y, Nakashima A, Sugita A, Mizumoto M (2004) Is death-feigning adaptive? Heritable variation in fitness difference of death-feigning behavior. Proc R Soc B 271:2293–2296
Miyatake T, Tabuchi K, Sasaki K, Okada K, Katayama K, Moriya S (2008) Pleiotropic antipredator strategies, fleeing and feigning death, correlated with dopamine levels in Tribolium castaneum. Anim Behav 75:113–121
Nakayama S, Miyatake T (2010a) A behavioral syndrome in the adzuki bean beetle: genetic correlation among death-feigning, activity, and mating behavior. Ethology 116:108–112
Nakayama S, Miyatake T (2010b) Genetic trade-off between abilities to avoid attack and to mate: a cost of tonic immobility. Biol Lett 6:18–20
Nakayama S, Sasaki K, Matsumura K, Lewis Z, Miyatake T (2012) Dopaminergic system as the mechanism underlying personality in a beetle. J Insect Physiol 58:750–755
Nokelainen O, Hegna RH, Reudler JH, Lindstedt C, Mappes J (2012) Trade-off between warning signal efficacy and mating success in the wood tiger moth. Proc R Soc B 279:257–265
Ohno T, Miyatake T (2007) Drop or fly? Negative genetic correlation between death-feigning intensity and flying ability as alternative anti-predator strategies. Proc R Soc B 274:555–560
Parker GA (1970) Sperm competition and its evolutionary consequences in the insects. Biol Rev 45:525–567
Parker GA (1978) Evolution of competitive mate searching. Ann Rev Entomol 23:173–196
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ruxton GD, Allen WL, Sherratt TN, Speed MP (2018) Avoiding attack: the evolutionary ecology of crypsis, warning signals, and mimicry, 2nd edn. Oxford University Press, Oxford
SAS Institute (2015) JMP 12.2.0. SAS Institute, Cary
Schlager G (1960) Sperm precedence in the fertilization of eggs in Tribolium castaneum. Ann Entomol Soc Am 53:557–560
Sharma MD, Hunt J, Hosken DJ (2012) Antagonistic responses to natural and sexual selection and the sex-specific evolution of cuticular hydrocarbons in Drosophila simulans. Evolution 66:665–677
Sih A (1992) Prey uncertainty and the balancing of antipredator and feeding needs. Am Nat 139:1052–1069
Simmons LW (2001) Sperm competition and its evolutionary consequences in the insects. Princeton University Press, Princeton
Sokoloff A, Slatis HM, Stanley J (1960) The black mutation in Tribolium castaneum. J Hered 51:131–135
Suzuki T (1980) 4,8-Dimethyldecanal: the aggregation pheromone of the flour beetle, Tribolium castaneum and T. confusum (Coleoptera: Tenebrionidae). Agric Biol Chem 44:2519–2520
Thornhill R, Alcock J (1983) The evolution of insect mating systems. Harvard University Press, Cambridge
Wilber DH (1989) The influence of sexual selection and predation on the mating and postcopulatory guarding behavior of stone crabs (Xanthidae, Menippe). Behav Ecol Sociobiol 24:445–451
Zuk M, Simmons LW (1997) Reproductive strategies of the crickets (Orthoptera: Gryllidae). In: Crespi BJ (ed) The evolution of mating strategies in insects and arachnids. Cambridge University Press, Cambridge, pp 89–109
Acknowledgement
This work was supported by a Grant-in-Aid for Scientific Research (KAKENHI 26291091 and 18H02510, Ministry of Education, Culture, Sports, Science and Technology, Ministry of Education, Culture, Sports, Science and Technology) to T. M.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
K. Matsumura and T. Miyatake declare that they have no conflict of interest.
Ethical approval
All applicable international, national, and institutional guidelines for the care and use for animals were followed.
Research involving human participants and/or animals
This article does not contain any studies with human participants performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Matsumura, K., Miyatake, T. Influence of artificial selection for duration of death feigning on pre- and post-copulatory traits in male Tribolium castaneum. J Ethol 37, 265–270 (2019). https://doi.org/10.1007/s10164-019-00596-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10164-019-00596-4