
Abstract
Recent studies indicate that supporting cells play important roles in inner ear development, function, and regeneration after injury, but the molecular mechanisms underlying these processes remain poorly understood. Inducible cell-specific gene recombination in supporting cells could be a powerful tool to study the roles of specific molecules in these cells. Here we tested the feasibility, effectiveness, and cell specificity of inducible Cre-mediated gene recombination in the postnatal inner ear using mice that express an inducible form of Cre (CreERT) under the transcriptional control of the proteolipid protein (PLP) promoter. We assessed the pattern of tamoxifen-induced gene recombination in the inner ear using the ROSA26-LacZ reporter line, in which the β-galactosidase gene is expressed only after Cre-mediated excision of a loxP-flanked stop cassette. Recombination was detected in cochlear inner phalangeal cells, supporting cells surrounding hair cells in vestibular maculae and cristae. Recombination also occurred in Schwann cells. We also found that this CreERT line can be used to increase and decrease the levels of expression of a trophic factor, brain-derived neurotrophic factor, specifically in supporting cells. These results show that PLP/CreERT mice are a powerful tool to dissect gene function in inner ear supporting cells.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cochlear and vestibular sensory epithelia contain two primary cell types, hair cells and supporting cells. Although the roles of hair cells and their associated sensory neurons are well established, the functions of supporting cells are now beginning to emerge. There is increasing evidence that these non-neuronal cells play important roles in the development, maintenance, and function of the inner ear. For example, supporting cells act as precursors for hair cells during embryogenesis and can later trans-differentiate into hair cells in the adult avian inner ear (Morest and Cotanche 2004; Ryals and Rubel 1988). While adult mammalian supporting cells do not normally generate hair cells after injury, some studies indicate that they can do it if they are induced to express specific genes, such as the transcription factor Math1 (Gubbels et al. 2008). Thus, manipulation of supporting cell gene expression could one day become a tool to treat some forms of hearing loss.
Supporting cells also appear to be important regulators of synaptic activity in the sensory epithelia. Specifically, supporting cells of the organ of Corti contribute to spontaneous activity of the auditory nerve prior to the onset of hearing (Tritsch et al. 2007). In the mature sensory epithelia, glutamate transporters expressed by supporting cells have been implicated in the regulation of neurotransmission at the hair cell-afferent fiber synapse (Glowatzki et al. 2006). There is also evidence indicating that supporting cells contribute to the long-term survival of type I spiral ganglion neurons (Stankovic et al. 2004; Sugawara et al. 2005). This promotion of neuronal survival appears to be mediated by supporting cell-derived neurotophic factors such as neurotrophin 3 (Stankovic et al. 2004).
Further understanding of the roles of supporting cells in the postnatal inner ear would be greatly facilitated by the use of genetically modified mice. However, since many of the genes of interest are critical to the development and function of other tissues and cell types, it would be crucial to find a way to temporally alter gene expression specifically in supporting cells. We decided to test if this could be achieved using transgenic mice expressing tamoxifen-inducible Cre recombinase. We identified a mouse line that was likely to express CreERT in supporting cells under the control of the proteolipid protein (PLP) promoter (Doerflinger et al. 2003). While the PLP gene encodes for proteolipid protein known to be expressed by myelin-producing cells of the nervous system (Schwann cells and oligodendrocytes; Fuss et al. 2000; Mallon et al. 2002; Wight et al. 1993), a recent study showed that the PLP promoter is active in supporting cells of the inner ear at embryonic stages (Morris et al. 2006). Here we show that the PLP promoter continues to be active in supporting cells of the cochlea and vestibular organs at postnatal ages. Using the ROSA26-LacZ reporter mouse line, we found that tamoxifen treatment of PLP/CreERT mice any time between birth and postnatal day 17 (P17) induces effective gene recombination in cochlear inner phalangeal cells, supporting cells surrounding hair cells in vestibular maculae and Schwann cells in the osseous spiral lamina. Furthermore, we also found that expression of the trophic factor brain-derived neurotrophic factor (BDNF) can either be knocked out or induced in the vestibular epithelia of mice at the same ages using this PLP/CreERT line. These results show that PLP/CreERT mice are a powerful tool to dissect gene function in inner ear supporting cells and glia.
Materials and methods
Animals
PLP/CreERT (Doerflinger et al. 2003), BDNFf/f (Rios et al. 2001), and ROSA26-LacZ (Soriano 1999) mice were obtained from the Jackson Laboratory. Previously characterized mice expressing eGFP under the control of the PLP gene promoter were used to study the pattern of PLP-promoter activity in the postnatal inner ear (Mallon et al. 2002). The BDNFstop mouse line (Chang et al. 2006) was provided by Dr. Rudolf Jaenisch, Whitehead Institute for Biomedical Research, Cambridge, MA, USA. Mice were maintained in the animal facility at Children’s Hospital Boston, and all procedures were carried out following protocols approved by the Children’s Hospital Animal Care and Use Committee. At least three animals (six ears) were evaluated for recombination. Age-matched, wild-type littermates were used as controls.
Visualization of GFP
Mice were anesthetized with 2.5% Avertin (0.2 ml/10 g body weight) and fixed by intracardial perfusion with 4% paraformaldehyde. Temporal bones were dissected, incubated in 30% sucrose, and frozen. Sections (10 μm) were cut by cryostat. For whole-mount images, cochleas, utricles, and cristae were dissected and immediately observed under the fluorescent microscope.
Tamoxifen injection
Tamoxifen (Sigma, St. Louis, MO, USA) was dissolved in corn oil (Sigma) at a concentration of 10 mg/ml at 55°C. A 30 G1/2 needle insulin syringe was used for intraperitoneal injections in pups. All different combinations of mice were injected for seven consecutive days, starting at postnatal day 0 (P0) P5 or P10 with tamoxifen at a dose of 33 mg/kg body weight or with the corn oil carrier alone.
LacZ (β-galactosidase) staining
Mice were anesthetized as described above and fixed by intracardial perfusion with 2% paraformaldehyde–0.2% glutaraldehyde in 0.1 M phosphate-buffered saline (PBS, pH 7.4). The temporal bones were dissected and perfused by flushing the fixative solution through the oval window. The temporal bones were then washed three times for 30 min at room temperature and incubated in staining solution for 6 h at 37°C in the dark. The staining solution contained 5 mM K3Fe(CN)6, 5 mM K4Fe(CN)6.3 H2O, 2 mM MgCl2.6 H2O, 0.01% Na-deoxycholate, 0.02% NP-40, and 1 mg/ml X-gal. Tissues were rinsed three times with PBS for 5 min, postfixed overnight with the same fixing solution as above, washed with PBS, and decalcified for 2–3 days in 4% EDTA. Then, the tissues were embedded in araldite by using a rapid dehydration protocol to minimize washout of reaction product. No differences in the intensity or pattern of lacZ staining were observed before and after dehydration. Araldite-embedded materials were sectioned at 20 μm on a microtome (Leica). Ears of PLP/CreERT::ROSA26-LacZ double-transgenic mice injected with vehicle alone were processed in parallel as control.
Immunohistochemistry
Mice were anesthetized with 2.5% Avertin (0.2 ml/10 g body weight). The temporal bones were dissected, and the utricular maculae were removed and fixed in methanol at 4°C for 20 min as described in Montcouquiol et al. (1998). Tissues were washed in phosphate buffer and blocked for 2 h in 10% normal goat serum, 0.1% Triton-X, and 0.2% Tween. Chicken anti-BDNF (1:50, Promega, Madison, WI, USA) was incubated overnight at 4°C. Tissues were then washed in phosphate buffer and followed by overnight incubation with goat anti-chicken Alexa-488 (1:300, Molecular Probes, Eugene, OR, USA) at 4°C.
Real-time quantitative RT-PCR
Real-time quantitative RT-PCR was performed as in Stankovic and Corfas (2003). Briefly, P21 mice were killed, inner ears were extracted, cochlear and vestibular organs were dissected from otic capsule in RNAlater (Ambion, Austin, TX, USA), and total RNA was purified using RNeasy spin-columns (Qiagen, Valencia, CA, USA). Two inner ears from a single animal were processed as one sample. Only RNA samples that fulfilled the criteria that were established previously (Stankovic and Corfas 2003) were used for cDNA synthesis using iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA). Samples without reverse transcriptase were processed in parallel and served as negative controls. Measurements were made with an iCycler machine (Bio-Rad) and IQ SYBR Green supermix kit (Bio-Rad). For each well of the 96-well plate (Bio-Rad), the 25 μl reaction contained 12.5 μl of 2X IQ SYBR Green supermix, 0.5 μM each forward and reverse primer, 7.5 μl of RNase-free H2O, and 2.5 μl cDNA template. The cycling conditions were as follows: 95°C for 15 min followed by 50 cycles of 94°C for 30 s, 54°C for 30 s, and 72°C for 60 s. Each sample was loaded in triplicate. The following forward (F) and reverse (R) primers were used: for 18S rRNA (gene accession number X00686), F: CGGCTACCACATCCAAGGAA, R: GCTGGAATTACCGCGGCT (Hellstrom et al. 2001), which generates a 187-bp amplicon. For BDNF (gene accession number NM_007540), F: GTGTGTGACAGTATTAGCGAGTGG, R: GATACCGGGACTTTCTCTAGGAC, which generates a 101-bp amplicon.
Results
Pattern of PLP-promoter activity in the postnatal inner ear
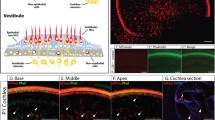
A recent study showed that the PLP promoter drives eGFP expression in supporting cells of the embryonic cochlea (Morris et al. 2006). Since other cells that express PLP during development continue to express relatively high levels of this molecule in the adult, e.g., oligodendrocytes (Mallon et al. 2002), we hypothesized that the same would be true for inner ear supporting cells. To test this possibility and to expand the analysis of PLP-promoter activity to the vestibular system, we examined the pattern of eGFP expression in the inner ear of PLP-eGFP mice at different postnatal ages (P3, P5, P15, and P21). Since results were similar at all ages, only P15 images are shown. In the organ of Corti, the only supporting cells that are positive for eGFP are those that surround the IHCs, i.e., the inner phalangeal cells, their fluorescence being intense without any obvious apex-to-base gradient. Schwann cells in the osseous spiral lamina also express eGFP (Fig. 1A, B). In the utricle and saccule, most supporting cells abutting hair cells also express eGFP, although the intensity of labeling varies among them (Fig. 1C, D). Similar to the organ of Corti, vestibular Schwann cells are also eGFP positive (Fig. 1D). In contrast to the maculae, not all supporting cells express eGFP in the cristae ampullaris (Fig. 1E, F). Rather, eGFP expression is restricted to supporting cells located in the base of the cristae.

PLP promoter activity in the postnatal inner ear. Bird’s eye view of whole mounts (A, C, and E) and images of cross sections (B, D, and F) of organ of Corti (A, B), utricular macula (C, D), and cristae ampullaris (E, F) from P15 PLP-eGFP mice (top imaging of eGFP, bottom imaging of eGFP (green) and nuclear staining (blue)). The images show that the PLP promoter drives transgene expression in supporting cells (SCs) that surround inner hair cells (IHCs) and Schwann cells in the cochlea (A, B), supporting cells abutting hair cells (HCs) in the vestibular maculae (C, D) and in some supporting cells in the cristae (E, F). Vestibular Schwann cells also express eGFP (D, E, and F). OHC outer hair cell, IPCs inner phalangeal cells. Scale bars = 100 μm (A, C, and E) and 25 μm (B, D, and F).
Using a reporter line to characterize the pattern of tamoxifen-induced recombination in the inner ear of postnatal PLP/CreERT mice
To directly test the ability of the PLP promoter to drive inducible gene recombination in postnatal inner ear supporting cells, we obtained a transgenic line in which the tamoxifen-inducible Cre (CreERT) is under the control of the PLP promoter (Doerflinger et al. 2003) and crossed it with the ROSA26-LacZ reporter mouse line in which the LacZ gene encoding β-galactosidase (β-gal) is expressed only after the Cre-mediated excision of a loxP-flanked stop cassette (Soriano 1999). PLP/CreERT::ROSA26-lacZ mice were injected intraperitoneally with either vehicle or tamoxifen at a dose of 33 mg/kg body weight daily for seven consecutive days beginning at P10. Temporal bones were collected 1 day after the last injection (P17), processed for β-gal histochemistry, and embedded in plastic. Examination of temporal bone whole mounts showed that tamoxifen induced effective recombination in the inner ear of PLP/CreERT::ROSA26-LacZ double-transgenic mice (Fig. 2A). LacZ staining was intense in the sensory epithelia of the cochlea without any obvious apex-to-base gradient. Strong staining was also evident in the sensory epithelia of the vestibular organs. Importantly, no staining was present when double-transgenic mice were injected with vehicle alone, indicating that recombination occurred only upon tamoxifen injection (Fig. 2B). These results show that tamoxifen can efficiently cross the barrier that exists between the blood circulation of the inner ear and the endolymph surrounding hair cells to effectively induce CreERT activity in the inner ear.

Visualization of PLP/CreERT-induced recombination using ROSA26-LacZ reporter mouse. A β-Galactosidase histochemistry in a temporal bone whole-mount from a PLP/CreERT::ROSA26-LacZ double-transgenic mouse injected with tamoxifen from P10 to P16 and harvested at P17. B Whole-mount staining from a PLP/CreERT::ROSA26-LacZ double-transgenic mouse injected with vehicle alone. Key structures are identified in A. Cochlear apex and base are indicated. Vestibular organs are labeled, including the cristae of the three semicircular canals (posterior [P], lateral [L], and superior [S]) and the maculae of the utricle and saccule. Scale bar = 0.5 mm.
To investigate the cell types in which gene recombination occurred—i.e., to identify the cells expressing the β-gal—the plastic-embedded whole mounts were sectioned and analyzed at higher magnification using established morphological criteria to identify the different cell types (Pannese 1981; Rio et al. 2002). In the organ of Corti, specific β-gal activity was detected in inner phalangeal cells adjacent to IHCs along the entire length of the cochlea (Fig. 3A, B). No signal was detected in hair cells or other supporting cell populations of the inner or outer hair cell areas. β-Gal was also expressed in Schwann cells in the osseous spiral lamina. In the spiral ganglion, no signal was apparent in the sensory neurons nor in the satellite cells that surround them (Pannese 1981; Fig. 3A, B, D, E). Double-transgenic mice injected with vehicle alone did not show evidence of recombination (Fig. 3C, F).

PLP/CreERT-induced gene recombination in the cochlea. β-Galactosidase (β-gal) histochemistry as seen in plastic sections (20 μm) through the organ of Corti (A, B) and spiral ganglion (D, E) from a P17 PLP/CreERT::ROSA26-LacZ double-transgenic mouse injected with tamoxifen from P10 to P16. Images were taken from the basal turn, corresponding to a frequency of roughly 25 kHz (Sugawara et al. 2005). C and F show that β-gal staining is absent from PLP/CreERT::ROSA26-LacZ double-transgenic mouse injected with vehicle alone. IHC inner hair cell, OHCs outer hair cells, IPCs inner phalangeal cells. Scale bar = 25 μm in A, C, D, and F; 50 μm in B and E.
Tamoxifen-induced Cre activity was also present in the vestibular end organs. In the utricular maculae, recombination was robust in most supporting cells (Fig. 4A). Also, in the saccular maculae we observed recombination in most supporting cells surrounding hair cells (not shown). In contrast, recombination in the cristae ampullaris was less reliable, occurring only in a fraction of the supporting cells (Fig. 4B), and in some cases β-gal activity was undetectable in these organs (not shown). As in the cochlea, recombination was detected in vestibular Schwann cells (Fig. 4A, B) and was absent from vestibular sensory neurons (Fig. 4C). However, unlike the cochlea, recombination was strong in vestibular satellite cells (Fig. 4C). No β-gal activity was detected in the vestibular system of PLP/CreERT::ROSA26-LacZ double-transgenic mice injected with vehicle alone (Fig. 4D–F). To study the pattern of tamoxifen-induced recombination at early time points, PLP/CreERT::ROSA26-lacZ mice were injected daily for seven consecutive days beginning at P0 and P3. Temporal bones were collected 1 day after the last injection and processed for β-gal histochemistry as described above. We observed that gene recombination occurred in the same cell types as mice that were injected beginning at P10 (data not shown).

PLP/CreERT-induced gene recombination in the vestibular organs. β-Galactosidase histochemistry as seen in plastic sections (20 μm) of the utricle (A), ampulla of the posterior canal (B), and vestibular ganglia (C) from a P17 PLP/CreERT::ROSA26-LacZ double-transgenic mouse injected with tamoxifen from P10 to P16. D, E, and F show that β-gal staining is absent from the utricle (D), ampulla of the posterior canal (E), and vestibular ganglia (F) from PLP/CreERT::ROSA26-LacZ double-transgenic mouse at P17 injected with vehicle alone. HC hair cell, SC supporting cell. Scale bar = 25 μm.
Tamoxifen induces effective recombination of the BDNF locus in the inner ear of postnatal PLP/CreERT mice
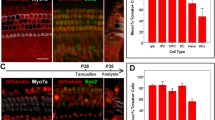
The evidence provided by the reporter mice indicated that the PLP/CreERT mice could be used to increase or decrease the expression of molecules of interest in supporting cells and some glia in the postnatal inner ear. We decided to test this by focusing on the neurotrophin BDNF, a molecule that has been shown to play important roles in the inner ear during embryonic stages (Ernfors et al. 1995; Ernfors et al. 1994) but whose function or expression pattern in the postnatal ear has not been defined. To test our ability to knockdown BDNF expression we crossed PLP/CreERT mice with mice in which the BDNF gene has been flanked by loxP sites (BDNFf/f; Rios et al. 2001). PLP/CreERT::BDNFf/f and BDNFf/f mice were injected with tamoxifen for seven consecutive days (P5–P11). When the animals reached P21, inner ears were dissected and analyzed. First, genomic DNA was purified and subjected to PCR with primers to test for recombination in the BDNF locus. Optic nerves were processed in parallel as positive controls since the nerve is primarily comprised of PLP-expressing oligodendrocytes. As shown in Figure 5A, recombination of the BDNF locus in the optic nerve was almost complete. In this tissue, the unrecombined BDNF locus most probably represents astrocytes, endothelial and connective tissue cells, which do not express PLP. In the inner ear, the level of recombination was significant, although lower than in the optic nerve, consistent with the lower percentage of PLP-expressing cells. No recombination was observed in animals that did not carry the PLP/CreERT transgene. Consistent with the apparent recombination of the BDNF gene, quantitative real-time PCR of inner ear extracts showed a dramatic reduction in the expression of BDNF in PLP/CreERT::BDNFf/f mice after tamoxifen injection (Fig. 5B). Similarly, semi-quantitative immunostaining of BDNF in P21 utricular maculae showed that the levels of BDNF protein were lower in tissues from PLP/CreERT::BDNFf/f mice after tamoxifen injection (Fig. 5C). In both cases, mRNA and protein, there was some residual BDNF signal in the vestibular system of PLP/CreERT::BDNFf/f mice, most likely a reflection of incomplete knockdown. The immunostaining also showed that at this age BDNF expression is uniformly restricted to vestibular supporting cells and absent from hair cells, as shown previously by (Montcouquiol et al. 1998). Remarkably, the inner ears of mice in which BDNF was knocked down after P5 appeared normal under gross histological analysis, i.e., in size and general cellular organization (data not shown).

The PLP/CreERT transgene allows for conditional BDNF knockdown in the postnatal inner ear. A Tamoxifen induces recombination of the BDNF gene in the inner ear of PLP/CreERT:: BDNFf/f mice. PCR of inner ear and optic nerve genomic DNA extracts with primers that produce different bands for the unrecombined (1 kb) and recombined (0.25 kb) BDNF alleles. No recombination is observed in animals that do not carry the PLP/CreERT transgene. B Real-time quantitative RT-PCR of whole inner ear RNA shows that tamoxifen induces knockdown of BDNF expression in the inner ear of mice carrying the PLP/CreERT transgene and two copies of the BDNFf allele (BDNFf/f). C Immunostaining with BDNF antibodies of the utricular maculae shows that tamoxifen induces a reduction of BDNF expression in supporting cells of mice carrying the PLP/CreERT transgene and two copies of the BDNFf allele (BDNFf/f).
To test if the levels of BDNF expression in supporting cells and Schwann cells can be increased in the postnatal inner ear we used a mouse line with an inducible BDNF transgene (BDNFstop; Chang et al. 2006). BDNFstop::PLP/CreERT and PLP/CreERT mice were injected with tamoxifen for seven consecutive days starting at P5, and BDNF expression levels were examined at P21. Quantitative real-time PCR of inner ears showed that tamoxifen induced a 2.5-fold increase in BDNF expression in the double-transgenic mice (Fig. 6A). Semi-quantitative analysis of BDNF immunostaining in the utricular maculae indicated that the levels of BDNF protein were higher in tissues from mice that carry the PLP/CreERT transgene in addition to the BDNFstop transgene (Fig. 6B).

The PLP/CreERT transgene allows for conditional BDNF over-expression in postnatal supporting cells. Immunostaining with BDNF antibodies (A) and quantitative real-time RT-PCR of whole inner ear RNA (B) show that tamoxifen increases BDNF expression in the utricular macula of mice carrying an inducible BDNF transgene (BDNFstop) when mice carry the PLP/CreERT transgene.
Discussion
Our results show that the PLP/CreERT mouse line represents a powerful tool to test the roles of supporting cells in the auditory and vestibular systems and the molecules involved in those processes. This mouse line will enable the manipulation of different classes of molecules in supporting cells, including trophic/growth factors or their receptors, transcription factors, cytoskeletal elements, and molecules involved in neurotransmitter function. Similarly, conditional manipulation of transcription factors involved in the genesis of hair cells by supporting cells could provide tests for the potential of hair cell regeneration in the adult (Gubbels et al. 2008). This Cre line, together with others that allow for inducible gene recombination in hair cells (Chow et al. 2006), provide unique opportunities to explore the cellular and molecular mechanisms that govern the inner ear and the pathogenesis of sensorineural hearing and balance disorders.
Our findings also provide another example of the analogies between supporting cells of the inner ear and glial cells. It is becoming clear that supporting cells have many similarities to glial cells, including the expression of numerous proteins, i.e., vimentin (Anniko et al. 1986), S100β (Pack and Slepecky 1995), glutamate-aspartate transporter (Furness and Lehre 1997; Glowatzki et al. 2006), low-affinity neurotrophin receptor p75 (Vega et al. 1999), the glial fibrillary acidic protein (GFAP; Rio et al. 2002), and the PLP (Morris et al. 2006). In addition, our present work has expanded on the results of Morris et al. (2006), indicating that the PLP promoter is not only active in embryonic supporting cells but it also continues to be active in mature supporting cells. Interestingly, our analysis shows that PLP is not expressed by all supporting cells, i.e., in the cochlea is only expressed by supporting cells surrounding IHCs. It is possible that other CreERT lines such as those using the GFAP promoter (Hirrlinger et al. 2006) may serve as drivers to other supporting cells, in which we have shown this promoter to be active (Rio et al. 2002).
References
Anniko M, Thornell LE, Gustavsson H, Virtanen I (1986) Intermediate filaments in the newborn inner ear of the mouse. ORL J Otorhinolaryngol Relat Spec 48(2):98–106
Chang Q, Khare G, Dani V, Nelson S, Jaenisch R (2006) The disease progression of Mecp2 mutant mice is affected by the level of BDNF expression. Neuron 49(3):341–348
Chow LM, Tian Y, Weber T, Corbett M, Zuo J, Baker SJ (2006) Inducible Cre recombinase activity in mouse cerebellar granule cell precursors and inner ear hair cells. Dev Dyn 235(11):2991–2998
Doerflinger NH, Macklin WB, Popko B (2003) Inducible site-specific recombination in myelinating cells. Genesis 35(1):63–72
Ernfors P, Lee KF, Jaenisch R (1994) Mice lacking brain-derived neurotrophic factor develop with sensory deficits. Nature 368(6467):147–150
Ernfors P, Kucera J, Lee KF, Loring J, Jaenisch R (1995) Studies on the physiological role of brain-derived neurotrophic factor and neurotrophin-3 in knockout mice. Int J Dev Biol 39(5):799–807
Furness DN, Lehre KP (1997) Immunocytochemical localization of a high-affinity glutamate-aspartate transporter, GLAST, in the rat and guinea-pig cochlea. Eur J Neurosci 9(9):1961–1969
Fuss B, Mallon B, Phan T, Ohlemeyer C, Kirchhoff F, Nishiyama A, Macklin WB (2000) Purification and analysis of in vivo-differentiated oligodendrocytes expressing the green fluorescent protein. Dev Biol 218(2):259–274
Glowatzki E, Cheng N, Hiel H, Yi E, Tanaka K, Ellis-Davies GC, Rothstein JD, Bergles DE (2006) The glutamate-aspartate transporter GLAST mediates glutamate uptake at inner hair cell afferent synapses in the mammalian cochlea. J Neurosci 26(29):7659–7664
Gubbels SP, Woessner DW, Mitchell JC, Ricci AJ, Brigande JV (2008) Functional auditory hair cells produced in the mammalian cochlea by in utero gene transfer. Nature 455(7212):537–541
Hellstrom A, Perruzzi C, Ju M, Engstrom E, Hard AL, Liu JL, Albertsson-Wikland K, Carlsson B, Niklasson A, Sjodell L, LeRoith D, Senger DR, Smith LE (2001) Low IGF-I suppresses VEGF-survival signaling in retinal endothelial cells: direct correlation with clinical retinopathy of prematurity. Proc Natl Acad Sci USA 98(10):5804–5808
Hirrlinger PG, Scheller A, Braun C, Hirrlinger J, Kirchhoff F (2006) Temporal control of gene recombination in astrocytes by transgenic expression of the tamoxifen-inducible DNA recombinase variant CreERT2. Glia 54(1):11–20
Mallon BS, Shick HE, Kidd GJ, Macklin WB (2002) Proteolipid promoter activity distinguishes two populations of NG2-positive cells throughout neonatal cortical development. J Neurosci 22(3):876–885
Montcouquiol M, Valat J, Travo C, Sans A (1998) A role for BDNF in early postnatal rat vestibular epithelia maturation: implication of supporting cells. Eur J Neurosci 10(2):598–606
Morest DK, Cotanche DA (2004) Regeneration of the inner ear as a model of neural plasticity. J Neurosci Res 78(4):455–460
Morris JK, Maklad A, Hansen LA, Feng F, Sorensen C, Lee KF, Macklin WB, Fritzsch B (2006) A disorganized innervation of the inner ear persists in the absence of ErbB2. Brain Res 1091(1):186–199
Pack AK, Slepecky NB (1995) Cytoskeletal and calcium-binding proteins in the mammalian organ of Corti: cell type-specific proteins displaying longitudinal and radial gradients. Hear Res 91(1–2):119–135
Pannese E (1981) The satellite cells of the sensory ganglia. Adv Anat Embryol Cell Biol 65:1–111
Rio C, Dikkes P, Liberman MC, Corfas G (2002) Glial fibrillary acidic protein expression and promoter activity in the inner ear of developing and adult mice. J Comp Neurol 442(2):156–162
Rios M, Fan G, Fekete C, Kelly J, Bates B, Kuehn R, Lechan RM, Jaenish R (2001) Conditional deletion of brain-derived neurotrophin factor in the postnatal brain leads to obesity and hyperactivity. Mol Endocrinol 15:1748–1757
Ryals BM, Rubel EW (1988) Hair cell regeneration after acoustic trauma in adult Coturnix quail. Science 240(4860):1774–1776
Soriano P (1999) Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat Genet 21(1):70–71
Stankovic KM, Corfas G (2003) Real-time quantitative RT-PCR for low-abundance transcripts in the inner ear: analysis of neurotrophic factor expression. Hear Res 185(1–2):97–108
Stankovic K, Rio C, Xia A, Sugawara M, Adams JC, Liberman MC, Corfas G (2004) Survival of adult spiral ganglion neurons requires erbB receptor signaling in the inner ear. J Neurosci 24(40):8651–8661
Sugawara M, Corfas G, Liberman MC (2005) Influence of supporting cells on neuronal degeneration after hair cell loss. J Assoc Res Otolaryngol 6(2):136–147
Tritsch NX, Yi E, Gale JE, Glowatzki E, Bergles DE (2007) The origin of spontaneous activity in the developing auditory system. Nature 450(7166):50–55
Vega JA, San Jose I, Cabo R, Rodriguez S, Represa J (1999) Trks and p75 genes are differentially expressed in the inner ear of human embryos. What may Trks and p75 null mutant mice suggest on human development? Neurosci Lett 272(2):103–106
Wight PA, Duchala CS, Readhead C, Macklin WB (1993) A myelin proteolipid protein-LacZ fusion protein is developmentally regulated and targeted to the myelin membrane in transgenic mice. J Cell Biol 123(2):443–454
Acknowledgments
We thank Dr. Rudolph Jaenisch for the generous gift of the BDNFstop transgenic line; Drs. Lihong Bu, Marcin Liana, Sylvia Chen, and Samir Koirala for their technical help; and Roman Corfas for his comments on the manuscript. This research was supported in part by grants from the NIDCD: R01 DC 004820 and P30 DC 005209 (to G.C.) and a Pew Latin American Fellow (to M.E.G.C).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gómez-Casati, M.E., Murtie, J., Taylor, B. et al. Cell-Specific Inducible Gene Recombination in Postnatal Inner Ear Supporting Cells and Glia. JARO 11, 19–26 (2010). https://doi.org/10.1007/s10162-009-0191-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10162-009-0191-x