
Abstract
Population dynamics of terrestrial vertebrates are affected by climatic fluctuations, notably in ectotherms. An understanding of the interaction between physiology and demographic processes is necessary to predict the impacts of climate change. Reptiles are particularly sensitive to temperature, but only a few studies have explored the relationship between thermoregulatory strategy and demography in these animals. Using 12 years of mark-recapture data on two sympatric colubrid snakes (Hierophis viridiflavus and Zamenis longissimus), we tested whether demographic parameters are influenced by contrasted thermoregulatory strategies. The thermophilic and conspicuous species (H. viridiflavus) grew faster than the thermoconforming and secretive species (Z. longissimus), and this difference was most pronounced in open habitats, suggesting that the metabolic benefits associated with high thermal preferences depend on environmental factors at small spatial scales. Survival varied annually in both species, but was not lower in H. viridiflavus despite a higher degree of exposure. In Z. longissimus, survival was negatively affected by low temperatures during the active season, possibly underlying an exposure trade-off.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Climatic conditions affect population dynamics by influencing a variety of ecological processes (Stenseth et al. 2002). The effects of seasonality, El Niño–Southern Oscillation (ENSO), and global warming on demography are well documented in a broad range of species (Jaksic et al. 1997; Sæther et al. 2000; Lima et al. 2003; McDonald et al. 2004). Over the past decades, most studies of demographic responses to climate have focused on endothermic mammals and birds. This is surprising, however, because ectotherms are particularly sensitive to climatic variations (Madsen and Shine 2000; Lourdais et al. 2004; Chamaillé-Jammes et al. 2006; Marquis et al. 2008; Le Galliard et al. 2010; Sinervo et al. 2010). Indeed, in ectotherms body temperature depends on environmental temperature and influences physiological performance (Huey and Stevenson 1979), metabolism (Secor and Nagy 1994; Dubois et al. 2008), locomotion (Bennett 1980; Stevenson et al. 1985; Ojanguren and Braña 2000), immune function (Mondal and Rai 2001), foraging ability (Ayers and Shine 1997; Carrière and Boivin 1997), and growth (Kingsolver and Woods 1997). Therefore, we should expect a particularly strong effect of climate on demography in ectotherms (Kearney and Porter 2004; Helmuth et al. 2005; Deutsch et al. 2008; Le Galliard et al. 2012; Weatherhead et al. 2012).
In terrestrial reptiles, species subjected to similar environmental constraints nonetheless can diverge in thermoregulatory strategy (Belliure et al. 1996; Scheers and Van Damme 2002; Du et al. 2006). Under cool climates, thermoregulation primarily involves direct exposure to solar radiation (Huey and Slatkin 1976; Blouin-Demers and Nadeau 2005). Because basking behaviour is associated with increased exposure in open habitats, it may entail significant survival costs (Webb and Whiting 2005). On the other hand, a thermophilic species should show enhanced performance and obtain fitness benefits through faster growth and earlier sexual maturity. Because of coadaptation between traits (Angilletta et al. 2006), species with distinct thermoregulatory strategies can exhibit divergent energy budgets (Secor and Nagy 1994), foraging modes (Belliure et al. 1996), habitat use (Du et al. 2006; Carfagno and Weatherhead 2008), and degrees of exposure (Daly et al. 2008). Therefore, we should expect divergent thermoregulatory strategies to be associated with different demographic characteristics (growth, survival) and sensitivity to climatic conditions. Yet, this issue has heretofore not been investigated.
Detailed information on the effect of thermal ecology on demography is not currently available for snakes; yet lizard studies (Sceloporus sp.) revealed complex links between thermal environment, population dynamics, and many life history traits (Angilletta et al. 2002). Although conducting long-term demographic studies on a suite of sympatric and syntopic species differing in their thermal ecologies would afford the most inferential power, this may not prove logistically feasible. Therefore, species pair comparisons provide an initial step. To be meaningful, a comparison of two species must rely on the analysis of a set of life history traits that allow a priori predictions on the direction of the trait considered (Garland and Adolph 1994). For instance, two species comparisons have been used successfully in sympatric snakes with contrasted foraging modes (a sit-and-wait and an active forager) to compare activity, growth, maturity, survival, and energy budgets (Secor and Nagy 1994; Webb et al. 2003).
We studied two sympatric colubrid snakes, the European whip snake Hierophis viridiflavus (Lacépède, 1789; HV) and the Aesculapian snake Zamenis longissimus (Laurenti, 1768; ZL) to examine the consequences of thermoregulatory strategy on demography while minimizing the limitations inherent to two-species comparisons (Garland and Adolph 1994). Both species are morphologically and ecologically similar for several major traits (body size, diet, habitat, reproductive mode, and activity periods; Naulleau 1984; Lelièvre et al. 2012), and are also largely sympatric and syntopic (Luiselli and Capizzi 1997; Naulleau 1997a, b). However, they exhibit contrasted exposure strategies, thermal specialization and energy requirements (Lelièvre et al. 2010, 2011). HV is fast, active, and performs optimally at high temperatures whereas ZL is elusive and prefers cooler temperatures. The two species are so similar in aspects other than their thermal ecologies (see “Methods”) that we can confidently rule out many factors (e.g., food availability) as the source of variation in demography. We used mark-recapture data gathered over 12 years on syntopic populations of HV and ZL in Western France. We tested the hypothesis that the demography of ectotherms depends on their thermoregulatory and exposure strategies. More specifically, that species preferring high body temperatures should be characterized by fast life histories. We tested the following set of predictions:
-
1.
Since physiological performances depend on body temperature, the thermophilic HV should grow faster than the thermoconforming ZL, particularly as juveniles because adults invest energy in reproduction;
-
2.
Growth should be more dependent on environmental conditions (e.g., basking site availability) in HV because of higher energy requirements;
-
3.
Because high thermal requirements imply basking, and thus higher risks of predation, HV should exhibit lower survival.
Methods
Study species and study site
Hierophis viridiflavus and Zamenis longissimus are medio-European sympatric colubrid snakes that frequently occur in syntopy (Naulleau 1997a, b; Rugiero et al. 2002). Both species have approximately equal body size (~130–150 cm) and are active foragers showing important overlap in their diets (mostly composed of small mammals; Naulleau 1984; Capizzi et al. 1995; Lelièvre et al. 2012). On the other hand, they diverge in thermoregulatory and exposure strategies. HV is a typical racer (Luiselli 2006) characterized by fast moving ability, high levels of activity and exposure, and elevated preferred body temperatures (Tset 27.5–31.0 °C; Lelièvre et al. 2010). Conversely, ZL is an elusive ratsnake (similar to Elaphe, Pantherophis, or Ptyas genera; Lenk et al. 2001), using closed microhabitats (Lelièvre et al. 2011), and selecting cooler temperatures (Tset 21.5–25.5 °C; Lelièvre et al. 2010). We have previously shown that both species differ in optimal temperatures for physiological performance and in the shape of their performance curves (Lelièvre et al. 2010). Energy requirements reflect contrasted thermal preference and are higher in the racer (Lelièvre et al. 2010).
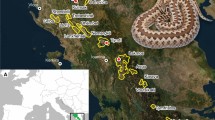
Population monitoring was conducted at the Centre d’Études Biologiques de Chizé in Western France (46°07′N; 00°25′W; Fig. 1). The study site is a 2600-ha biological reserve managed by the Office National des Forêts. The forest is dominated by deciduous trees (Fagus, Quercus, Carpinus, Acer), regeneration areas characterized by scrub species (Rubus, Clematis), and grasslands. Climatic conditions are temperate oceanic with annual precipitations between 800 and 1,000 mm, annual mean temperature of 12 °C, and an average of 2,000 h of sunshine per year.

Location of the four study sites at the Centre d’Études Biologiques de Chizé (Western France). Area 1 is grassland. Areas 2, 3, and 4 are forest. Dashed line indicates the border of the biological reserve
Capture-mark-recapture
Snakes were captured during the active season (April–September) under undulated concrete boards (120 × 80 cm) disposed in four areas of the study site for this purpose (Bonnet et al. 1999; Shine and Bonnet 2009; Fig. 1). Importantly, these four areas are distant enough that individuals almost never move between them. Data were gathered on 1,608 snakes between 1997 and 2009 (844 ZL; 764 HV; Table 1). We recorded snout–vent length (SVL: from the tip of the snout to the cloacal scale) to the nearest 0.5 cm, body mass to the nearest 1 g in adults and to the nearest 0.1 g in small snakes (less than 100 g), and sex by eversion of the hemipenes and from tail shape. Each snake was individually and permanently identified by burning ventral scales in unique combinations. Snakes were then released at their capture site as soon as possible.
Climatic data
Data were gathered from Météo-France station in Niort situated 20 km from the study site. Available variables were daily minimum and maximum temperatures, and daily precipitation between 1997 and 2009.
Growth
Growth in snakes is the increase in SVL over time. To limit the influence of measurement error, which can be substantial in snakes (Luiselli 2005), we restricted our analyses to long time intervals (>60 days) between captures. Negative values of growth were not removed because we assumed that positive measurement errors should be equally prevalent, but not identifiable (Blouin-Demers et al. 2002; Bonnet et al. 2002). As juvenile snakes show much lower capture rates than adults (Pike et al. 2008), we were unable to build robust Von Bertalanffy growth models. Instead, growth rate was expressed as the difference in body size (SVL in cm) between captures divided by the time elapsed in days. Overwintering (150 days per year) was deducted from growth intervals for captures not in the same active season because snakes are inactive and do not grow during this period. We used time between captures as a covariate in the model because it may influence error in growth rate if significant measurement error occurred. We also used SVL at first capture as a covariate because snake growth decreases with increasing body size (Blouin-Demers et al. 2002). In our study site, both species become sexually mature at a similar size (~70 cm SVL, X. Bonnet, personal observation). We used the same criterion for males and females to separate juveniles from adults. We compared juvenile growth rates in two habitats: grassland (Area 1 in Fig. 1) and forest (Areas 2, 3, and 4 in Fig. 1).
Survival
We estimated local annual survival and recapture probabilities from 1997 to 2009 using the Cormack–Jolly–Seber (CJS) model in program MARK 4.1 (White and Burnham 1999). Assumptions of capture-recapture modeling were tested by a χ² goodness-of-fit test (Pollock et al. 1990) using program U_CARE 2.2 (Choquet et al. 2009). The model adequately fitted the data (χ 2 = 68.73; df = 125; P = 0.99) as we found evidence for neither trap dependence (Test2.CT: P = 0.97) nor transience (Test3.SR: χ2 = 1.62, P = 0.10).
A set of candidate models with various parameterizations for survival rate (φ) and capture probabilities (p) (i.e., time-dependent (t), constant over time (.), group dependent (g), group being sex or species) were then developed and we used Akaike’s Information Criteria (AICc) to rank and select models (QAIC; Burnham and Anderson 2002). If models could not be classified from AICc (Δ AICc < 2), we performed model averaging (Buckland et al. 1997) using the AIC weight to recalculate an intermediate estimate (Burnham and Anderson 2002).
Model selection based on AICc showed that capture probability was time-dependant and differed slightly between the sexes in HV (average p = 47.1 %, 95 % CI: 24.6–68.8, in males vs. 33.1 %, 95 % CI: 15.0–56.7, in females) and ZL (average p = 48.3 %, 95 % CI: 28.8–76.6, in males vs. 25.8 %, 95 % CI: 16.3–59.8, in females). Because we captured snakes under shelter, capture probability cannot be used to estimate level of exposure.
We tested for the influence of climatic covariates (temperature, rainfall) on survival rates. For temperature, we used the mean of daily maxima during the snake activity season (April–September) and for rainfall we used the total precipitation during the same period (Fig. 2). We used the procedure ANODEV in MARK 4.1 to test whether the variance explained by these covariates was statistically significant (Harris et al. 2005). We assessed the fraction of temporal variation in survival explained by covariates using the following formula:

Climatic data for the study area in Western France used as covariates in survival models for Z. longissimus and H. viridiflavus from 1997 to 2009. Lines show mean daily maximum temperatures during snake activity season (April–September) and the histogram shows cumulated rainfall during the same period. Data were obtained from Niort meteorological station (Météo France) situated approximately 20 km from the study area
where φ(.) is the constant model, φ(t) is the time dependent model, and φ(cov) is the model with covariate.
All values are reported as mean ± SE.
Results
Growth
In both species, growth rates decreased with body size (regressions initial SVL-growth rate in HV: df = 1,402; R 2 = 0.33; P < 0.0001; in ZL: df = 1,284; R 2 = 0.19; P < 0.0001; Fig. 3). Growth rates of juvenile HV (N = 95) were significantly higher than those of ZL [N = 51; general linear model (GLM) with change in body size as the dependent variable, species and sex as factors, and initial SVL and time between recaptures as covariates; F 1,140 = 5.79; P = 0.02; with no significant interaction between species and sex F 1,140 = 0.47; P = 0.49; Fig. 4]. No interspecific difference in growth rate was detected in adults (N = 308 and 234; GLM; F 1,536 = 0.05; P = 0.83; interaction between species and sex F 1,536 = 0.78; P = 0.38; Fig. 4).

Individual growth rates obtained through mark recapture of H. viridiflavus (HV) and Z. longissimus (ZL) in the Centre d’Études Biologiques de Chizé, Western France, from 1997 to 2009 (white circles for HV and black circles for ZL). Curves (grey for HV and black for ZL) represent logistic regression fits

Comparison between growth rates of juvenile and adult snakes in H. viridiflavus (HV) and Z. longissimus (ZL) obtained through mark recapture in the Centre d’Études Biologiques de Chizé, Western France, from 1997 to 2009 (light grey bars for HV and dark grey bars for ZL). Bars represent SE and same letters indicate no significant difference. Sample sizes are in italic
Importantly, juveniles grew faster in grassland than in forest (N = 41 and 70; GLM; F 1,105 = 7.81; P = 0.002; Fig. 5) with an interaction between species and area (GLM; F 1,105 = 9.59; P = 0.003). The effect of habitat on growth was pronounced in juvenile HV [least significant difference (LSD) Fisher post hoc test; P < 0.0001), but absent in ZL (LSD Fisher post hoc test; P = 0.56).

Comparison between growth rates in juvenile H. viridiflavus (HV) and Z. longissimus (ZL) captured in different habitats obtained through mark recapture in the Centre d’Études Biologiques de Chizé, Western France, from 1997 to 2009 (light grey bars for HV and dark grey bars for ZL). Bars represent SE and same letters indicate no significant difference. Sample sizes are in italic
Survival
For HV, the most parsimonious survival model identified by AIC was φ(t) p(g + t) (Table 2). Survival was time-dependant and did not differ between the sexes (average φ = 54.7 %, 95 % CI: 36.8–82.7; Fig. 6). The model failed to estimate survival in 2000, 2004, and 2008 because of low captures. For ZL, three models were equally parsimonious, and we thus performed model averaging on survival parameters (Table 2). Averaged survival was time-dependant and did not differ between the sexes (average φ = 50.6 %, 95 % CI: 22.8–76.6). Survival rates did not differ between species. The best-supported two-species model (with species as group factor and pooled sexes) was φ(t) p(g + t), where survival was time-dependant and capture probability varied with time and species.

Annual variations in mean temperature (thick grey line) and adult survival of Z. longissimus (ZL) and H. viridiflavus (HV) obtained through mark recapture in the Centre d’Études Biologiques de Chizé, Western France, from 1997 to 2009 (light grey triangles for HV and dark grey squares for ZL). Both sexes are pooled (see text for model selection). Bars represent SE
Influence of climate on demography
Maximum temperature and precipitation during the snake activity season were strongly negatively correlated (R 2 = 0.99, P < 0.0001). Each parameter significantly affected adult survival in ZL when taken independently. Maximum temperatures explained 43.3 % of interannual variation in survival (ANODEV; F 1,8 = 6.10; P = 0.04) and precipitation explained 62.2 % (ANODEV; F 1,8 = 13.19; P = 0.007). Nevertheless, precipitation had no effect when added to temperature (ANODEV; F 1,7 = 1.35; P = 0.28), which is unsurprising given their very high correlation. Survival was highest in ZL when climatic conditions were hot and dry during the activity season (Figs. 2, 6). Conversely, none of the climatic variables tested explained variation in adult survival for HV (maximal temperature: ANODEV; F 1,7 = 0.39; P = 0.55; precipitation: ANODEV; F 1,7 = 2.06; P = 0.19).
Discussion
This long-term study allowed us to examine the hypothesis that the demography of snakes depends on their thermoregulatory strategy. As predicted, the thermophilic species exhibited faster juvenile growth. By selecting higher body temperatures, HV can enhance foraging performance and digestion speed (Lelièvre et al. 2010, 2011), and potentially better exploit prey populations. As both species mature at a similar length, the faster growth of HV should result in a lower age at maturity compared to ZL and, if prey are readily available, this could result in a higher population growth rate in HV compared to ZL.
Growth rates were also affected by habitat type in HV: HV exhibited faster growth in grasslands than in forest. This habitat difference in growth was possibly driven by divergent food availability since rodents, and notably common voles (Naulleau 1984), are more abundant in open fields compared to forest (Gauffre et al. 2008). Food availability is probably a key parameter affecting growth rate and reproductive output in these active predators. The difference in growth across habitats could also be related to enhanced physiological performances associated with open habitat use since open habitats allow to maintain higher body temperatures (Lelièvre et al. 2011, but see also Michel and Bonnet 2010). Because snakes were monitored in a single grassland site, however, we cannot tease apart site from habitat effects and further study is thus required.
The thermophilic HV did not exhibit lower survival, contrary to our prediction. Survival estimates were not available every year for HV, thereby reducing our ability to detect a difference. Alternatively, our initial hypothesis on survival might have been too simplistic. For instance, a high exposure thermal strategy may be associated with compensatory responses that minimize predation risk. Selection of high body temperatures enhances locomotor performance and fleeing ability (Arnold and Bennett 1984; Bauwens et al. 1995; Brodie and Russel 1999), notably in HV (Lelièvre et al. 2010). In addition, disruptive colouration in HV may help evade predators (Stevens et al. 2008). A comparative study conducted on two Australian nocturnal elapid snakes showed the same pattern: the two species exhibited similar adult survival rates despite contrasted foraging modes (active vs. sit-and-wait) and exposure strategies (Webb et al. 2003).
Climatic conditions during the activity season do not affect survival in HV, while ZL was sensitive to fluctuations in rainfall and temperature. The relatively low thermal requirements of ZL are related to a more secretive lifestyle (Lelièvre et al. 2011). When thermal conditions are hot, closed habitats such as scrub and forest provide favourable temperatures for ZL (Lelièvre et al. 2011). Conversely, closed habitats are usually too cool for HV, even in hot conditions. Therefore, colder conditions should be more detrimental to ZL because these conditions would impose the need for individuals to bask and thus potentially increase their risk of predation. Predation pressure is high in our study site because of the presence of raptors foraging on snakes: Circaetus gallicus and Buteo buteo (Bonnet et al. 1999; Gil and Pleguezuelos 2001; Selas 2001). Moreover, both ZL and HV exhibit extensive movements during the reproductive period (Bonnet et al. 1999). Given the poor crawling performance of ZL (Lelièvre et al. 2010), a higher sensitivity to avian predation is expected.
Overall, our empirical data allow a better understanding of the consequences of different thermal strategies on demography in ectotherms. Our results also underline the importance of considering thermal specialization as well as exposure strategies to elaborate realistic predictions on the potential effects of climate change on reptiles (Huey et al. 2009; Weatherhead et al. 2012). The impact of global warming on European reptiles attracted significant interest (Araujo et al. 2006; Le Galliard et al. 2012). In our study system, survival should be positively influenced by a mild warming in the elusive species while growth should be more affected in the exposed species because of increased thermoregulatory opportunities. The exact response will depend on the extent of warming, however, notably in the thermoconformer that displays lower thermal preferences and depends on the coolest habitats. In this species, deviant (high) temperature experienced during the day should have negative impacts such as elevated maintenance costs, altered digestive efficiency and a reduction in the active time window. However, these potential costs can be alleviated by a behavioural shift from diurnal to nocturnal activity (Sinervo et al. 2010; Weatherhead et al. 2012) as observed in southern populations (L. Luiselli, personal communication).
Thermoregulatory and exposure strategies should also influence the biogeographical response to climate change. Mobile species characterized by high activity and “risk prone” behavioural tactics should respond more dynamically and be better able to colonize newly favourable habitats. The distribution of HV has shown a clear progression at the northern margin of its range over the past 60 years (Naulleau 2003). Such observations provide evidence for a rapid response to global change (climate and habitat) in this species. Further studies conducted on sympatric reptiles that differ on their physiological and ecological traits would help evaluate the generality of these differences in the ability to respond to perturbations (Huey et al. 2009).
References
Angilletta MJ, Niewiarowski PH, Navas CA (2002) The evolution of thermal physiology in ectotherms. J Therm Biol 27:249–268
Angilletta MJ, Bennett AF, Guderley H, Navas CA, Seebacher F, Wilson RS (2006) Coadaptation: a unifying principle in evolutionary thermal biology. Physiol Biochem Zool 79:282–294
Araujo MB, Thuiller W, Pearson RG (2006) Climate warming and the decline of amphibians and reptiles in Europe. J Biogeogr 33:1712–1728
Arnold SJ, Bennett AF (1984) Behavioral variation in natural populations. III. Antipredator displays in the garter snake Thamnophis radix. Anim Behav 32:1108–1138
Ayers DY, Shine R (1997) Thermal influences on foraging ability: body size, posture and cooling rate of an ambush predator, the python Morelia spilota. Funct Ecol 11:342–347
Bauwens D, Garland T Jr, Castilla AM, Van Damme R (1995) Evolution of sprint speed in Lacertid lizards: morphological, physiological and behavioral covariation. Evolution 49:848–863
Belliure J, Carrascal LM, Diaz JA (1996) Covariation of thermal biology and foraging mode in two Mediterranean Lacertid lizards. Ecology 77:1163–1173
Bennett AF (1980) The thermal dependence of lizard behavior. Anim Behav 28:752–762
Blouin-Demers G, Nadeau P (2005) The cost-benefit model of thermoregulation does not predict lizard thermoregulatory behaviour. Ecology 86:560–566
Blouin-Demers G, Prior KA, Weatherhead PJ (2002) Comparative demography of black ratsnakes (Elaphe obsoleta) in Ontario and Maryland. J Zool 256:1–10
Bonnet X, Naulleau G, Shine R (1999) The dangers of leaving home: dispersal and mortality in snakes. Biol Conserv 89:39–50
Bonnet X, Pearson DJ, Ladyman M, Lourdais O, Bradshaw SD (2002) ‘Heaven’ for serpents? A mark-recapture study of tiger snakes (Notechis scutatus) on Carnac Island, Western Australia. Austral Ecol 27:442–450
Brodie ED, Russel NH (1999) The consistency of individual differences in behaviour: temperature effects on antipredators behaviour in garter snakes. Anim Behav 57:445–451
Buckland ST, Burnham KP, Augustin NH (1997) Model selection: an integral part of inference. Biometrics 53:603–618
Burnham KP, Anderson DR (2002) Model selection and multimodel inference. Springer, New York
Capizzi D, Luiselli L, Capula M, Rugiero L (1995) Feeding habits of a Mediterranean community of snakes in relation to prey availability. Rev Ecol-Terre Vie 50:353–363
Carfagno GLF, Weatherhead PJ (2008) Energetics and space use: intraspecific and interspecific comparisons of movements and home ranges of two Colubrid snakes. J Anim Ecol 77:416–424
Carrière Y, Boivin G (1997) Evolution of thermal sensitivity of parasitization capacity in egg parasitoids. Evolution 51:2028–2032
Chamaillé-Jammes S, Massot M, Aragon P, Clobert J (2006) Global warming and positive fitness response in mountain populations of common lizards Lacerta vivipara. Glob Change Biol 12:392–402
Choquet R, Lebreton JD, Gimenez O, Reboulet AM, Pradel R (2009) U-CARE: utilities for performing goodness of fit tests and manipulating capture-recapture data. Ecography 32:1071–1074
Daly BG, Dickman CR, Crowther MS (2008) Causes of habitat divergence in two species of agamid lizards in arid central Australia. Ecology 89:65–76
Deutsch CA, Tewksbury JJ, Huey RB, Sheldon KS, Ghalambor CK, Haak DC, Martin PR (2008) Impacts of climate warming on terrestrial ectotherms across latitude. Proc Natl Acad Sci USA 105:6668–6672
Du WG, Shou L, Shen JY (2006) Habitat selection in two sympatric Chinese skinks, Eumeces elegans and Sphenomorphus indicus: do thermal preferences matter? Can J Zool 84:1300–1306
Dubois Y, Blouin-Demers G, Thomas DW (2008) Temperature selection in wood turtles (Glyptemys insculpta) and its implications for energetics. Écoscience 15:398–406
Garland T Jr, Adolph SC (1994) Why not to do 2-species comparative studies—Limitations on inferring adaptation. Physiol Zool 67:797–828
Gauffre B, Estoup A, Bretagnolle V, Cosson JF (2008) Spatial genetic structure of a small rodent in a heterogeneous landscape. Mol Ecol 17:4619–4629
Gil JM, Pleguezuelos JM (2001) Prey and prey-size selection by the short-toed eagle (Circaetus gallicus) during the breeding season in Granada (south-eastern Spain). J Zool 255:131–137
Harris MP, Anker-Nilssen T, McCleery RH, Erikstad KE, Shaw DN, Grosbois V (2005) Effect of wintering area and climate on the survival of adult Atlantic puffins Fratercula arctica in the eastern Atlantic. Mar Ecol-Prog Ser 297:283–296
Helmuth B, Kingsolver JG, Carrington E (2005) Biophysics, physiological ecology, and climate change: does mechanism matter? Annu Rev Physiol 67:177–201
Huey RB, Slatkin M (1976) Costs and benefits of lizard thermoregulation. Quart Rev Biol 51:363–384
Huey RB, Stevenson RD (1979) Integrating thermal physiology and ecology of ectotherms: a discussion of approaches. Am Zool 19:357–366
Huey RB, Deutsch CA, Tewksbury JJ, Vitt LJ, Hertz PE, Perez HJA, Garland T (2009) Why tropical forest lizards are vulnerable to climate warming. Proc Roy Soc B-Biol Sci 276:1939–1948
Jaksic FM, Silva SI, Meserve PL, Gutierrez JR (1997) A long-term study of vertebrate predator responses to an El Nino (ENSO) disturbance in western South America. Oikos 78:341–354
Kearney M, Porter WP (2004) Mapping the fundamental niche: physiology, climate, and the distribution of a nocturnal lizard. Ecology 85:3119–3131
Kingsolver JG, Woods HA (1997) Thermal sensitivity of growth and feeding in Manduca sexta caterpillars. Physiol Zool 70:631–638
Le Galliard JF, Marquis O, Massot M (2010) Cohort variation, climate effects and population dynamics in a short-lived lizard. J Anim Ecol 79:1296–1307
Le Galliard J-F, Massot M, Baron J-P, Clobert J (2012) Ecological effects of climate change on European reptiles. In: Brodie J, Post E, Doak D (eds) Wildlife conservation in a changing climate. University of Chicago Press, Chicago, pp 179–203
Lelièvre H, Le Hénanff M, Blouin-Demers G, Naulleau G, Lourdais O (2010) Thermal strategies and energetics in two sympatric colubrid snakes with contrasted exposure. J Comp Physiol B 180:415–425
Lelièvre H, Blouin-Demers G, Pinaud D, Lisse H, Bonnet X, Lourdais O (2011) Contrasted thermal preferences translate into divergences in habitat use and realized performance in two sympatric snakes. J Zool 284:265–275
Lelièvre H, Legagneux P, Blouin-Demers G, Bonnet X, Lourdais O (2012) Trophic niche overlap in two syntopic colubrid snakes (Hierophis viridiflavus and Zamenis longissimus) with contrasted lifestyles. Amphibia-Reptilia 33:37–44
Lenk P, Joger U, Wink M (2001) Phylogenetic relationships among European ratsnakes of the genus Elaphe Fitzinger based on mitochondrial DNA sequence comparisons. Amphibia-Reptilia 22:329–339
Lima M, Stenseth NC, Leirs H, Jaksic FM (2003) Population dynamics of small mammals in semi-arid regions: a comparative study of demographic variability in two rodent species. Proc Roy Soc Ser B-Biol Sci 270:1997–2007
Lourdais O, Shine R, Bonnet X, Guillon M, Naulleau G (2004) Climate affects embryonic development in a viviparous snake, Vipera aspis. Oikos 104:551–560
Luiselli L (2005) Snakes don’t shrink, but ‘shrinkage’ is an almost inevitable outcome of measurement error by the experimenters. Oikos 110:199–202
Luiselli L (2006) Ecological modelling of convergence patterns between European and African ‘whip’ snakes. Acta Oecol 30:62–68
Luiselli L, Capizzi D (1997) Influences of area, isolation and habitat features on distribution of snakes in Mediterranean fragmented woodlands. Biodiv Conserv 6:1339–1351
Madsen T, Shine R (2000) Rain, fish and snakes: climatically driven population dynamics of Arafura filesnakes in tropical Australia. Oecologia 124:208–215
Marquis O, Massot M, Le Galliard JF (2008) Intergenerational effects of climate generate cohort variation in lizard reproductive performance. Ecology 89:2575–2583
McDonald PG, Olsen PD, Cockburn A (2004) Weather dictates reproductive success and survival in the Australian brown falcon Falco berigora. J Anim Ecol 73:683–692
Michel CL, Bonnet X (2010) Contrasted thermal regimes do not influence digestion and growth rates in a snake from temperate climate. Physiol Biochem Zool 83:924–931
Mondal S, Rai U (2001) In vitro effect of temperature on phagocytic and cytotoxic activities of splenic phagocytes of the wall lizard, Hemidactylus flaviviridis. Comp Biochem Physiol 129:391–398
Naulleau G (1984) Les serpents de France Revue Française d’Aquariologie, 11è année, 3 et 4, Paris (in French)
Naulleau G (1997a) Coluber viridiflavus (Lacépède, 1789). In: Gasc JP, Cabela A, Crnobrnja-Isailovic J, Dolmen D, Grossenbacher K, Haffner P, Lescure J, Martens H, Martinez Rica JP, Maurin H, Oliveira ME, Sofianidou TS, Veith M, Zuiderwijk A (eds) Atlas of amphibians and reptiles in Europe. Societas Herpetologica et Muséum National d’Histoire Naturelle, Paris, pp 342–343
Naulleau G (1997b) Elaphe longissima (Laurenti, 1768). In: Gasc JP, Cabela A, Crnobrnja-Isailovic J, Dolmen D, Grossenbacher K, Haffner P, Lescure J, Martens H, Martinez Rica JP, Maurin H, Oliveira ME, Sofianidou TS, Veith M, Zuiderwijk A (eds) Atlas of amphibians and reptiles in Europe. Societas Herpetologica et Muséum National d’Histoire Naturelle, Paris, pp 356–357
Naulleau G (2003) Evolution de l’aire de répartition en France, en particulier au Centre Ouest, chez trois serpents: extension vers le nord (la Couleuvre verte et jaune, Coluber viridiflavus Lacépède et la Vipère aspic Vipera aspis Linné) et régression vers le Nord (la Vipère péliade, Vipera berus Linné). Biogeographica 79:59–69 (in French with English abstract)
Ojanguren AF, Braña F (2000) Thermal dependence of swimming endurance in juvenile brown trout. J Fish Biol 56:1342–1347
Pike DA, Pizzatto L, Pike BA, Shine R (2008) Estimating survival rates of uncatchable animals: the myth of high juvenile mortality in reptiles. Ecology 89:607–611
Pollock KH, Nichols JD, Brownie C, Hines JE (1990) Statistical inference for capture-recapture experiments. Wildl Monogr 107:1–97
Rugiero L, Capizzi D, Luiselli L (2002) Interactions between sympatric snakes, Coluber viridiflavus and Elaphe longissima: are there significant inter-annual differences in coexistence patterns? Ecol Mediterr 28:75–91
Sæther BE, Tufto J, Engen S, Jerstad K, Rostad OW, Skatan JE (2000) Population dynamical consequences of climate change for a small temperate songbird. Science 287:854–856
Scheers H, Van Damme R (2002) Micro-scale differences in thermal habitat quality and a possible case of evolutionary flexibility in the thermal physiology of Lacertid lizards. Oecologia 132:323–331
Secor SM, Nagy KA (1994) Bioenergetic correlates of foraging mode for the snakes Crotalus cerastes and Masticophis flagellum. Ecology 75:1600–1614
Selas V (2001) Predation on reptiles and birds by the common buzzard, Buteo buteo, in relation to changes in its main prey, voles. Can J Zool 79:2086–2093
Shine R, Bonnet X (2009) Reproductive biology, population viability, and options for field management. In: Mullin SJ, Seigel RA (eds) Snakes: Ecology and conservation. Cornell University Press, Ithaca, pp 172–200
Sinervo B, Méndez-de-la-Cruz F, Miles DB, Heulin B, Bastiaans E, Villagrán-Santa Cruz M, Lara-Resendiz R, Martínez-Méndez N, Calderón-Espinosa ML, Meza-Lázaro RN, Gadsden H, Avila LJ, Morando M, De la Riva IJ, Sepulveda PV, Duarte Rocha CF, Ibargüengoytía N, Puntriano CA, Massot M, Lepetz V, Oksanen TA, Chapple DG, Bauer AM, Branch WR, Clobert J, Sites JW Jr (2010) Erosion of lizard diversity by climate change and altered thermal niches. Science 328:894–899
Stenseth NC, Mysterud A, Ottersen G, Hurrell JW, Chan KS, Lima M (2002) Ecological effects of climate fluctuations. Science 297:1292–1296
Stevens M, Yule DH, Ruxton GD (2008) Dazzle coloration and prey movement. Proc Roy Soc B-Biol Sci 275:2639–2643
Stevenson RD, Peterson CR, Tsuji J (1985) The thermal dependence of locomotion, tongue flicking, digestion, and oxygen consumption in the wandering garter snake. Physiol Zool 58:46–57
Weatherhead PJ, Sperry JH, Carfagno GLF, Blouin-Demers G (2012) Latitudinal variation in thermal ecology of North American ratsnakes and its implications for the effect of climate warming on snakes. J Therm Biol 37:273–281
Webb JK, Whiting MJ (2005) Why don’t small snakes bask? Juvenile broad-headed snakes trade thermal benefits for safety. Oikos 110:515–522
Webb JK, Brook BW, Shine R (2003) Does foraging mode influence life history traits? A comparative study of growth, maturation and survival of two species of sympatric snakes from south-eastern Australia. Austral Ecol 28:601–610
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46:120–139
Acknowledgments
This research was made possible by the financial support of the Conseil Général des Deux-Sèvres, the Région Poitou–Charentes, and the ANR (ECTOCLIM project). We warmly thank the ONF for authorizing X. Bonnet to set up the capture-mark-recapture study in the forest of Chizé and for continuous logistical support. We are also very grateful to all the students who actively participated to the mark-recapture survey since 1997. We also thank G. Naulleau who initiated field surveys in the CÉBC.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lelièvre, H., Rivalan, P., Delmas, V. et al. The thermoregulatory strategy of two sympatric colubrid snakes affects their demography. Popul Ecol 55, 585–593 (2013). https://doi.org/10.1007/s10144-013-0388-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10144-013-0388-z