
Abstract
For the first time an international acoustic survey dataset covering three decades was used to investigate the factors shaping the spatial and temporal patterns in the condition of sprat and herring in the Baltic Proper. Generalized additive models showed that the spatial and temporal fluctuations in sprat density have been the main drivers of the spatio-temporal changes of both sprat and herring condition, evidencing intra- and inter-specific density dependence mediated by the size and distribution of the sprat population. Salinity was also an important predictor of herring condition, whereas temperature explained only a minor part of sprat model deviance. Herring density was an additional albeit weak significant predictor for herring condition, evidencing also intra-specific density dependence within the herring population. For both species, condition was high and similar in all areas of the Baltic Proper until the early 1990s, coincident with low sprat densities. Afterwards, a drop in condition occurred and a clear south–north pattern emerged. The drop in condition after the early 1990s was stronger in the northern areas, where sprat population increased the most. We suggest that the increase in sprat density in the northern areas, and the consequent spatial differentiation in clupeid condition, have been triggered by the almost total disappearance of the predator cod from the northern Baltic Proper. This study provides a step forward in understanding clupeid condition in the Baltic Sea, presenting evidence that density-dependent mechanisms also operate at the spatial scale within stock units. This stresses the importance of spatio-temporal considerations in the management of exploited fish.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Marine ecosystems are severely impacted by overfishing and climate change. Largely restructured food webs are often the result of these perturbations (Cury et al. 2008) with important implications for the goods and services these ecosystems provide to humans (Hughes et al. 2000). The developing concept of ecosystem-based management requires detailed knowledge on, and integration of, the spatial and temporal processes leading to the observed ecosystem changes (Marasco et al. 2007).
Morphometric condition indices based on fish weight and length are often used in fisheries science and fish population dynamics to compare the well-being or condition of individuals or populations, based on the hypothesis that heavier fish of given length are in better condition (Bagenal and Tesch 1978). As such, condition is a crucial parameter for the success of fish populations, since it may influence individual survival (Lambert and Dutil 2000), migration patterns (Slotte 1999), reproductive output (Blanchard et al. 2003; Trippel and Neil 2004) and recruitment (Marshall and Frank 1999). Condition may be dependent on several, often inter-connected, factors such as density dependence (Winters and Wheeler 1994), hydro-climatic conditions (Lloret and Rätz 2000), parasitic infection (Bagamian et al. 2004) and selective fishing (Vainikka et al. 2009). Therefore, knowledge of the temporal as well as spatial variability of condition as a response to larger ecosystem changes is crucial for understanding and forecasting the dynamics of fish stocks.
During the last three decades, the central Baltic Sea (the Baltic Proper, Fig. 1) has experienced considerable changes on all trophic levels, which show the characteristics of ecosystem regime shifts and trophic cascades (Casini et al. 2008, 2009; Möllmann et al. 2008, 2009). A major component of these changes is the switch from cod (Gadus morhua) to sprat (Sprattus sprattus) domination in the fish community [International Council for the Exploration of the Sea (ICES 2009)]. Since the early 1990s, the release from predation by the collapsed cod stock, intertwined with an increase in seawater temperature, has triggered a drastic increase in the sprat stock (MacKenzie and Köster 2004; Casini et al. 2008). In this shifting ecological scenario, the growth and condition of sprat and herring (Clupea harengus), which are the main pelagic fish species in the Baltic Sea, has also experienced remarkable changes (Cardinale and Arrhenius 2000; Cardinale et al. 2002; ICES 2009). These changes have important implications not only for fish population dynamics, e.g., fecundity and reproductive potential (Laine et al. 1998; Laine and Rajasilta 1999; Cardinale et al. 2009), but also for predictions and thus management of the Baltic fisheries (ICES 2008a).

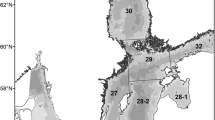
Map of the Baltic Sea. The study area is marked in dark gray (Baltic Proper) and corresponds to ICES subdivisions (SDs) 25–29. The names of the countries surrounding the Baltic Proper are indicated
In the Baltic Proper, temporal variations in clupeid condition have been related before to either hydro-climate changes (i.e., salinity, Möllmann et al. 2003), density-dependent factors and feeding competition (Möllmann et al. 2005; Casini et al. 2006), or a combination of the two forces prevailing in different periods (Casini et al. 2010). However, the spatial aspect in the investigations of clupeid condition has not been considered so far in the Baltic Sea. Especially, the question of how area-specific ecosystem changes have affected condition of herring and sprat in the entire Baltic Proper has not been addressed. To our best knowledge, large-scale spatial investigations of fish condition for pelagic stocks are very scarce in literature (but see Husebø et al. 2007).
Here we used for the fist time a unique, extensive dataset of sprat and herring biological data collected during international acoustic surveys in 1978–2008 by several countries bordering the Baltic Sea. The aim of our study is to investigate how the temporal and spatial structural changes in the Baltic Proper ecosystem affected the condition of sprat and herring. Specifically, the extensive material allowed us (1) to model the spatio-temporal changes in clupeid condition using spatially resolved biotic and abiotic time series, and (2) to investigate the potential spatial differences in clupeid condition and the underlying causes.
Methods
Data
Fish sampling and condition computation
Sprat and herring were collected in September–October during the period 1978–2008 in the open areas of the Baltic Proper [corresponding to ICES subdivisions (SD) 25–29, Fig. 1]. The fish samples were collected by the countries participating in the autumn Baltic International Acoustic Surveys (BIAS), under the coordination of ICES (2008b). Fishing was carried out using mid-water trawls in the pelagic zone and near the bottom layers, depending on the fish vertical distribution detected by echosounder (see ICES 2008b for details on fishing operation and sampling procedure). Prior to the statistical analysis, fish body weights were rounded to the nearest 1 g and total lengths were rounded down to the nearest 0.5 cm.
For estimation of fish condition, a year-specific regression of ln L (total length) versus ln W (total weight) was fitted. Thereafter, the condition was estimated as the weight at the grand mean length of the dataset, i.e., 12 cm for sprat and 18 cm for herring. This approach is particularly suitable when studying pelagic fish (Winters and Wheeler 1994; Tanasichuk 1997), and it has been previously used for estimating the condition of sprat and herring in the Baltic Sea (e.g., Cardinale et al. 2002; Möllmann et al. 2003; Casini et al. 2006). To avoid the use of age groups consisting of only few specimens (i.e., <10 individuals), analysis was restricted to ages 1–5 for sprat and 1–7 for herring. The number of fish used in the condition calculation by year and SD, along with the country of origin, is presented in Table 1.
The database we used was constituted by a mixture of fish that had already spawned at least once (generally ages 2+) and juvenile immature fish (most of age 1 fish) (ICES 2009). The sporadic information on maturity stage in the database did not allow us to account for it in the computation of the condition time series. To test the impact of maturity stage on the condition time series, we compared a time series where all the fish were used (ages 1+) with a time series where only repeated spawners were included (ages 2+). The time series were highly correlated (R 2 in the range 0.95–0.99 for herring, and 0.94–0.99 for sprat, in different SDs). This indicates that the inclusion of juvenile fish (age 1) did not have an impact on the condition time series and further analyses.
Previous investigations have shown that the seasonal variations in morphometric condition indices, calculated based on fish weight and length, of clupeids (e.g., Baltic sprat and spring-spawning Atlantic herring) follow the seasonal changes in the lipid content of their muscles (Kondratjeva 1993; Winters and Wheeler 1994). These studies strongly support that the condition computed in our study reflects the fat and energy content of the fish.
Stock size
Time series of sprat and herring stock abundance (total number, ages 1+) by SD were calculated based on autumn hydroacoustic measurements regularly used as tuning indices in stock assessment (ICES 2009). The data were extracted from the ICES BAD1 database (ICES 2008b) and survey reports stored at the Swedish Board of Fisheries, Institute of Marine Research. Subsequently, sprat and herring stock density (number × km−3) by SD was calculated. The water volume in the different SDs was extracted from the data assimilation system (DAS) provided by the Baltic Nest Institute of Stockholm University. We restricted the density of sprat and herring to the 0–100 m depth according to the vertical distribution of clupeids in the Baltic Proper (Orlowski 2005). Only for SD 25, the water volume was calculated between 0 and 80 m due to the shallower depth of this area. Biomasses of cod stock in SDs 25, 26 and 28 were made available by the Study Group on Multispecies Assessment in the Baltic (ICES 2006).
Hydro-climate
Data on summer water temperature and salinity for each SD were extracted from the Helsinki Commission (HELCOM) database stored on the ICES website (http://www.ices.dk). Water temperature (°C) and salinity (psu) values were averaged between 0 and 100 m depth (samples at surface and at 10 m depth intervals), reflecting the depth distribution of the clupeids in the Baltic Proper. Only for SD 25, the average was taken between 0 and 80 m (same depth intervals) due to the shallower depth of this area. Summer is the main feeding and growth season of Baltic clupeids, and therefore is a critical period in determining sprat and herring condition in the early autumn (Casini et al. 2006).
Statistical analysis
Time series of annual condition of sprat and herring were calculated for each country and SD. Successively, a generalized linear model (GLZM) was used to predict the year effect on clupeid condition for each SD after scaling out the country effect. Typically, during the survey each country covers a specific area within the same SD. Therefore, this correction allowed accounting for potential spatial differences in condition within a SD when its area was not entirely covered by the survey. Condition data were normally distributed, and thus the normal distribution with an identity function was used in the GLZMs.
To analyse the effect of different predictors on clupeid condition we used generalized additive models (GAMs, Hastie and Tibshirani 1990). The following additive formulation was used:
where a is the intercept, s is the thin-plate smoothing spline function (Wood 2003), \( V_{i} , \ldots ,V_{n} \) are the predictors, and ε is the random error. Y and SD indicate year and subdivision, respectively.
As predictors for clupeid condition (response) the following variables were employed in the full models: top-down density-dependent forces (i.e., sprat density and herring density) and hydrological data (summer salinity and temperature). These predictors were selected based on acknowledged ecological and physiological mechanisms (Casini et al. 2006). Clupeid density in number, rather than biomass, was used in the GAMs because biomass contains already by definition a growth signal, making response (condition) and predictor (biomass) not independent a priori. All variables were log-transformed and expressed as standardized anomalies prior to analyses.
We retained only the predictors that were statistically significant (final model) using a backward stepwise procedure. From the full model, the non-significant predictor with the lowest significance level was excluded at each step and the model run again. This procedure was repeated until all the predictors were significant. We limited the maximum degrees of freedom acceptable for each term to k = 4. A normal distribution with an identity function was used in the GAMs. We calculated the deviance explained by the final models and the percent explained deviance (PED) of each predictor selected in the final models. PED was estimated by comparing the deviance explained by the models before and after a specific predictor was subtracted from the final model. Residuals were inspected using graphical methods (Cleveland 1993).
We also analysed clupeid condition applying GAMs for each SD separately, using the same procedure as explained above.
The statistical analyses were performed using the mgcv library of R 2.10.0 (www.r-project.org) and Statistica 6.1. The significance level was set to α = 0.05 for all tests.
Results
Spatio-temporal patterns in clupeid condition, clupeid density and hydrological factors
The temporal development of clupeid condition in the different SDs of the Baltic Proper is shown in Fig. 2a, b. Sprat condition showed high values from the late 1970s up to the early 1990s, followed by a drop which ended in 1996–1998. Afterwards, sprat condition has oscillated, although never reaching the values of the 1970s to 1980s (Fig. 2a). Herring condition showed a similar trend, even though the decrease at the beginning of the time series started earlier than for sprat (Fig. 2b). These patterns were common for all the SDs of the Baltic Proper. However, the decrease in the condition of both sprat and herring in the early 1990s was more evident in the northern SDs. In fact, condition was very similar in all SDs in 1978–1991, whereas evident spatial differences occurred afterwards, for both species (Fig. 2a–d). Threshold generalized additive models (TGAMs, Ciannelli et al. 2004) confirmed that clear-cut differences in fish condition between SDs became apparent only after the early 1990s [Fig. A1 in Electronic Supplementary Material (ESM)].

Temporal trends in a sprat condition and b herring condition in the different SDs. c, d Mean sprat and herring condition in 1978–1991 and 1992–2008, respectively (bars show 95% confidence intervals)
Acoustic measurements revealed that sprat density followed opposite trends to clupeid condition. Sprat density was low in all the SDs of the Baltic Proper from the late 1970s to the early 1990s, followed by a steep rise which culminated in the mid-late 1990s (Fig. 3a). The increase in density was mostly evident in the northern areas (SDs 27–29) of the Baltic Proper (Fig. 3a, b). Since the mid-1990s, sprat density has oscillated at relatively high levels. TGAMs (Ciannelli et al. 2004) confirmed that clear-cut differences in sprat density among SDs became apparent only after the early 1990s (Fig. A1 in ESM).

a Temporal trends in sprat density in the different SDs. b Mean sprat density in 1978–1991 and 1992–2008 (bars show 95% confidence intervals)
Distribution maps were produced to visualize at higher resolution the spatial patterns in fish condition and sprat abundance below and above the temporal threshold (Fig. 4). Mean condition was computed by averaging the data from each ICES statistical rectangle (0.5° in latitude and 1° in longitude) in the periods 1984–1991 and 1992–2008. Data prior to 1984 were not available at this spatial resolution and thus could not be used to create the maps. In the period 1984–1991, fish condition and sprat abundance were spatially homogeneous, whereas in the period 1992–2008 they were more spatially heterogeneous and a clear south–north gradient emerged (Fig. 4).

Spatial patterns in mean clupeid condition (a, b for sprat and c, d for herring) and sprat abundance (e, f) in 1984–1991 and 1992–2008. Only years with at least 10 individuals per rectangle were used in the condition calculation
Herring density showed oscillations that were very similar in all the SDs, although SD 29 presented some isolated peaks in certain years not observed in the other SDs (Fig. 5a). After the sprat outburst, herring density overall has been considerably lower than sprat density in all the SDs (Fig. 5a). Salinity decreased up to the early 1990s, whereas an increase has been noticeable thereafter. These patterns were generally common to all the areas, although in SD 25 the changes in salinity were not as evident as in the other SDs (Fig. 5b). Temperature increased throughout the study period in all SDs (Fig. 5c). A clear south–north gradient in both salinity and temperature is noticeable along the whole time series, with higher values consistently observed in the southern areas (Fig. 5b, c).

Temporal trends in a herring density, b salinity 0–100 m (0–80 m in SD 25), and c temperature 0–100 m (0–80 m in SD 25), in the different SDs
Modelling clupeid condition
The results of the GAMs using the whole dataset are presented in Table 2 and Fig. 6. Sprat and herring final models explained, respectively, 71.2% and 54% of the total deviance. Sprat density was the most important predictor (negative effect, Fig. 6a, c) of both sprat and herring condition, explaining 97.2% and 68.9% of the model deviance (PED), respectively. Temperature was also a significant predictor for sprat condition (positive effect, Fig. 6b), although it explained only 2.8% of the model deviance. Herring density did not have any effect on sprat condition. Herring density and salinity were also significant predictors for herring condition, explaining 4.4% and 26.7% of the model deviance (negative and positive asymptotic effect, respectively, Fig. 6d, e). Overall, the density-dependent factors (sprat + herring densities) explained 73.3% of the deviance of the herring condition model (Table 2). The analyses of the residuals of the GAMs are presented in Fig. A2 in ESM.

Results of the GAMs (final models). The partial effects of each predictor on sprat and herring condition are shown. See Table 2 for details on the statistics
Considering each SD separately, sprat density was the most important predictor of sprat condition in all the SDs, whereas temperature was also included in the sprat final model for SD 27 (Table A1 in ESM). Sprat density was also the most important predictor for herring condition in all the SDs, whereas salinity was included in the herring final model for SD 28 (Table A1 in ESM). The analyses of the residuals of the GAMs for each SD are presented in Figs. A3 and A4 in ESM.
The use of interaction terms did not improve the final models, and therefore they are not discussed further. The use of 1-year time lag between the predictors (year t) and clupeid condition (year t + 1) decreased the explained deviance of all final models.
When spring salinity and temperature data were included in the GAMs as additional predictors, they were discarded by the backward stepwise procedure and thus excluded from all the final models.
Discussion
We investigated the spatio-temporal patterns in condition of sprat and herring during a period of major ecosystem modifications in the Baltic Proper. The restructuring of the ecosystem, characterized by a drop in the predator cod and an outburst of the sprat stock (ICES 2009), was clearly accompanied by drastic changes in the condition of sprat and herring in the whole Baltic Proper. These changes were, however, more evident in the northern areas where the sprat density changed the most. Overall, our study suggests that, during the past 30 years, density dependence, mediated by the large spatial and temporal variations in sprat stock size, has been the main regulator of the spatio-temporal changes of both sprat and herring condition in the Baltic Proper.
Although correlations do not prove a cause–effect link between variables, we believe that the negative relation between sprat density and clupeid condition is truly mechanistic. The large diet overlap between sprat and herring (Casini et al. 2004; Möllmann et al. 2004) suggests the existence of both intra- and inter-specific food competition. It has been shown that sprat abundance correlates negatively to zooplankton summer biomass and clupeid stomach fullness in SD 28, this providing an ecological explanation of the negative relation between sprat abundance and clupeid condition in this area (Casini et al. 2006). Bioenergetic studies have also supported the empirical evidence that clupeid predation can affect zooplankton dynamics (e.g., Rudstam et al. 1992; Raid et al. 2010). Overall, these observations indicate that the density dependence found in our study acts at least partly through food competition. However, we also found that the effect of sprat density was stronger on sprat than on herring condition. This is likely due to the omnivore nature of larger herring, feeding not only on zooplankton but also on zoobenthos (Aneer 1980; Raid and Lankov 1995; Szypuła et al. 1997; Casini et al. 2004; Möllmann et al. 2004), decreasing the interaction strength with sprat. Moreover, the low density of herring, if compared with sprat, could be the reason for the weak intra-specific density dependence within the herring population.
Density-dependent growth and condition has been suggested for other pelagic fish stocks around the world (see Husebø et al. 2007 and references therein). In most studies, the density dependence has been shown as negative relation between fish body size and stock abundance of a single species (e.g., herring, Winters and Wheeler 1994; Tanasichuk 1997; Melvin and Stephenson 2007), whereas in the Baltic Proper both sprat and herring condition are linked to the sprat stock, evidencing the occurrence of intra- as well as inter-specific density dependence. The importance of considering the dynamics of potentially competing species in condition studies has also been shown for other species, e.g., mackerel (Scomber japonicus) and jack mackerel (Trachurus symmetricus) in Southern California (Parrish and Mallicoate 1995).
The effects of temperature and salinity found in our study likely act both directly on fish metabolism (Jobling 1994) and indirectly through regulation of food resources (Möllmann et al. 2005). Temperature was positively albeit weakly related to sprat condition, whereas salinity had a positive effect on herring condition that reached a plateau at high salinity values. This is in agreement with the general understanding of the dynamics of these two species in the Baltic Sea. Sprat is considered a thermophilic species, with higher temperature being also favourable for sprat egg and larval survival and therefore recruitment success (Nissling 2004). The link between herring condition and salinity, on the other side, could be mediated through the availability of one of the main prey for herring, the marine copepod Pseudocalanus spp., whose reproductive performance is enhanced at high salinities (Möllmann et al. 2003). This plankter, however, is also under the strong influence of sprat predation (Möllmann et al. 2008), this explaining the strong inter-specific density dependence found for herring condition.
Our analysis encompassing the whole Baltic Proper is a follow-up of previous studies (Casini et al. 2006) which have shown the density-dependent nature of clupeid condition in restricted areas of the Baltic Sea. However, our study adds a spatial dimension to the temporal one. In fact, previous investigations lacked spatially resolved time series of sprat and herring density, and the impact of the different factors on fish condition in a large area of clupeid distribution could not be investigated.
The newly available, spatially resolved data allowed us to find that the decrease in clupeid condition has been generally stronger in the northern than in the southern Baltic Proper. An inverse trend was shown by sprat density, which increased mostly in the northern SDs, shifting the centre of distribution of the sprat stock to the northern Baltic Proper after the early 1990s. These results lead to two major conclusions:
-
1.
The permanent southwest–northeast gradient in hydrological features characterizing the Baltic Proper does not directly translate into a similar clear-cut geographical gradient in clupeid condition, which the use of only the most recent data would instead wrongly suggest. A weak south–north gradient in condition was observed also in the 1970s to 1980s, but after the early 1990s the gradient became notably stronger.
-
2.
The clear spatial pattern in clupeid condition that occurred after the early 1990s is mainly due to the geographical reallocation of the sprat population into the northern SDs of the Baltic Proper. This implies that density-dependent processes in this region are acting not only on inter-annual basis (Casini et al. 2006; Möllmann et al. 2005) but also at a spatial scale. We suggest that the geographical reallocation of the sprat population into the northern Baltic Proper was triggered by the stronger decrease in the population of its main predator, the cod, in these areas (see also ICES 2006, Fig. 7). The shift of the centre of distribution of the sprat population to northern areas was unlikely caused by hydro-climatic changes, since sprat is favoured by warmer conditions (e.g., Nissling 2004; Alheit et al. 2005; MacKenzie and Köster 2004) encountered mainly in the southern Baltic Proper. However, the general warming of the Baltic Sea, intertwined with the increase in average salinity since the mid-1990s (Fig. 5b, c), might have created less harsh hydro-climate circumstances for sprat in the northern SDs, and therefore facilitated the northward expansion of its population when the cod predation pressure became negligible in these areas.
Fig. 7 
Percentage of the sprat stock in the northern Baltic Proper (SDs 27–29) as calculated by acoustic surveys, and percentage of the cod stock in the northern Baltic Proper (SD 28) from MultiSpecies Virtual Population Analysis (MSVPA, ICES 2006). Trends in cod stock size from MSVPA are not available for SDs 27 and 29

It is noteworthy that the herring stocks inhabiting the areas adjacent to SDs 28–29 (i.e., the Gulf of Riga and Bothnian Sea stocks) have increased during the past decades (ICES 2009). The intra-specific density dependence within the herring population found in our study could therefore have been facilitated by spillover of these adjacent herring stocks into the northern Baltic Proper. There are also indications that the population of three-spined stickleback (Gasterosteus aculeatus) has increased in the northern Baltic Proper (Ljunggren et al. 2010). The increase of this additional planktivorous fish could have amplified the intra-specific feeding competition (Peltonen et al. 2004), contributing to the strong decrease in clupeid condition in these areas. The potential impact of three-spined stickleback on sprat and herring growth is worth further analyses. Additionally, the potential effects of size-selective fishing pressure might be also explored in future investigations on Baltic clupeid condition (Vainikka et al. 2009). In our study this could not be accomplished because of the lack of spatially resolved size-selectivity data.
To our knowledge, within-stock spatial variations in pelagic fish condition, and investigations of their potential causes, have been presented only by Husebø et al. (2007) for Norwegian spring-spawning herring. In Husebø et al. (2007) it was shown that the permanent south–north gradient in temperature along the Norwegian coast creates a latitudinal pattern in the condition and growth of juvenile herring in their nursery grounds, even though local herring stock abundance was also found to have an effect. Compared with Husebø et al. (2007), we show for the Baltic Proper that, during the past 30 years, the direct hydro-climate effects have been of secondary importance for clupeid condition, which has been mainly regulated by the spatial distribution of the sprat stock. Hydro-climate, however, has likely operated indirectly on clupeid condition, facilitating the northern distribution of the sprat population (see discussion above), but also affecting the distribution of its main predator, the cod. In fact, the decrease in salinity up to the mid-1990s caused a contraction of the cod population to the southern and more salty areas (Figs. 5b, 7; ICES 2006), where cod still encountered suitable conditions for reproduction. After the mid-1990s, however, despite an increase in deep-water salinity, the cod stock did not show a geographical re-expansion, likely due to its enduring very low population size linked to high fishing pressure (ICES 2009). Therefore, a combination of hydrological and fishery-related factors have created long-lasting particularly advantageous circumstances for sprat in the northern areas of the Baltic Proper, where cod predation pressure has been negligible for nearly two decades (ICES 2006). An analytical investigation of the reasons behind the changes in spatial distribution of sprat is beyond the aim of our paper.
The timing (early 1990s) of the observed spatio-temporal changes in clupeid condition and sprat stock distribution coincides with other temporal shifts in ecosystem structure and functioning revealed in the Baltic Proper by previous investigations (Casini et al. 2008, 2009; Möllmann et al. 2008, 2009). Accordingly, a regime shift has been shown for the Baltic Proper in the early 1990s (Möllmann et al. 2009) coupled with a threshold-like switch in specific food-web interactions (Casini et al. 2009, 2010). Our study reveals for the first time also the occurrence of spatial shifts within the Baltic Proper. This highlights the fact that spatial differences in some biological features may arise among the SDs of the Baltic Proper, and that therefore the magnitude of the changes can be area specific. This may have important implications for the management of exploited fish, since in the Baltic Proper both herring and sprat are currently assessed and managed as single stock units without spatial considerations.
By using extensive spatially resolved biological, acoustic and abiotic datasets we have here for the first time provided empirical evidence that the temporal and spatial changes in clupeid condition in the Baltic Proper are a result of the temporal and spatial dynamics of the pelagic fish competing for common resources. Especially we have shown that (1) during the last 30 years, temporal fluctuations in both sprat and herring condition in all areas of the Baltic Proper can be linked to drastic variations in sprat density, and (2) clear spatial differences in clupeid condition have arisen after the early 1990s in response to large changes in the geographical distribution of the sprat population. We suggest that the spatial reallocation of the sprat population to northern SDs occurred mainly because of the disappearance of its main predator, the cod, from these areas.
The spatial issue in fish population dynamics has only started to gain attention in fisheries research very recently (Hsieh et al. 2010). However, as shown by other investigations (e.g., Kritzer and Sale 2004; Hsieh et al. 2008) and the present study, spatial considerations are crucial for understanding fish population dynamics and therefore for the management of exploited stocks.
References
Alheit J, Möllmann C, Dutz J, Kornilovs G, Loewe P, Mohrholz V, Wasmund N (2005) Synchronous ecological regime shifts in the central Baltic and the North Sea in the late 1980s. ICES J Mar Sci 62:1205–1215
Aneer G (1980) Estimates of feeding pressure on pelagic and benthic organisms by Baltic herring (Clupea harengus v. membras L.). Ophelia 1(Suppl):265–275
Bagamian KH, Heins DC, Baker JA (2004) Body condition and reproductive capacity of three-spined stickleback infected with the cestode Schistocephalus solidus. J Fish Biol 64:1568–1576
Bagenal TB, Tesch FW (1978) Age and growth. In: Bagenal T (ed) Methods for assessment of fish production in fresh waters. Blackwell, Oxford, pp 101–136
Blanchard JL, Frank KT, Simon JE (2003) Effects of condition on fecundity and total egg production of eastern Scotian Shelf haddock (Melanogrammus aeglefinus). Can J Fish Aquat Sci 60:321–332
Cardinale M, Arrhenius F (2000) Decreasing in weight-at-age of Baltic herring (Clupea harengus) between 1986 and 1996: a statistical analysis. ICES J Mar Sci 57:882–893
Cardinale M, Casini M, Arrhenius F (2002) The influence of biotic and abiotic factors on the growth of sprat (Sprattus sprattus) in the Baltic Sea. Aquat Living Resour 15:273–281
Cardinale M, Möllmann C, Bartolino V, Casini M, Kornilovs G, Raid T, Margonski P, Grzyb A, Raitaniemi J, Gröhsler T, Flinkman J (2009) Effect of environmental variability and spawner characteristics on the recruitment of Baltic herring Clupea harengus populations. Mar Ecol Progr Ser 388:221–234
Casini M, Cardinale M, Arrhenius F (2004) Feeding preferences of herring (Clupea harengus) and sprat (Sprattus sprattus) in the southern Baltic Sea. ICES J Mar Sci 61:1267–1277
Casini M, Cardinale M, Hjelm J (2006) Inter-annual variation in herring (Clupea harengus) and sprat (Sprattus sprattus) condition in the central Baltic Sea: what gives the tune? Oikos 112:638–650
Casini M, Lövgren J, Hjelm J, Cardinale M, Molinero JC, Kornilovs G (2008) Multi-level trophic cascades in a heavily exploited open marine ecosystem. Proc R Soc B Biol Sci 275:1793–1801
Casini M, Hjelm J, Molinero JC, Lövgren J, Cardinale M, Bartolino V, Belgrano A, Kornilovs G (2009) Trophic cascades promote threshold-like shifts in pelagic marine ecosystems. Proc Natl Acad Sci USA 106:197–202
Casini M, Bartolino V, Molinero JC, Kornilovs G (2010) Linking fisheries, trophic interactions and climate: threshold dynamics drive herring Clupea harengus growth in the central Baltic Sea. Mar Ecol Progr Ser 413:241–252
Ciannelli L, Chan KS, Bailey K, Stenseth NC (2004) Nonadditive effect of the environment on the survival of a large marine fish population. Ecology 85:3418–3427
Cleveland WS (1993) Visualizing data. Hobart, Summit
Cury PM, Shin Y-J, Planque B, Durant JM, Fromentin J-M, Kramer-Schadt S, Stenseth NC, Travers M, Grimm V (2008) Ecosystem oceanography for global change in fisheries. Trends Ecol Evol 23:338–346
Hastie T, Tibshirani R (1990) Generalized additive models. Chapman & Hall, London
Hsieh CH, Reiss SC, Hewitt RP, Sugihara G (2008) Spatial analysis shows fishing enhances the climatic sensitivity of marine fishes. Can J Fish Aquat Sci 65:947–961
Hsieh CH, Yamauchi A, Nakazawa T, Wang W-F (2010) Fishing effects on age and spatial structures undermine population stability of fishes. Aquat Sci 72:165–178
Hughes TP, Bellwood DR, Folke C, Steneck RS, Wilson J (2000) New paradigms for supporting the resilience of marine ecosystems. Trends Ecol Evol 20:380–386
Husebø Å, Slotte A, Stenevik EK (2007) Growth of juvenile Norwegian spring-spawning herring in relation to latitudinal and interannual differences in temperature and fish density in their coastal and fjord nursery areas. ICES J Mar Sci 64:1161–1172
ICES (2006) Report of the study group on multispecies assessment in the Baltic (SGMAB). ICES CM 2006/BCC:07. International Council for the Exploration of the Sea, Copenhagen
ICES (2008a) Report of the workshop on reference points in the Baltic Sea (WKREFBAS). ICES CM2008/ACOM:28. International Council for the Exploration of the Sea, Copenhagen
ICES (2008b) Report of the Baltic International Fish Survey Working Group (WGBIFS). ICES CM 2008/LRC:08. International Council for the Exploration of the Sea, Copenhagen
ICES (2009) Report of the Baltic Fisheries Assessment Working Group (WGBFAS). ICES CM 2009/ACOM:07. International Council for the Exploration of the Sea, Copenhagen
Jobling M (1994) Fish bioenergetics. Fish and fisheries series, vol 13. Chapman & Hall, London
Kondratjeva N (1993) Biochemical and morphophysiological parameters of Baltic sprat in relation to age and annual cycle periods. ICES CM J 25:1–13
Kritzer JP, Sale PF (2004) Metapopulation ecology in the sea: from Levins’ model to marine ecology and fisheries science. Fish Fish 5:131–140
Laine P, Rajasilta M (1999) The hatching success of Baltic herring eggs and its relation to female condition. J Exp Mar Biol Ecol 237:61–73
Laine P, Eklund J, Soikkeli M (1998) Reproduction of Baltic herring after dissimilar growth and overwintering seasons. Ophelia 48:155–165
Lambert Y, Dutil J-D (2000) Energetic consequences of reproduction in Atlantic cod (Gadus morhua) in relation to spawning level of somatic energy reserves. Can J Fish Aquat Sci 57:815–825
Ljunggren L, Sandström A, Bergström U, Mattila J, Lappalainen A, Johansson G, Sundblad G, Casini M, Kaljuste O, Eriksson BK (2010) Recruitment failure of coastal predatory fish in the Baltic Sea coincident with an offshore ecosystem regime shift. ICES J Mar Sci 67:1587–1595
Lloret J, Rätz H-J (2000) Condition of cod (Gadus morhua) off Greenland during 1982–1998. Fish Res 48:79–86
MacKenzie B, Köster FW (2004) Fish production and climate: sprat in the Baltic Sea. Ecology 85:784–794
Marasco RJ, Goodman D, Grimes CB, Lawson PW, Punt AE, Quinn TJ II (2007) Ecosystem-based fisheries management: some practical suggestions. Can J Fish Aquat Sci 64:928–939
Marshall CT, Frank KT (1999) The effect of interannual variation in growth and condition on haddock recruitment. Can J Fish Aquat Sci 56:347–355
Melvin GD, Stephenson RL (2007) The dynamics of a recovering fish stock: Georges Bank herring. ICES J Mar Sci 64:69–82
Möllmann C, Kornilovs G, Fetter M, Köster FW, Hinrichsen H-H (2003) The marine copepod, Pseudocalanus elongatus, as a mediator between climate variability and fisheries in the Central Baltic Sea. Fish Oceanogr 12:360–368
Möllmann C, Kornilovs G, Fetter M, Köster FW (2004) Feeding ecology of central Baltic Sea herring and sprat. J Fish Biol 65:1563–1581
Möllmann C, Kornilovs G, Fetter M, Köster FW (2005) Climate, zooplankton, and pelagic fish growth in the central Baltic Sea. ICES J Mar Sci 62:1270–1280
Möllmann C, Müller-Karulis B, Kornilovs G, St. John AM (2008) Effects of climate and overfishing on zooplankton dynamics and ecosystem structure: regime shifts, trophic cascades, and feedback-loops in a simple ecosystem. ICES J Mar Sci 65:302–310
Möllmann C, Diekmann R, Müller-Karulis B, Kornilovs G, Plikshs M, Axe P (2009) Reorganization of a large marine ecosystem due to atmospheric and anthropogenic pressure: a discontinuous regime shift in the Central Baltic Sea. Glob Change Biol 15:1377–1393
Nissling A (2004) Effects of temperature on egg and larval survival of cod (Gadus morhua) and sprat (Sprattus sprattus) in the Baltic Sea—implications for stock development. Hydrobiologia 514:115–123
Orlowski A (2005) Experimental verification of the acoustic characteristics of the clupeoid diel cycle in the Baltic. ICES J Mar Sci 62:1180–1190
Parrish RH, Mallicoate DL (1995) Variation in the condition factors of California pelagic fishes and associated environmental factors. Fish Oceanogr 4:171–190
Peltonen H, Vinni M, Lappalainen A, Pönni J (2004) Spatial feeding patterns of herring (Clupea harengus L.), sprat (Sprattus sprattus L.), and the three-spined stickleback (Gasterosteus aculeatus L.) in the Gulf of Finland, Baltic Sea. ICES J Mar Sci 61:966–971
Raid T, Lankov A (1995) Recent changes in the growth and feeding of Baltic herring and sprat in the north-eastern Baltic Sea. Proc Estonian Acad Sci Ecol 5:38–55
Raid T, Kornilovs G, Lankov A, Nisumaa A-M, Shpilev H, Järvik A (2010) Recruitment dynamics of the Gulf of Riga herring stock: density-dependent and environmental effects. ICES J Mar Sci 67:1914–1920
Rudstam LG, Hansson S, Johansson S, Larsson U (1992) Dynamics of planktivory in a coastal area of the northern Baltic Sea. Mar Ecol Progr Ser 80:159–173
Slotte A (1999) Effects of fish length and condition on spawning migration in Norwegian spring spawning herring (Clupea harengus L.). Sarsia 84:111–127
Szypuła J, Grygiel W, Wyszyński M (1997) Feeding of Baltic herring and sprat in the period 1986–1996 in relation to their state and biomass. Bull Sea Fish Inst Gdynia 3:73–83
Tanasichuk RW (1997) Influence of biomass and ocean climate on the growth of Pacific herring (Clupea pallasi) from the south-west coast of Vancouver Island. Can J Fish Aquat Sci 54:2782–2788
Trippel EA, Neil SRE (2004) Maternal and seasonal differences in egg sizes and spawning activity of northwest Atlantic haddock (Melanogrammus aeglefinus) in relation to body size and condition. Can J Fish Aquat Sci 61:2097–2110
Vainikka A, Mollet F, Casini M, Gårdmark A (2009) Spatial variation in growth, condition and maturation reaction norms of the Baltic herring Clupea harengus membras. Mar Ecol Progr Ser 383:285–294
Winters GH, Wheeler JP (1994) Length-specific weight as a measure of growth success of adult Atlantic herring (Clupea harengus). Can J Fish Aquat Sci 51:1169–1179
Wood SN (2003) Thin-plate regression splines. J R Stat Soc B 65:95–114
Acknowledgments
We are grateful to the personnel at the Institute of Marine Research of Lysekil for compiling the old acoustic and biological databases. Niklas Larson and Nils Håkansson provided advice on the use of the old acoustic data. We thank Miguel Rodriguez Medina at the Baltic Nest Institute, Stockholm University, for assistance with the DAS software. Two anonymous reviewers provided very constructive comments and feedback on previous versions of the manuscript. Michele Casini was partially funded by the PLAN FISH project. We dedicate this paper to Nils Håkansson and Eberhard Götze, two of the pioneers of fisheries acoustics in the Baltic Sea, who have recently passed away.
Author information
Authors and Affiliations
Corresponding author
Additional information
Dr. V. Feldman passed away on 6 May 2007.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Casini, M., Kornilovs, G., Cardinale, M. et al. Spatial and temporal density dependence regulates the condition of central Baltic Sea clupeids: compelling evidence using an extensive international acoustic survey. Popul Ecol 53, 511–523 (2011). https://doi.org/10.1007/s10144-011-0269-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10144-011-0269-2