
Abstract
Executive functions (EFs) are a set of cognitive processes used for effortful self-regulation of behaviour. They include inhibition, working memory, cognitive flexibility and, in some models, attention. In humans, socioeconomic factors and life experiences shape development of EFs. Domestic dogs (Canis familiaris) must often regulate their behaviour in the human environment (e.g. no jumping up on humans or chasing cats), and life experiences also probably influence the development of EFs in dogs. Research into dog cognition and behaviour has been thriving, and some methods used to explore these concepts (e.g. object-choice task, questionnaires measuring traits like distraction and aggression) are likely to be sensitive to differences in EFs, even if that is not their stated aim. Here we examine relevant studies to identify experiential factors which may influence the development of EFs in dogs living in human care. These are early experience, training, housing and stress. We conclude that the development of dogs’ EFs may be negatively affected by hardships, and positively by surmountable challenges, early in life. Training methods appear important, with punishment-based methods leading to poorer dog EFs. Kennel environments seem to affect dog EFs negatively. While mild stressors might enhance the development of EFs, too much stress seems to have negative effects. Regulation of behaviour, a key outcome of EFs, is crucial for dogs’ integration into human society. We should, therefore, strive to better understand how the environment shapes dogs’ EFs.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Executive functions (EFs) is an umbrella term for cognitive processes that are used for effortful self-regulation of behaviour (Diamond 2013; Karr et al. 2018; Olsen 2018; Pecora et al. 2017). These skills are needed to keep focused on a stimulus in the face of distractions, to temporarily store useful information in memory and to adapt to changes in the environment that require flexible behaviour. EFs are needed to execute intentional behaviours that are goal-oriented, and not merely reactions to environmental stimuli. EFs are important for life outcomes in humans and known to affect behaviour (Alloway and Alloway 2010; Granvald and Marciszko 2016; Moffitt 2012). Some evidence is accumulating that cognitive traits are important for dogs’ life outcomes (Bray et al. 2017a; Cohen 2018). Experiences shape EFs in humans (Diamond and Ling 2016; Fay‐Stammbach et al. 2014). The aim of the review is to identify experiential factors that might influence the development of EFs in dogs. The review summarises articles describing studies that may assess some form of dogs’ EF capacity, either as a stated aim of the original research or through measurements that we believe to be associated with EF, in relation to life experiences (early experience, training and housing). Stress as a potential mediating factor and limitations of the results are discussed.
Importance of EFs for life outcomes in humans
In humans, EFs have been argued to be important for a myriad of life outcomes, such as mental health, physical health, and quality of life (Diamond 2013). Better-developed EF skills are linked to outcomes such as improved health, increased wealth, and reduced substance abuse and jail time (Moffitt 2012; Moffitt et al. 2011). Higher EF skills are associated with better life outcomes, even when controlling for general intelligence (Moffitt et al. 2011; Sherman and Brooks 2010). They are important for school readiness and success in the education system, job success, and marital harmony (Diamond 2013). Taken together, EFs in humans have been shown to be important for success across a wide array of areas in life.
Components of EFs in humans
Commonly identified EF components are working memory (WM), inhibition, and cognitive flexibility (CF), with attention as a potential fourth component (Diamond 2013; Garon et al. 2008; Miyake et al. 2000; Pecora et al. 2017). These components often interact with each other (Fig. 1), and it is difficult to measure them independently (Diamond 2013). Garon (2008) describes that, indeed, earlier research often viewed these components as a unitary construct, whereas later it was argued that they are dissociable components. Evidence suggests a synthesis of these approaches, with EFs in humans consisting of partly separable traits with an underlying unitary component (Miyake et al. 2000). EF skills impact individuals’ behaviours, and EF can be assessed through behaviour rating scales in human adolescents (Gioia et al. 2000) and young children (Sherman and Brooks 2010). EFs have been intensively studied in humans (Aadland et al. 2017; Aran-Filippetti and Richaud de Minzi 2012; Cepeda et al. 2001; Diamond 2013; Miyake et al. 2000) and been investigated in a variety of other species (MacLean et al. 2014) including fish and invertebrates (Cleal et al. 2020), birds, rats and monkeys (Puig et al. 2014), as well as dogs (Olsen 2018).

Relationship between core executive function skills (adapted from Diamond 2013). Working memory (WM) and inhibition are basic skills that rely on each other. Distractions must be inhibited from entering WM (i.e. attention towards distractions must be inhibited). Similarly, to be able to inhibit irrelevant information, the goal or task at hand must be kept firmly in WM (Diamond 2013). Cognitive flexibility, which builds on working memory and inhibition, is the skill enabling individuals to adjust to different environmental demands and conditions flexibly (Diamond 2013)
Working memory
While the exact structure and components of working memory are controversial, a commonly used definition describes WM as a cognitive trait enabling individuals to temporarily hold information in a state of increased accessibility (Cowan 2017). Definitions differ in whether they include a long-term memory aspect, whether processing of information is part of WM or not, and whether WM consists of one or multiple storage components (Cowan 2017). Diamond describes WM as temporarily holding information, that is no longer perceptually present, in an active state in mind while also manipulating it (Diamond 2013). It is essential for executing a task that takes any length of time, as the task objective has to be kept in WM (Diamond 2013).
Working memory (WM) has been linked to various behaviours. Deficits in working memory in children can be assessed with a behavioural rating scale (Alloway et al. 2009b). Behaviours that are associated with poor WM are poor attention and elevated levels of distractibility, and difficulties in areas such as planning and organising information (Alloway et al. 2009a, b; Gathercole et al. 2008; Lui and Tannock 2007). WM also seems to be related to aggression in children, with lower WM capacity associated with higher aggression (Granvald and Marciszko 2016). In adult police officers low WM capacity has been associated with a heightened likelihood to shoot unarmed targets and lowered likelihood to shoot armed targets (Alloway et al. 2005). Poor WM has also been linked to other problematic behaviours in adults, such as alcohol abuse (Finn and Hall 2004; Houben et al. 2011).
Inhibition and attention
Inhibition is needed to control pre-potent behavioural responses (behavioural inhibition) and to control thoughts and attention (interference control/cognitive inhibition, Diamond 2013). It is required when a well-practiced, previously rewarded or prepotent behaviour or thought is inappropriate for the situation, and a novel or less-practiced behaviour or thought is more suitable. Inhibition and WM are closely linked to each other and are argued to always be used in conjunction (Diamond 2013). WM is needed to keep information actively in mind and manipulate it. Distractions must be inhibited from entering WM (i.e. attention towards distractions must be inhibited). Similarly, to be able to inhibit irrelevant information, the goal or task at hand has to be kept firmly in WM (Diamond 2013).
Attention is the ability to focus on one stimulus or task in the environment and is crucial for any goal-directed behaviour. In a review, Garon and colleagues (2008) explain that it is argued attention is the basis for the development of other EF abilities, and that attention is an underlying factor responsible for observed correlations between different EF skills. Other models see attention as part of inhibition, specifically interference control (Diamond 2013). It is argued that to be attentive to a stimulus, a subject must cognitively inhibit the focus of attention being distracted by other aspects of the environment. Staying focused on an objective or thought draws on WM, as the focused-on objective has to be kept active in WM (Diamond 2013). In this review, we treat attention as a part of inhibition (Diamond 2013). It is difficult to disentangle components of EF even in specifically designed studies (Miyake et al. 2000), and the only study we are aware of in dogs that measured both inhibition in a motor inhibition task and attention, as rated by owners, found an association between the two (Bunford et al. 2019).
Like WM, inhibition has been linked to aggression. Children with low levels of behavioural inhibition are more likely to develop aggression, particularly unprovoked aggression than children with higher levels of inhibition (Kimonis et al. 2006; Poland et al. 2016; Raaijmakers et al. 2008). Inhibition was found to be related to social and reactive types of aggression, with lower inhibition being linked with higher levels of aggression in these (Granvald and Marciszko 2016). There seems to be a relationship between inhibition, attention-deficit/hyperactivity disorder (ADHD) and aggression. Gathercole and colleagues (2008) explain that inhibition deficiencies are believed to be a key element of ADHD, and aggression is a regular feature observed in patients with ADHD. Relationships between aggression and inhibition have not only been found in humans but also in mice (Sallinen et al. 1998) and rats (Fone and Porkess 2008; Homberg et al. 2007).
Inhibition has also been found to be associated with emotional regulation. Emotional regulation is the initiation, inhibition and modulation of emotions and accompanying behaviours (Eisenberg and Spinrad 2004). Deficits in inhibition have been linked to reduced capacity for emotional regulation (Leen-Feldner et al. 2004; Li and Sinha 2008; Reese et al. 2015; Willie 2011). This association has been found for the regulation of positive and negative emotions (Carlson and Wang 2007).
Cognitive flexibility
Cognitive flexibility is the skill enabling individuals to adjust to different environmental demands and conditions flexibly (Diamond 2013). CF is needed in daily life. For instance, when cooking a recipe and realising an ingredient is missing, trying to replace the ingredient with a suitable substitute that is available is an example of CF. CF relies on inhibition and WM. To be able to adjust or change strategies, the previous thoughts have to be inhibited, and new information has to be loaded into WM (Cepeda et al. 2001; Diamond 2013). In the cooking example, it would mean inhibiting focus on the missing ingredient and activating the substitute ingredient into WM. Because of this dependency, CF is thought to develop later in life than WM and inhibition (Garon et al. 2008). CF, or lack thereof, has been associated with psychological disorders such as obsessive–compulsive disorder (OCD) (Meiran et al. 2011), autism spectrum disorder (ASD) (Lawson et al. 2015) and related restricted, repetitive symptoms (Lopez et al. 2005).
Experiences influencing the development of EFs in humans
As outlined above, EFs are linked to behaviours and life outcomes in a myriad of ways. But what factors influence the development of EFs? In addition to genetic factors (Bowirrat et al. 2012; Eisenberg and Berman 2010; Logue and Gould 2014), experiences influence the development of EFs (Aran-Filippetti and Richaud de Minzi 2012; Diamond and Ling 2016). Parenting factors such as supporting individuals in developing their own thoughts and solutions through verbal or physical guidance (i.e. scaffolding) and offering enriched interactions (i.e. stimulation) influence EF skills in children positively (Fay‐Stammbach et al. 2014; Hughes and Devine 2019). Socioeconomic status and housing conditions (Aran-Filippetti and Richaud de Minzi 2012; Welsh et al. 2018), and time spent in prison (Meijers et al. 2015) can have an impact on EFs. Furthermore, EFs can be positively influenced by a variety of training programs and interventions (Diamond and Ling 2016). Cognitive training programs (Au et al. 2015; von Bastian and Oberauer 2013) and physical exercise (Aadland et al. 2017; Audiffren and André 2015) have been examined, with mixed results (for a review see Diamond and Ling 2016; Hillman et al. 2019). Diamond and Ling (2016) point out that one robust finding seems to be that individuals with the lowest baseline EFs benefit the most from any kind of intervention aimed at improving EFs. Experiences can affect the development of EFs, but also the expression of EFs. For example, sleep deprivation can negatively affect the expression of EFs (Tucker et al. 2010).
The importance of executive functions in dogs
We have summarised that EFs in humans appear to be important for behaviour control and life success and that they are shaped by experiences throughout life. Might this effect of experience on EF also pertain to dogs? Being able to control one’s behaviour in an intentional way, and not just reacting to stimuli as they appear, is crucial for dogs’ success in the modern human environment. While a large number of dogs (Canis familiaris) worldwide are feral (Boitani and Ciucci 1995; Hughes and Macdonald 2013), many dogs live in very close proximity to and interact regularly with, humans. These dogs live in laboratories, in shelters, as pets and working dogs. Working roles range from scent detection for explosives or medical conditions, to military and police dogs, and assistance dogs for people with psychiatric, intellectual, and physical disabilities (Bray et al. 2017b; Cobb et al. 2015; Cohen 2018; MacLean and Bray 2019; Rooney et al. 2009; Troisi et al. 2019). Even stray and free-roaming dogs often live close to human societies (Boitani and Ciucci 1995; Hughes and Macdonald 2013), and might regularly encounter humans at various times in their lives.
The different roles filled by dogs in human societies demand very different things from dogs. Some dogs are expected to be able to work or live quite independently, while others must work or live very closely with their handler. Some dogs are required to be calm, while others should be highly energetic. Despite different demands on dogs, one thing they all have in common is that the dog needs to effortfully self-regulate its behaviour. This need for regulation is true whether it is a pet dog that needs to inhibit chewing on furniture, a scent detection dog that needs to keep the target scent in mind while sniffing an array of different odours, or a guide dog that needs to keep current commands in mind while navigating the environment and ignoring distractions. For dogs living within the human environment, behavioural regulation seems crucial for a harmonious life.
It seems very likely that EFs used for the regulation of behaviour are important for dogs’ success as pets or in working roles. There is some evidence that working dogs’ performance is influenced by cognitive skills (Bray et al. 2017a, b; Cohen 2018; MacLean and Bray 2019), but only recently has emphasis been placed on measuring EF skills, per se, in dogs (Olsen 2018). More research into how EF skills present and develop in dogs, and how they influence dogs’ success in pet and working roles, will be valuable.
Influences of experiences on the development of EFs in dogs
Considering the influence experiences have on the development of EFs in humans, a question arises: how do experiences shape the development of EFs in dogs? Knowing the influences that shape a dog’s EFs throughout its lifetime may enable us to develop efficient interventions and training practices to assist each dog to reach his or her full EF capacity. This might lead to more harmonious lives of families with pet dogs, fewer dogs being surrendered into shelters, and fewer working dogs being released from their training programmes. Better outcomes for pet and working dogs would be beneficial to the welfare of dogs and humans alike.
Studies have shown that aged dogs experience a decline in EF skills, and that environmental enrichment, such as exercise, play, social contact, and diet can influence individuals’ EF skills in aging dogs. This phenomenon has been intensively studied and reviewed (Adams et al. 2000a, b; Chapagain et al. 2018a, b; Cotman and Head 2008; Cotman et al. 2002; Davis et al. 2017; de Rivera et al. 2005; Fahnestock et al. 2012; Head et al. 2012). However, much less is known about how experiences during the dog’s puppyhood, adolescence and early adult life shape its EFs.
Aim and scope: influence of experiences on the development of EF in dogs
The aim in this study was to identify and synthesize available knowledge of experiential influences on the development of EFs in young to adult dogs. Even though studies specifically examining EFs in dogs are only beginning to emerge (Olsen 2018), research into broader aspects of dog cognition has been thriving for decades (Bensky et al. 2013; Miklósi 2014). While not specifically aiming to measure EFs, some tasks used in these studies appear to be sensitive to one or more of the above-described EF skills in dogs. Hence, it is timely to review these studies in accordance with the framework provided by EFs. Because the studies in question are very rarely identified in connection with EFs, we did not consider it appropriate to conduct a formal, systematic review using the strict guidelines associated with an approach like PRISMA (Moher 2009). Instead, a narrative approach was employed to ensure the widest possible coverage of relevant literature.
Method
A literature search was carried out in the PsychInfo and ProQuest databases in January 2021. The search was based on three concepts: (a) “dogs” and related terms, (b) “executive function” and related terms, and (c) “experience” and related terms (Table 1). Keywords within each concept were combined with “or”, meaning that at least one of the keywords needed to be present in selected publications. The concepts were combined with “and”, so that only publications containing all three concepts were selected.
The search yielded 605 studies whose abstracts were then screened for relevance. Studies were included if they measured some aspect of puppy or adult dog behaviour or cognition, as well as some experiential influence that was measured simultaneously. Studies on aging dogs were excluded, as well as studies that only measured aspects of cognition or behaviour without influences of experiences and vice versa. 29 studies were retained with these criteria.
Results
The results of the review are summarised in Tables 2 and 3, which show studies grouped by whether they measured potentially corresponding behavioural traits (Table 2) or behavioural tests (Table 3).
Potential experiential influences on EFs in dogs
Early experience
Early life experience is known to be important for physical, behavioural and cognitive development (Bray et al. 2017a; Hughes and Devine 2019; Williams et al. 2009). From studies investigating influences on behaviour and general cognition, it is possible to extract information about how early experience may affect the development of EFs in dogs.
Negative effects of early hardships
Shelter puppies
Zaine et al. (2015) compared interspecies social cognition (i.e. dogs’ ability to follow human pointing cues) in puppies that were living with families as pet dogs and puppies that were housed in a shelter. They exposed the puppies to object-choice tasks with three different pointing cues: a dynamic proximal point, a momentary proximal point, and a momentary distal point. While all of these have been used extensively in dogs to measure social cognition (Clark et al. 2019), two of them appear to also measure working memory. In the momentary pointing conditions, the human raises his/her arm in the direction of an object that contains food, either at a distance (distal point) or close to the object (proximal point). In both cases, the arm is then lowered back into a neutral position before the puppy is released. Thus, the cue is not perceptually present while the puppy approaches the objects. In contrast, in the dynamic pointing cue, the human maintains the pointing position throughout the trial. The cue is perceptually present while the dog approaches the objects. During momentary point trials, the puppy must keep the direction the human pointed towards in its WM when deciding between the objects.
Puppies that lived in human homes before entering the shelter outperformed puppies found as strays in momentary pointing conditions, which might rely on WM (Zaine et al. 2015). The authors concluded that exposure to human social cues resulted in improved performance in the non-stray puppies. Pet puppies are likely to be more comfortable around humans, affecting their motivational state when participating in the task. In addition, improved performance might rely on WM in the context of social communication, and living in a home might have allowed for better WM development. Less developed WM in stray pups is further supported by the fact that stray pups easily followed the human social cue which did not rely so heavily on WM (dynamic proximal point).
It could be argued that the object-choice task has little demand on WM, as dogs can choose the object right after the disappearance of the pointing cue. The only difference between the dynamic and momentary cues is whether the cue is perceptually present during decision-making. To be successful in the dynamic conditions, dogs could use trial and error learning or lower-level strategies, such as local enhancement, or they might understand human communicative intent. However, in the momentary condition, dogs must remember the current location of the food reward without a visual cue being present. Therefore, the current food location could be kept active in WM. Dogs might also use other cognitive strategies related to EF, such as focusing their attention onto the correct location throughout the trial. Further research comparing performance in the pointing task with added delays and comparison to WM tasks without a human/social component will be valuable.
Breeding facilities
Puppies bred in high-volume commercial breeding establishments and/or sold through pet stores have been reported to experience increased behavioural problems compared to puppies that were bred by smaller non-commercial breeders (McMillan 2017). As outlined in the introduction, aggression has been found to be associated with inhibition in humans, mice and rats (Fone and Porkess 2008; Homberg et al. 2007; Kimonis et al. 2006; Poland et al. 2016; Sallinen et al. 1998).
A common problem found in dogs from pet stores and commercial breeding facilities is aggression towards household members (Bennett and Rohlf 2007; Casey et al. 2014) and owner-directed aggression (Pirrone et al. 2016). It is likely that aggression is caused by fear towards humans due to a lack of socialisation. McMillan (2017) points out that the ways in which puppies are bred, housed, weaned, transported, handled and homed can all influence their behavioural development. Given the link between aggression and inhibition in other species (Fone and Porkess 2008; Homberg et al. 2007; Kimonis et al. 2006; Poland et al. 2016; Sallinen et al. 1998), an additional possible explanation might be a low inhibition due to poor conditions during sensitive periods of early cognitive development. However, to our knowledge, no studies have directly tried to examine a possible link between inhibition and aggression in dogs, and future research on this area will be valuable.
McMillan and colleagues (2013) compared dogs that were acquired as puppies, either from pet stores or from non-commercial breeders, on behavioural measures using the C-BARQ (Hsu and Serpell 2003). Pet store puppies scored higher on a variety of behaviours, namely being more aggressive, excitable, energetic, and attached, as well as less trainable. As outlined earlier, regulation of emotions and accompanying behaviours is associated with inhibition in humans (Carlson and Wang 2007; Leen-Feldner et al. 2004; Li and Sinha 2008; Reese et al. 2015; Willie 2011). A link between emotion regulation and inhibition might be true in dogs as well. If a dog becomes overly excited by regular occurrences, such as seeing another dog or greeting a person, the dog might be failing to regulate its emotions and, by extension, these behaviours. This is supported by studies in dogs showing that dogs that are more impulsive as rated by their owners exhibit lowered inhibition in cognitive tasks (Brady et al. 2018; Bunford et al. 2019; Wright et al. 2012). Arousal has been shown to influence inhibition in dogs (Bray et al. 2015), and more behavioural studies examining a possible link between inhibition and excitability in dogs will be valuable.
In sum, studies looking at the impact of commercial breeding establishments on later dog behaviour found an increased risk of aggression (McMillan 2017). As with the strays from Zaine’s (2015) study, puppies that experienced fewer human interactions might have reduced EF skills. Whether this was due to the rearing environment is difficult to determine, since there might also be an inherent difference in how owners that purchase puppies from commercial breeding establishments care for and train their dogs, compared to owners that purchase from small, non-commercial breeders, which could also influence adult dog behaviour. Further research is required to investigate and disentangle these possible effects.
Effects of early isolation
In the beginning of dog behaviour research in the 1950s and 1960 s, two studies investigated the effect of isolation of young puppies on later dog cognition. Isolation-reared beagle puppies were compared to puppies that received some human contact and exercise twice daily (Fuller 1966). Both groups were placed in isolation cages at 3 weeks of age, meaning that both were weaned early and deprived of maternal and littermate contact. The puppies were tested on a reversal-learning task starting when they were 23–26 weeks old. First, puppies were trained to remove a cover from food bowls in which dog food was hidden. Two food bowls were presented simultaneously, both baited and with removable covers. After establishing the puppies’ preferred side, the cover on the preferred side was locked in position, making the food unreachable for the puppies, while the opposite side cover remained removable. A correct response was measured if the puppy obtained food from the accessible food bowl (Fuller 1966). To be successful, the puppies had to inhibit their previously rewarded response of approaching their preferred food bowl, and flexibly adjust their behaviour to approach the non-preferred food bowl. The puppies reared with some human contact made fewer errors, especially in the first reversal series. Some stimulation, compared to no stimulation during early life, may have enhanced the development of EFs, such as inhibition and CF.
Similarly, Thompson & Heron (1954) compared isolation-reared puppies to puppies reared as pets or in the laboratory with more human contact and stimulation, on a series of problem-solving tasks when they were between 7 and 12 months old. In one task, puppies learned the location of a bowl with food, which was then moved to another position in the puppies’ full view. Puppies were then released and allowed to run towards the food. To be successful, puppies had to inhibit running towards the previously rewarded location, and flexibly adjust their behaviour to run to the new location with the food bowl. Pet-reared puppies outperformed the isolation-reared puppies, with pet-reared puppies making almost no mistakes whereas isolation-reared puppies failed on more than half of the trials. Similar results were obtained for detour tests around a barrier. Additionally, a delayed-choice task was administered, in which puppies were trained in choosing one of two locations which they had seen being baited with food. A delay was then introduced before the dogs could make their choice, ranging from 5 to 300 s. To succeed at this task, dogs must keep the location of the food bowl in their working memory when deciding which bowl to approach. The groups differed significantly, with isolation-reared puppies succeeding with delays of only 0–25 s, whereas pet-reared puppies succeeded with delays of 50–300 s. Overall, pet-reared puppies outperformed isolation-reared puppies on tasks that required inhibition, CF and WM.
Positive effects of experiences early in life
While early hardships potentially have detrimental effects on EFs, mild challenges and enrichment might foster EF development. Some evidence comes from studies with guide dog candidates. One of the studies that investigated early life experience and how that affects later dog behaviour implemented a standardised socialisation program (Vaterlaws-Whiteside and Hartmann 2017). The study was done with litters of a guide dog population in addition to their normal socialisation routine. Puppies were exposed to daily interactions with a handler, aiming at tactile, auditory and visual stimulation, and had interactions with people and the environment. The daily stimulation was developed to be age appropriate, increasing in intensity as the puppy developed from birth to 6 weeks. For example, in the first week the interaction with people consisted of picking up the puppy and carrying it around the kennel, whereas in week 6 the puppy was gently restrained for 20 s. Many of the stimuli used, like rolling balls within the puppy’s reach and hanging tinsel above the puppy, would have encouraged the use of focused attention towards stimuli in the environment (Vaterlaws-Whiteside and Hartmann 2017).
The effects of the programme were evaluated with a Puppy Profiling Assessment (PPA) at 6 weeks, which measures responsiveness to human or environmental stimuli (Asher et al. 2013) and with a Puppy Walker Questionnaire (PWQ, Harvey et al. 2016) when puppies were 8 months old (Vaterlaws-Whiteside and Hartmann 2017). The PWQ looks at different behavioural traits, two of which might be relevant for EF assessment: distractibility and excitability. Items in the questionnaire aiming to determine distractibility asked for the dogs’ behaviours when encountering distractions such as children, other dogs, food on the floor and whether the dog pulled towards distractors when on leash. Puppies that received the extra socialisation scored more favourably on the PPA and had significantly more favourable distraction scores and a trend for better excitability scores on the PWQ. Less distractibility and excitability could be mediated by improved inhibition in comparison to puppies that did not receive the additional socialisation. As the dogs in this study were guide dog candidates, they were undergoing training to inhibit impulses towards distractors. Being distracted by food, other dogs or members of the public shows a lack of behavioural inhibition. The gentle introduction of stimuli throughout the first weeks of life might have challenged the puppy while not being overwhelming, enhancing the development of EFs.
Bray et al. (2017a) followed a population of guide dogs from birth to programme completion (i.e. either placement as a guide dog or failure to be placed due to behavioural issues). They observed maternal behaviour during the first 3 weeks of life, recording levels of overall maternal behaviour and nursing styles. Dogs can nurse in varying positions which might differ in their difficulty for the puppy to stay attached to the nipples, two of which are nursing while standing and laying down. Vertical nursing is more challenging for puppies, with more effort required to attach to the nipples and remain attached. Maternal behaviours like time spent in contact with the puppies and licking and grooming of puppies differed between litters, as did the time the bitches spent nursing in a lateral position (laying on their side) or a vertical position (standing or sitting).
When testing the same puppies as young adults, using a multistep problem-solving task, dogs learned to remove bones and to spin an apparatus to expose hidden treats (the Nina Ottoson Dog Tornado game; Bray et al. 2017a). To get to the food reward, the bone needs to be removed before the food can be uncovered. Perseveration was measured as the time the dog manipulated the area under which the treat was hidden, while the access was still blocked by the bone. Pre-training consisted of dogs having to remove bones, and all dogs could do this. Therefore, next to physical reasoning and persistence, perseveration as described in the study can be seen as a measure of inhibition. To get to the food, dogs need to divert their attention away from the food and to the bone first. Manipulating the area that is baited while the bone is still in place may show a lack of inhibition.
More maternal care during early development was associated with more perseveration (Bray et al. 2017a). Lowered inhibition might, therefore, be linked with increased maternal care. The authors argued that puppies in this population all received sufficient maternal care not to be disadvantaged, but that higher amounts of maternal care may mean that the puppies were less challenged. This link is in line with Vaterlaws-Whiteside’s study (2017), where challenges during early life likely enhanced the development of inhibition.
Early experience seems to be important for the development of EF in dogs. Hardships in the sensitive period of the first weeks of life might negatively affect the development of EFs. Potential evidence comes from stray puppies (Zaine et al. 2015) with a reduced WM duration, in puppies from commercial breeding establishments, with increased aggression and excitability (McMillan 2017), potentially linked to lowered inhibition, and in isolation-reared laboratory puppies with reduced inhibition, CF and WM (Fuller 1966; Thompson and Heron 1954). On the other hand, small age-appropriate challenges and enrichment appear to foster the development of EF. These small challenges can be presented through human interaction (Vaterlaws-Whiteside and Hartmann 2017) or be imposed by amount and type of maternal care (Bray et al. 2017a).
Training
Dogs experience a wide range of training during their development. Some dogs kept as pets never experience any formal training, some get obedience training ranging from basic commands to a variety of elaborate tricks, and some train for agility or scent tracking with their owners. Working dogs undergo meticulous and standardised training procedures which will qualify them as scent detection, military or assistance dogs, to name just a few. During basic obedience, or in more elaborate working roles, dogs must remember commands, block out distractions, and adjust flexibly to changing circumstances. Using these EF skills during training is likely to affect EF development.
Effects of training amount and type
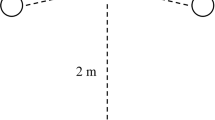
Training seems to have positive effects on inhibition, with multiple studies demonstrating a relationship between training level and inhibition, measured in behavioural tasks (Barrera et al. 2018; Fagnani et al. 2016; Marshall-Pescini et al. 2016) as well as via owner ratings (Vas et al. 2007). One of the behavioural measures used is the A-not-B task, which stems from early studies on object permanence in human children (Piaget 1954). It has been used in a variety of contexts with a variety of species including lizards (Szabo et al. 2019), alpacas (Abramson et al. 2018), marmosets (Kis et al. 2012), dogs (Barrera et al. 2018; Bray et al. 2014; Kelly et al. 2019) and cross-species comparisons (MacLean et al. 2014). During the task, the subject sees one cup/location (location A), out of two or more, being repeatedly baited with food or a toy, and the subject can then retrieve the food/toy from there. After a specified number of trials, the reward is placed in location A but then switched to a different cup/location (location B) in full view of the subject. The subject is then allowed to retrieve the reward from location B (Fig. 2). If the subject approaches the previously rewarded location A instead of location B, a so-called A-not-B error is committed. Traditionally it has been argued that the A-not-B task is a measure of inhibition, as the subject must inhibit the previously rewarded and, therefore. pre-potent, motor response to retrieve the reward from cup B (Diamond 1985; Marcovitch and Zelazo 1999). It is also a form of reversal learning, in which the subject must switch response strategies, which is a measure of CF. Furthermore, even though the subject can see the location being baited, the reward is typically out of sight (i.e. usually underneath a cup) when the subject makes its decision to approach, so it must also use WM to keep the current reward location in mind (Espy et al. 1999) or via allocating attentional resources. All these are components of EFs.

A-not-B task. During A-trials, the dog sees location A being baited and is then released to make a choice for one of the locations. During B trials, the dog sees location A being baited. The bait is then removed, in full view of the dog, from location A and moved to location B instead before the dog is released to make a choice for one of the locations
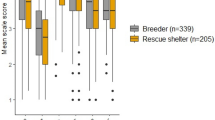
In a study designed to investigate EF in dogs, Barrera et al. (2018) used the A-not-B task to compare performance of water rescue dogs to pet dogs. The water rescue dogs used in this study were not from a specific breeding stock, but, rather, were “family dogs that learn a specific social work” (Barrera et al. 2018), which made it possible to look at the effect of training, without the interference of other components such as heritability. Water rescue dogs committed fewer A-not-B errors than pet dogs. The authors concluded that water rescue dogs exhibited better inhibition than pet dogs and that the training these water rescue dogs went through during their development provided more opportunities to improve their inhibition. They compared their result to a study looking at performance in the A-not-B task in pet dogs compared to shelter dogs (Fagnani et al. 2016). Pet dogs outperformed shelter dogs in the A-not-B task. It is likely that shelter dogs are less comfortable around humans, as they have limited human interaction. Limited human contact might affect their motivation to participate. Training differences between the two groups are another potential influence.
Another behavioural task that aims to measure inhibition is the detour task. In a study on problem-solving ability in dogs of different training backgrounds and breed groups, Marshall-Pescini and colleagues (2016) used a detour fence task. A v-shaped fence made from wire mesh was placed in a room, with a toy placed at the inner side of the intersecting angle. The toy was pointed out to the dog through the wire mesh before it was brought to a starting point two metres away from the fence. The dog was released and had one minute to reach the toy. To get to the toy, the dog needed to inhibit going straight towards it, instead moving in a different direction first. The subjects for this study were divided by breed group as well as training status, whereby trained meant competing in a sports discipline or certified working dogs and untrained meant no training or basic obedience. Most (71%; 42 out of 59) trained dogs successfully detoured within one minute and reached the toy, whereas only 45% (29 out of 64) of the untrained dogs were successful. When comparing only successful dogs, trained dogs were faster to reach the reward than untrained dogs. Trained dogs were exhibiting greater behavioural inhibition.
Next to behavioural tasks measuring inhibition, owner-rated aspects of inhibition have been shown to be related to the level of training dogs had received. Vas and colleagues (2007) developed a scale to measure attention deficit hyperactivity disorder (ADHD) like behaviours in dogs, using an established human ADHD questionnaire. In humans ADHD is known to be accompanied by an EF deficit (Eakin et al. 2004; Martel et al. 2007; Toplak et al. 2008). Using factor analysis of the 13 items that were included in the questionnaire two main factors emerged, which the authors named ‘inattention’ and ‘activity-impulsivity’. It can be argued that this questionnaire partly measures EF skills. The ‘activity-impulsivity’ factor is likely to measure aspects of inhibition, as impulsive actions like lunging and pulling on the lead show a failure to inhibit motor responses when they are not appropriate. Additionally, as outlined above in humans ADHD is closely linked to deficits in inhibition (Nigg 2001). Similarly, the ‘inattention’ factor measures attention which is seen as a component of inhibition (Diamond 2013). To keep attention focused on a task or cue, diverting attention towards distractions in the environment must be inhibited.
The study (Vas et al. 2007) looked at the effect of training history on the dogs’ ADHD-like behaviour scores. Subjects were divided into three training groups—those that had participated in a special training course (i.e. advanced), basic training course (i.e. beginners) or were untrained or trained at home (i.e. untrained). The ‘activity-impulsivity’ score was not affected by the dogs’ training level, but the ‘inattention’ score was associated with the level of training. Beginners and advanced dogs, who had experienced structured training, had lower ‘inattention’ scores than dogs that were not trained. It appears that training influenced inhibition in some contexts, but not in others. This difference between measures of inhibition is in line with other research showing that dogs’ inhibition responses in different tasks are not correlated, so inhibition appears to be context dependent (Bray et al. 2014; Vernouillet et al. 2018).
A set of studies investigated attention and inhibition with behavioural tests, regarding level of training in the dogs. Chapagain and colleagues (2017) first calculated pet dogs’ (110 dogs of various breeds and 75 Border Collies), training score. Owners answered questions about the dog’s experience with 13 different types of training including basic obedience and working dog training (e.g. puppy school, obedience, service dog training). To measure attentional capture and sustained attention, a toy was attached to the ceiling and could be moved with a string. The dog’s latency to look at the moving stimulus, as well as the duration of looking throughout the one-minute trial, were measured. In a social version of the task, an experimenter moved their arm up and down vertically, facing away from the dog to not establish eye contact, and the dog’s looking behaviours were recorded. Dogs with a higher training score looked at both stimuli, the social and non-social one, for longer in comparison to dogs with a lower training score (Chapagain et al. 2017). Sustaining attention to a stimulus is closely linked to cognitive inhibition, as diverting attention to other stimuli must be inhibited.
In the same study, dogs’ selective attention was measured using a task in which dogs had to shift their attention from food that was being dropped to the floor to making eye contact with the experimenter (Chapagain et al. 2017). When eye contact was established the experimenter marked it with a clicker and dropped another piece of food to the floor. To be successful at this task dogs need to selectively switch their attention from the floor to the experimenter’s eyes while inhibiting searching the floor for food when eye contact was being established. This switching relies on CF and inhibition. A higher training score and experience with clicker training were associated with a shorter latency to re-establish eye contact, which means that dogs that experienced more training throughout life were better at inhibiting searching for food on the floor and more quickly shifted their attention towards the experimenter. Border Collies and other dog breeds only differed in their latency to find food, not in any other measure of attention captured in this study (Chapagain et al. 2017). However, in a study using the same tasks in a group of border collies, with the same assessment of lifelong training score, no effect of training on any of the tasks was found (Wallis et al. 2014). Wallis and colleagues reported that the dogs in their study had participated in five different types of training on average. Hence, it is possible that the high training score masked any potential effects of training on performance.
Mongillo and colleagues (2016) examined the effects of training on attention using a ‘stay’ exercise in three different conditions. During the task, dog owners were asked to have their dogs stay in a desired posture for 30 s, stay in the presence of food for 30 s, and stay in the presence of a toy for 30 s. Outcome measures were the time the dog stayed in the desired position, therefore, keeping the command to ‘stay’ active in WM and inhibiting the impulse to break the ‘stay’ command, as well as the time spent looking at the owner, thereby inhibiting the impulse to look at the distractor. Subjects in this study were divided into three groups of different training levels. ‘Novice’ dogs had received no professional training, ‘intermediate’ dogs were receiving obedience training with a professional trainer at the time of testing, and ‘expert’ dogs had completed an obedience course with a professional trainer. The amount of time dogs held the stay command in the no-distractor condition, meaning they kept the command in working memory and inhibited moving, was longer for expert than novice dogs, whereas the maintenance time was not significantly affected by training level in the food or the toy condition. When comparing the time dogs looked at their owner, expert dogs looked longer towards their owner than intermediate dogs, which looked longer at their owner than novice dogs. All dogs looked longer at their owner in the condition without food or toy, than with. This difference in performance suggests that higher training levels are associated with greater attention towards the owner and inhibition to look toward the distractor. Similarly, a large-scale citizen study (Horschler et al. 2019) found that training history as reported by the owner was significantly positively related to an experimental measure of inhibition. The owner placed a treat in front of the dog with a verbal command for the dog not to take the treat. The latency for the dog to take the treat is a measure of inhibition. Dogs that received more training took longer to take the treat.
Inhibition seems to be positively influenced by training, with multiple studies demonstrating a relationship between training level and inhibition, measured in behavioural tasks (Barrera et al. 2018; Chapagain et al. 2017; Fagnani et al. 2016; Horschler et al. 2019; Marshall-Pescini et al. 2016; Mongillo et al. 2016) as well as via owner ratings (Vas et al. 2007). However, Wallis and colleagues (Wallis et al. 2014) did not find an effect of training. Additionally, looking at other EF skills, no effect of training on performance on an object discrimination task was found (Chapagain et al. 2018b). The dog’s training score was calculated in the same way as in the previous studies, and the task included a touch-screen apparatus, in which the dog needed to distinguish between positive and negative stimuli. Dogs had to remember the correct stimulus from the last trial when making a decision, which likely relies on WM. When switching from a wrong response to a different response option, dogs likely had to rely on CF. Dogs were trained on the task over multiple sessions, so it is possible that initial differences in WM and CF might have been masked. In a study comparing pet dogs and detection dogs on an object-choice task with momentary cues, no difference in performance was found (Lazarowski et al. 2020). Furthermore, Horschler and colleagues (2019) also used a delayed memory task in their study, which is a measure of WM. No effect of training on performance was found.
Overall, training might have a positive effect on dogs’ inhibition, but the results are not clear. Less is known of the effects on CF and WM. More studies are needed that specifically aim to assess the relationship between a dog’s training experience and EF skills.
Effects of training method
Dog training methods might influence a dog’s EFs differently. Methods range from reward-based approaches to aversive-based approaches. Reward-based approaches aim at increasing desired behaviours by presenting pleasant stimuli when the behaviours are executed; stimuli used are normally food or praise. Conversely, pleasant stimuli are withheld if the dog executes undesired behaviours. Aversive-based approaches, on the other hand, deliver unpleasant stimuli, such as scolding or physical pain, upon execution of undesired behaviours. Training methods sometimes combine these approaches (Fernandes et al. 2017). Numerous studies have shown increased aggression in dogs trained with aversive-based training methods (Ziv 2017), but some methodological issues are pointed out by Fernandes and colleagues (2017).
Studies using owner reports of training methods and dog behaviour showed higher aggression and/or excitability with more punishment used (Arhant et al. 2010; Casey et al. 2014) and lower aggression and/or over-excitement associated with reward-based methods (Herron et al. 2009; Hiby et al. 2004), although the direction of causality cannot be determined. Again, these studies do not focus specifically on dog EFs, but excitability might be a measure of inhibition, as dogs fail to inhibit inappropriate motor responses such as jumping up or pulling on the lead. Additionally, aggression has been shown to be linked to lowered inhibition in humans and other species (Fone and Porkess 2008; Homberg et al. 2007; Kimonis et al. 2006; Poland et al. 2016; Sallinen et al. 1998). Future studies should directly examine the effects of training methods on dog cognition and EFs.
Haverbeke and colleagues (2008) looked at team performance in military working dogs and their handlers. They assessed the amount of reward- and aversive-based methods the handler used in an evaluation using obedience and protection work exercises. The distraction of the dog was measured by their head and body orientation, with head/body orientation towards the handler indicating focus, and orientation in a different direction for more than 1 s considered to be indicative of distraction. Distracted dogs showed lower performance on the tasks, and lower performing dogs received more aversive stimuli. The direction of causality here is not clear, but it is possible that more aversive stimuli led to dogs being more distracted and hence performing worse. To keep the focus on the task and the handler, the dog needs to inhibit paying attention to distracting stimuli in the environment. The dog’s inhibition might have been compromised by receiving aversive stimuli. Future studies should aim at examining the relationship between aversive based training methods and dogs’ EF skills.
It appears that aversive-based training methods may impact dogs’ EFs, increasing aggression, distraction and excitability (i.e. lowering inhibition), although it will be necessary to investigate dogs’ EFs more directly when comparing them to the training method used. Ratings of dog behaviour by owners that use aversive or reward-based training methods might be biased towards their own preferred method of training. Administering laboratory tests designed to measure behaviours associated with inhibition (such as the A-not-B), WM (such as a delayed response test) and CF (such as reversal learning tests) to compare those to owner-reported training methods will be more informative.
Housing
Dogs’ housing conditions vary immensely, from comfortable, environmentally enriched homes to kennels. Dogs are kept in kennels for a multitude of reasons. Animal shelters house abandoned, abused and stray animals. Assistance dogs are often housed in kennels when they are undergoing an intense training program, and other working dogs such as police or military dogs are often housed in kennels when not working. Dogs, mostly beagles, are bred and kept in laboratory kennels for research purposes. In these instances, dogs are often housed in kennels during developmentally important early stages of life. Kennel environments are known to be noisy, impose spatial restrictions and offer little environmental enrichment, and can, therefore, affect the dogs’ welfare (Gunter 2018; Polgar et al. 2019).
Udell and colleagues (2008) explored the performance of differently housed groups of dogs in the object-choice task. During the task, a momentary distal pointing cue was utilised. As described above, in this test, a human points to one of two objects for a few seconds, before the cue is removed, and the dog is then allowed to make a choice. To follow the point, the dog must keep the direction in WM. When testing pet dogs and shelter dogs under the same conditions, indoors with an unfamiliar experimenter, shelter dogs failed to perform above chance level, whereas pet dogs did. Shelter dogs were strays before living in the shelter. It is unclear how long these dogs had been in the shelter and whether their development was influenced by the shelter environment. Shelter dogs might have been affected by the human presence during the tasks, as they are less likely to being used to human interaction. However, the authors mentioned that shelter dogs readily interacted with the experimenter and ate food out of the experimenter’s hand (Udell et al. 2008). The inferior performance of the shelter dogs, therefore, cannot be attributed to an unwillingness to interact with the experimenter. A reduced WM in shelter dogs, on the other hand, could explain their performance, but whether this can be explained by shelter housing remains to be investigated.
Similarly, pet dogs outperformed laboratory dogs (Lazarowski and Dorman 2015) on an object-choice task. Dogs were tested with two point-following tasks, a momentary distal pointing cue, which requires WM, and a dynamic proximal pointing cue which is present while the dog makes its choice. The pet dogs followed the momentary pointing cue whereas laboratory dogs did not. However, laboratory dogs could follow the dynamic pointing cue. Laboratory dogs did not struggle to perform the mechanics of the task, demonstrating no deficits in motor function or motivation, but could not keep the communicated location in WM during the momentary pointing trials. The research dogs were born and raised in research kennels throughout their entire development, with some enrichment provided, such as tiered pens, play objects and daily interaction with humans for husbandry and veterinary purposes. Pet dogs had lived in a human home for at least one year. The kennel housing might, therefore, have affected the laboratory and shelter dogs’ EFs, specifically their WM measured in these tasks.
When a series of inhibition tasks was administered to sled dogs (Huskies) and pet dogs (various and mixed breeds), some differences in performance became apparent (Kelly et al. 2019). The traditional A-not-B task (Fig. 2), an A-not-B Barrier task and the Cylinder task were used in this study. As described above, the A-not-B task is a measure of EFs, specifically, behavioural inhibition and CF, during which the subject must inhibit approaching a previously rewarded location in favour of a new location. The barrier version of this task features a barrier which blocks access to a food reward, with a gap either left or right of it. Instead of the location of the food, the location of the gap is altered. The cylinder task is another classic task that aims to measure inhibition (Bray et al. 2014; Langbein 2018; MacLean et al. 2014). A food reward is placed in an opaque cylinder and subjects learn to approach the reward from the side of the cylinder. Then, a food reward is placed in a transparent, but otherwise identical, cylinder and is, therefore, visible for the subject. To get to the food, the subject needs to inhibit going straight for it but must detour to the side opening of the cylinder. In the A-not-B task, sled dogs chose the previously rewarded location more often, exhibiting a strong A-not-B error (Kelly et al. 2019). The groups did not differ in other tasks. Sled dogs had been living with the owner of the racing kennel since they were under 1 year old. They were training and racing, suggesting they were sufficiently exercised. However, they were housed in kennels when not training, individually or in pairs of two. Pet dogs were living in family homes. Kennel housing may have a negative effect on dogs’ inhibition, at least in some contexts, although many other possible explanations, like heritability and differences in breed and age, require further investigation.
When investigating the impact of housing conditions of guide dog candidates, Harvey and colleagues (2016) developed the Puppy Walker Questionnaire (PWQ) mentioned above. Amongst other things, this questionnaire measures distractibility and excitability. Both of those measures might be indicative of the dog’s inhibition. The more children that were living in the household, the more excitable and distractible the dogs were, which might be related to less inhibition in those dogs. Additionally, if the puppy walker had previous experience in raising an assistance dog, the dogs were less distractible, possibly indicating better inhibition.
Kennel housing appears to have detrimental effects on EFs of dogs living in shelters, laboratories and working dog facilities (Kelly et al. 2019; Lazarowski and Dorman 2015; Lefebvre et al. 2007; Udell et al. 2008, 2010). This effect on EF might influence adoption rates or success in a working role. Nonetheless, kennel housing for stray or working dogs cannot always be avoided, so it will be useful to design studies that investigate the effect of kennel housing on EFs in more detail. Social and non-social enrichment in kennelled dogs is well known to improve dog welfare and behaviour (Schipper et al. 2008; Wells 2004). However, to our knowledge, less is known about the effects of enrichment on kennelled dogs’ cognitive traits. Disentangling how social isolation, lack of exercise, lack of human contact and noise levels affect EFs could lead to more tailored interventions and enrichment to mitigate these effects.
Stress—a potential mediating factor
When thinking about possible common factors of experiences that are associated with reduced EF, stress comes to mind. Stress is known to affect EF skills in humans. Diamond (2013) lists negative influences on EF performance, namely stress, sadness, loneliness and sleep deprivation. When examining the influences that negatively impact the development of dogs’ EF discussed throughout this review—negative early experiences, aversive training methods, and kennel housing with lack of social or physical enrichment—stress could be the mediating factor. It is possible that all of these induce some form of stress in the developing dog. Dogs from strenuous breeding conditions show signs of poor welfare (Sonntag and Overall 2014), as do dogs trained with aversive methods (Haverbeke et al. 2008). Kennel housing is known to induce stress (Polgar et al. 2019) through factors including social isolation, lack of enrichment, and noise level (Coppola et al. 2006; Overall et al. 2019). More research is needed to investigate the relationship between environmental factors that induce stress and EF development.
Limitations and future directions
Some of the studies using behavioural tests discussed in this review that were designed to measured aspects of EF and cognition, give us a good idea about the experiential influences on EF development in dogs (Barrera et al. 2018; Bray et al. 2017a; Chapagain et al. 2018b; Fagnani et al. 2016; Fuller 1966; Horschler et al. 2019; Kelly et al. 2019; Marshall-Pescini et al. 2016; Thompson and Heron 1954). Other behavioural tests still provide valuable information, but the influence of EF skills on performance is less clear. Ontogenetic and other factors have been shown to be important for performance in the object-choice task (Oliva et al. 2015; Udell et al. 2010). However, they are likely to also measure at least some aspect of WM or attention in the context of social communication. This is especially true when comparing a dynamic cue which is perceptually present when the dog approaches the object, with a momentary cue which is no longer perceptually present when the dog approaches (Lazarowski and Dorman 2015; Lazarowski et al. 2020; Udell et al. 2008, 2010; Zaine et al. 2015). More studies are needed to investigate the experiential influences on WM more broadly, without the use of human social communication.
Some studies have shown a link between owner-rated impulsivity and cognitive tasks measuring inhibition in dogs (Brady et al. 2018; Bunford et al. 2019; Wright et al. 2012), while others did not detect such a link (Brucks et al. 2017; Mongillo et al. 2019). To our knowledge, there are no published studies whose stated aim was to examine the relationship of other behavioural traits, like aggression or excitability, to EF skills in dogs. Conclusions drawn in this review about possible relationships of these behavioural traits with EF skills must, therefore, be handled cautiously. Given the well-documented link of behavioural traits in humans (Alloway et al. 2009a; Gathercole et al. 2008; Kimonis et al. 2006; Poland et al. 2016), as well as other species (Homberg et al. 2007; Sallinen et al. 1998), to EF skills, it seems likely that other behavioural traits are also linked to EFs in dogs. Studies comparing owner-rated behavioural traits of dogs with the dogs’ performance in cognitive tests will be important to further our understanding of how EFs present in dogs.
Some studies included a comparison of behaviour in the presence of humans between dogs that experienced vastly different socialisation to humans throughout their lives (Barrera et al. 2018; Fagnani et al. 2016; Kelly et al. 2019; Lazarowski and Dorman 2015; Udell et al. 2008, 2010; Zaine et al. 2015). Dogs that are more accustomed to the presence of humans might be more comfortable and more likely to engage in the task. Dogs living as pets are more likely to be well socialised with humans, although there might be vast differences in the level of socialisation between different pet dogs. Dogs living in shelters, laboratories and as working dogs might experience a wide range of socialisation with humans. For example, some working dogs might be highly socialised (e.g. guide dogs) while other working dogs might be less socialised (e.g. military dogs). However, studies using the object-choice task comparing dogs from different populations found that less socialised dogs nevertheless succeeded in the easier version of the task, suggesting dogs were not too fearful to participate. The situation could have been perceived as stressful by the dogs and, therefore, affected expression of EFs (Lazarowski and Dorman 2015; Udell et al. 2008, 2010; Zaine et al. 2015). To clarify the effect of human presence on behaviour and performance, development of EF tasks that can be administered remotely, without the presence of humans, will be valuable.
Conclusions
A dog’s upbringing, housing and training experiences are all likely to impact EF-related task performance. Strenuous conditions without appropriate stimulation, be it during early life, housing or training, might be detrimental to dogs’ regulation of behaviour and might produce aggressive, distracted, and excitable dogs. On the other hand, small age-appropriate challenges throughout life might foster EF development. Given that regulation of behaviour, a key aspect of EFs seems to be crucial for dogs’ successful integration into human society, in pet or working roles, we should strive to better understand how the experiences we expose dogs to shapes their EF skills. Furthermore, research into interventions designed to enhance EF skills could prove beneficial for dogs with low EF skills, which are potentially caused, or exacerbated, by strenuous conditions. Increasing our understanding of what influences dog EFs may help us improve training, housing, or other practices, which can make dogs reach the full potential of their EF skills. Future studies should aim to specifically examine environmental influences on EFs. While results from studies using scales that assess ADHD-like symptoms (Vas et al. 2007) and problematic behaviours (Hsu and Serpell 2003) are useful in getting an idea about dogs’ EFs, a scale that aims to assess dogs’ full range of EF skills, not only pathological features, would be a good first step to measure EF and influences on EF on a larger scale.
References
Aadland KN, Moe VF, Aadland E, Anderssen SA, Resaland GK, Ommundsen Y (2017) Relationships between physical activity, sedentary time, aerobic fitness, motor skills and executive function and academic performance in children. Ment Health Phys Act 12:10–18
Abramson JZ, Soto D, Zapata S, Lloreda MVH (2018) Spatial perseveration error by alpacas (Vicugna pacos) in an A-not-B detour task. Anim Cogn 21:433–439
Adams B, Chan A, Callahan H, Milgram NW (2000a) The canine as a model of human cognitive aging: recent developments. Prog Neuropsychopharmacol Biol Psychiatry 24:675–692
Adams B et al (2000b) Use of a delayed non-matching to position task to model age-dependent cognitive decline in the dog. Behav Brain Res 108:47–56
Alloway TP, Alloway RG (2010) Investigating the predictive roles of working memory and IQ in academic attainment. J Exp Child Psychol 106:20–29
Alloway TP, Gathercole SE, Adams AM, Willis C, Eaglen R, Lamont E (2005) Working memory and phonological awareness as predictors of progress towards early learning goals at school entry. Br J Dev Psychol 23:417–426
Alloway TP, Gathercole SE, Kirkwood H, Elliott J (2009a) The cognitive and behavioral characteristics of children with low working memory. Child Dev 80:606–621
Alloway TP, Gathercole SE, Kirkwood H, Elliott J (2009b) The working memory rating scale: a classroom-based behavioral assessment of working memory. Learn Ind Differ 19:242–245
Aran-Filippetti V, Richaud de Minzi MC (2012) A structural analysis of executive functions and socioeconomic status in school-age children: cognitive factors as effect mediators. J Genet Psychol 173:393–416
Arhant C, Bubna-Littitz H, Bartels A, Futschik A, Troxler J (2010) Behaviour of smaller and larger dogs: effects of training methods, inconsistency of owner behaviour and level of engagement in activities with the dog. Appl Anim Behav Sci 123:131–142
Asher L et al (2013) A standardized behavior test for potential guide dog puppies: methods and association with subsequent success in guide dog training. J Vet Behav Clin Appl Res 8:431–438. https://doi.org/10.1016/j.jveb.2013.08.004
Au J, Sheehan E, Tsai N, Duncan GJ, Buschkuehl M, Jaeggi SM (2015) Improving fluid intelligence with training on working memory: a meta-analysis. Psychon Bull Rev 22:366–377. https://doi.org/10.3758/s13423-014-0699-x
Audiffren M, André N (2015) The strength model of self-control revisited: linking acute and chronic effects of exercise on executive functions. J Sport Health Sci 4:30–46
Barrera G, Alterisio A, Scandurra A, Bentosela M, D’Aniello B (2018) Training improves inhibitory control in water rescue dogs. Anim Cogn 22:127–131
Bennett PC, Rohlf VI (2007) Owner-companion dog interactions: relationships between demographic variables, potentially problematic behaviours, training engagement and shared activities. Appl Anim Behav Sci 102:65–84
Bensky MK, Gosling SD, Sinn DL (2013) The world from a dog’s point of view. In: Brockmann HJ, Roper TJ, Naguib M, Mitani JC, Simmons LW, Barrett L (eds) A review and synthesis of dog cognition research. Advances in the study of behavior, vol 45. Elsevier Academic Press, San Diego, pp 209–406 (xi, 492)
Boitani L, Ciucci P (1995) Comparative social ecology of feral dogs and wolves. Ethol Ecol Evol 7:49–72
Bowirrat A et al (2012) Neuropsychopharmacology and neurogenetic aspects of executive functioning: should reward gene polymorphisms constitute a diagnostic tool to identify individuals at risk for impaired judgment? Mol Neurobiol 45:298–313
Brady K, Hewison L, Wright H, Zulch H, Cracknell N, Mills D (2018) A spatial discounting test to assess impulsivity in dogs. Appl Anim Behav Sci 202:77–84
Bray EE, MacLean EL, Hare BA (2014) Context specificity of inhibitory control in dogs. Anim Cogn 17:15–31
Bray EE, MacLean EL, Hare BA (2015) Increasing arousal enhances inhibitory control in calm but not excitable dogs. Anim Cogn 18:1317–1329
Bray EE, Sammel MD, Cheney DL, Serpell JA, Seyfarth RM (2017a) Effects of maternal investment, temperament, and cognition on guide dog success. PNAS Proc Natl Acad Sci U S A 114:9128–9133. https://doi.org/10.1073/pnas.1704303114
Bray EE, Sammel MD, Seyfarth RM, Serpell JA, Cheney DL (2017b) Temperament and problem solving in a population of adolescent guide dogs. Anim Cogn 20:923–939. https://doi.org/10.1007/s10071-10017-11112-10078
Brucks D, Marshall-Pescini S, Wallis LJ, Huber L, Range F (2017) Measures of dogs’ inhibitory control abilities do not correlate across tasks. Front Psychol 8:849
Bunford N, Csibra B, Petak C, Ferdinandy B, Miklosi A, Gacsi M (2019) Associations among behavioral inhibition and owner-rated attention, hyperactivity/impulsivity, and personality in the domestic dog (Canis familiaris). J Comp Psychol 133:233–243
Carlson SM, Wang TS (2007) Inhibitory control and emotion regulation in preschool children. Cogn Dev 22:489–510
Casey RA, Loftus B, Bolster C, Richards GJ, Blackwell EJ (2014) Human directed aggression in domestic dogs (Canis familiaris): occurrence in different contexts and risk factors. Appl Anim Behav Sci 152:52–63
Cepeda NJ, Kramer AF, Gonzalez de Sather J (2001) Changes in executive control across the life span: examination of task-switching performance. Dev Psychol 37:715
Chapagain D, Viranyi Z, Wallis LJ, Huber L, Serra J, Range F (2017) Aging of attentiveness in border collies and other pet dog breeds: the protective benefits of lifelong training. Front Aging Neurosci 9:100
Chapagain D, Range F, Huber L, Viranyi Z (2018a) Cognitive aging in dogs. Gerontology 64:165–171
Chapagain D, Viranyi Z, Huber L, Serra J, Schoesswender J, Range F (2018b) Effect of age and dietary intervention on discrimination learning in pet dogs. Front Psychol 9:2217
Clark H, Elsherif MM, Leavens DA (2019) Ontogeny vs Phylogeny in primate/canid comparisons: a meta-analysis of the object choice task. Neurosci Biobehav Rev 105:178–189
Cleal M, Fontana BD, Ranson DC, McBride SD, Swinny JD, Redhead ES, Parker MO (2020) The Free-movement pattern Y-maze: a cross-species measure of working memory and executive function. Behav Res Methods 3:1–22
Cobb M, Branson N, McGreevy P, Lill A, Bennett P (2015) The advent of canine performance science: offering a sustainable future for working dogs. Behav Process 110:96–104. https://doi.org/10.1016/j.beproc.2014.1010.1012
Cohen EEB (2018) A longitudinal study of maternal style, young adult temperament and cognition, and program outcome in guide dogs. University of Pennsylvania, Pennsylvania
Coppola CL, Enns R, Grandin T (2006) Noise in the animal shelter environment: building design and the effects of daily noise exposure. J Appl Anim Welf Sci 9:1–7
Cotman CW, Head E (2008) The canine(dog) model of human aging and disease: dietary, environmental and immunotherapy approaches. J Alzheimer’s Dis 15:685–707
Cotman CW, Head E, Muggenburg BA, Zicker S, Milgram NW (2002) Brain aging in the canine: a diet enriched in antioxidants reduces cognitive dysfunction. Neurobiol Aging 23:809–818
Cowan N (2017) The many faces of working memory and short-term storage. Psychon Bull Rev 24:1158–1170. https://doi.org/10.3758/s13423-13016-11191-13426
Davis PR et al (2017) Abeta vaccination in combination with behavioral enrichment in aged beagles: effects on cognition Abeta, and microhemorrhages. Neurobiol Aging 49:86–99
de Rivera C, Boutet I, Zicker SC, Milgram NW (2005) A novel method for assessing contrast sensitivity in the beagle dog is sensitive to age and an antioxidant enriched food. Prog Neuropsychopharmacol Biol Psychiatry 29:379–387
Diamond A (1985) Development of the ability to use recall to guide action, as indicated by infants' performance on AB. Child Dev 1:868–883
Diamond A (2013) Executive functions. Ann Rev Psychol 64:135–168.
Diamond A, Ling DS (2016) Conclusions about interventions, programs, and approaches for improving executive functions that appear justified and those that, despite much hype, do not. Dev Cogn Neurosci 18:34–48
Eakin L et al (2004) The marital and family functioning of adults with ADHD and their spouses. J Attent Disord 8:1–10
Eisenberg DP, Berman KF (2010) Executive function, neural circuitry, and genetic mechanisms in schizophrenia. Neuropsychopharmacology 35:258–277
Eisenberg N, Spinrad TL (2004) Emotion-related regulation: sharpening the definition. Child Dev 75:334–339
Espy KA, Kaufmann PM, McDiarmid MD, Glisky ML (1999) Executive functioning in preschool children: performance on A-not-B and other delayed response format tasks. Brain Cogn 41:178–199
Fagnani J, Barrera G, Carballo F, Bentosela M (2016) Is previous experience important for inhibitory control? A comparison between shelter and pet dogs in A-not-B and cylinder tasks. Anim Cogn 19:1165–1172. https://doi.org/10.1007/s10071-10016-11024-z
Fahnestock M, Marchese M, Head E, Pop V, Michalski B, Milgram WN, Cotman CW (2012) BDNF increases with behavioral enrichment and an antioxidant diet in the aged dog. Neurobiol Aging 33:546–554
Fay-Stammbach T, Hawes DJ, Meredith P (2014) Parenting influences on executive function in early childhood: a review. Child Dev Perspect 8:258–264
Fernandes JG, Olsson I, Vieira de Castro AC (2017) Do aversive-based training methods actually compromise dog welfare?: A literature review. Appl Anim Behav Sci 196:1–12
Finn PR, Hall J (2004) Cognitive ability and risk for alcoholism: short-term memory capacity and intelligence moderate personality risk for alcohol problems. J Abnorm Psychol 113:569
Fone KC, Porkess MV (2008) Behavioural and neurochemical effects of post-weaning social isolation in rodents—relevance to developmental neuropsychiatric disorders. Neurosci Biobehav Rev 32:1087–1102
Fuller JL (1966) Transitory effects of experiential deprivation upon reversal learning in dogs. Psychon Sci 4:273–274
Garon N, Bryson SE, Smith IM (2008) Executive function in preschoolers: a review using an integrative framework. Psychol Bull 134:31–60. https://doi.org/10.1037/0033-2909.1134.1031.1031
Gathercole SE, Alloway TP, Kirkwood HJ, Elliott JG, Holmes J, Hilton KA (2008) Attentional and executive function behaviours in children with poor working memory. Learn Ind Differ 18:214–223
Gioia GA, Isquith PK, Guy SC, Kenworthy L (2000) Test review behavior rating inventory of executive function. Child Neuropsychol 6:235–238
Granvald V, Marciszko C (2016) Relations between key executive functions and aggression in childhood. Child Neuropsychol 22:537–555
Gunter L (2018) Understanding the impacts of breed identity, post-adoption and fostering interventions, and behavioral welfare of shelter dogs. Arizona State University, Arizona
Harvey ND, Craigon PJ, Blythe SA, England GC, Asher L (2016) Social rearing environment influences dog behavioral development. J Vet Behav Clin Appl Res 16:13–21
Haverbeke A, Laporte B, Depiereux E, Giffroy J, Diederich C (2008) Training methods of military dog handlers and their effects on the team’s performances. Appl Anim Behav Sci 113:110–122
Head E et al (2012) A combination cocktail improves spatial attention in a canine model of human aging and Alzheimer’s disease. J Alzheimer’s Dis 32:1029–1042
Herron ME, Shofer FS, Reisner IR (2009) Survey of the use and outcome of confrontational and non-confrontational training methods in client-owned dogs showing undesired behaviors. Appl Anim Behav Sci 117:47–54
Hiby E, Rooney N, Bradshaw J (2004) Dog training methods: their use, effectiveness and interaction with behaviour and welfare. Anim Welf Potters Bar Wheathampstead 13:63–70
Hillman CH, McAuley E, Erickson KI, Liu-Ambrose T, Kramer AF (2019) On mindful and mindless physical activity and executive function: a response to Diamond and Ling (2016). Dev Cogn Neurosci 37:100529
Homberg JR, Pattij T, Janssen MC, Ronken E, De Boer SF, Schoffelmeer AN, Cuppen E (2007) Serotonin transporter deficiency in rats improves inhibitory control but not behavioural flexibility. Eur J Neurosci 26:2066–2073
Horschler DJ, Hare B, Call J, Kaminski J, Miklósi Á, MacLean EL (2019) Absolute brain size predicts dog breed differences in executive function. Anim Cogn 22:187–198
Houben K, Wiers RW, Jansen A (2011) Getting a grip on drinking behavior: training working memory to reduce alcohol abuse. Psychol Sci 22:968–975
Hsu Y, Serpell JA (2003) Development and validation of a questionnaire for measuring behavior and temperament traits in pet dogs. J Am Vet Med Assoc 223:1293–1300
Hughes C, Devine RT (2019) For better or for worse? Positive and negative parental influences on young children’s executive function. Child Dev 90:593–609
Hughes J, Macdonald DW (2013) A review of the interactions between free-roaming domestic dogs and wildlife. Biol Cons 157:341–351
Karr JE, Areshenkoff CN, Rast P, Hofer SM, Iverson GL, Garcia-Barrera MA (2018) The unity and diversity of executive functions: a systematic review and re-analysis of latent variable studies. Psychol Bull 144:1147
Kelly DM, Adolphe JL, Vernouillet A, McCausland J, Rankovic A, Verbrugghe A (2019) Motoric self-regulation by sled dogs and pet dogs and the acute effect of carbohydrate source in sled dogs. Anim Cogn 22(6):931–946. https://doi.org/10.1007/s10071-10019-01285-y
Kimonis ER, Frick PJ, Boris NW, Smyke AT, Cornell AH, Farrell JM, Zeanah CH (2006) Callous-unemotional features, behavioral inhibition, and parenting: Independent predictors of aggression in a high-risk preschool sample. J Child Fam Stud 15:741–752
Kis A, Gácsi M, Range F, Virányi Z (2012) Object permanence in adult common marmosets (Callithrix jacchus): not everything is an “A-not-B” error that seems to be one. Anim Cogn 15:97–105. https://doi.org/10.1007/s10071-10011-10436-z
Langbein J (2018) Motor self-regulation in goats (Capra aegagrus hircus) in a detour-reaching task. PeerJ 6:e5139
Lawson RA et al (2015) Everyday executive function impairments predict comorbid psychopathology in autism spectrum and attention deficit hyperactivity disorders. Neuropsychology 29:445
Lazarowski L, Dorman DC (2015) A comparison of pet and purpose-bred research dog (Canis familiaris) performance on human-guided object-choice tasks. Behav Process 110:60–67.
Lazarowski L, Thompkins A, Krichbaum S, Waggoner LP, Deshpande G, Katz JS (2020) Comparing pet and detection dogs (Canis familiaris) on two aspects of social cognition. Learn Behav 48(4):1–12
Leen-Feldner EW, Zvolensky MJ, Feldner MT, Lejuez C (2004) Behavioral inhibition: relation to negative emotion regulation and reactivity. Personal Individ Differ 36:1235–1247
Lefebvre D, Diederich C, Delcourt M, Giffroy J-M (2007) The quality of the relation between handler and military dogs influences efficiency and welfare of dogs. Appl Anim Behav Sci 104:49–60
Li CsR, Sinha R (2008) Inhibitory control and emotional stress regulation: neuroimaging evidence for frontal–limbic dysfunction in psycho-stimulant addiction. Neurosci Biobehav Rev 32:581–597
Logue SF, Gould TJ (2014) The neural and genetic basis of executive function: attention, cognitive flexibility, and response inhibition. Pharmacol Biochem Behav 123:45–54
Lopez BR, Lincoln AJ, Ozonoff S, Lai Z (2005) Examining the relationship between executive functions and restricted, repetitive symptoms of autistic disorder. J Autism Dev Disord 35:445–460
Lui M, Tannock R (2007) Working memory and inattentive behaviour in a community sample of children. Behav Brain Funct 3:12
MacLean EL et al (2014) The evolution of self-control. Proc Natl Acad Sci 111:E2140–E2148. https://doi.org/10.1073/pnas.1323533111
MacLean EL, Bray EE (2019) Ontogeny and heritability of cognitive and behavioral traits linked to success as a military working dog. Dunke Univeristy, Durham
Marcovitch S, Zelazo PD (1999) The A-not-B error: results from a logistic meta-analysis. Child Dev 70:1297–1313
Marshall-Pescini S, Frazzi C, Valsecchi P (2016) The effect of training and breed group on problem-solving behaviours in dogs. Anim Cogn 19:571–579
Martel M, Nikolas M, Nigg JT (2007) Executive function in adolescents with ADHD. J Am Acad Child Adolesc Psychiatry 46:1437–1444
McMillan FD (2017) Behavioral and psychological outcomes for dogs sold as puppies through pet stores and/or born in commercial breeding establishments: current knowledge and putative causes. J Vet Behav Clin Appl Res 19:14–26
McMillan FD, Serpell JA, Duffy DL, Masaoud E, Dohoo IR (2013) Differences in behavioral characteristics between dogs obtained as puppies from pet stores and those obtained from noncommercial breeders. J Am Vet Med Assoc 242:1359–1363
Meijers J, Harte JM, Jonker FA, Meynen G (2015) Prison brain? Executive dysfunction in prisoners. Front Psychol 6:43
Meiran N, Diamond GM, Toder D, Nemets B (2011) Cognitive rigidity in unipolar depression and obsessive compulsive disorder: Examination of task switching Stroop, working memory updating and post-conflict adaptation. Psychiatry Res 185:149–156
Miklósi Á (2014) Dog behaviour, evolution, and cognition. OUP, Oxford
Miyake A, Friedman NP, Emerson MJ, Witzki AH, Howerter A, Wager TD (2000) The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: a latent variable analysis. Cogn Psychol 41:49–100. https://doi.org/10.1006/cogp.1999.0734
Moffitt TE et al (2011) A gradient of childhood self-control predicts health, wealth, and public safety. Proc Natl Acad Sci 108:2693–2698
Moffitt TE (2012) Childhood self-control predicts adult health, wealth, and crime. In: Multi-Discipl. Symp. Improv. Well-Being Children Youth, 2012
Moher D, Liberati A, Tetzlaff J, Altman DG, P. Grp (2009) Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement (Reprinted from Annals of Internal Medicine). Phys Ther 89:873–880
Mongillo P, Pitteri E, Candaten M, Marinelli L (2016) Can attention be taught? Interspecific attention by dogs (Canis familiaris) performing obedience tasks. Appl Anim Behav Sci 182:30–37
Mongillo P, Scandurra A, Eatherington CJ, D’Aniello B, Marinelli L (2019) Development of a spatial discount task to measure impulsive choices in dogs. Animals 9:469
Nigg JT (2001) Is ADHD a disinhibitory disorder? Psychol Bull 127:571
Oliva J, Rault J, Appleton B, Lill A (2015) Oxytocin enhances the appropriate use of human social cues by the domestic dog (Canis familiaris) in an object choice task. Anim Cogn 18:767–775
Olsen MR (2018) A case for methodological overhaul and increased study of executive function in the domestic dog (Canis lupus familiaris). Ani Cogn 21:175–195. https://doi.org/10.1007/s10071-10018-11162-10076
Overall KL, Dunham AE, Scheifele P, Sonstrom Malowski K (2019) Fear of noises affects canine problem solving behavior and locomotion in standardized cognitive tests. Appl Anim Behav Sci 221:104863
Pecora G, Zoratto F, Paoletti M, Bellagamba F, Paglieri F, Addessi E (2017) Executive Function. In: Vonk J, Shackelford T (eds) Encyclopedia of animal cognition and behavior. Springer International Publishing, Cham, pp 1–5. doi:https://doi.org/10.1007/978-3-319-47829-6_1048-1
Piaget J (1954) The construction of reality in the child (M. Cook, Trans.). New York, NY, USA
Pirrone F, Pierantoni L, Pastorino GQ, Albertini M (2016) Owner-reported aggressive behavior towards familiar people may be a more prominent occurrence in pet shop-traded dogs. J Vet Behav 11:13–17
Poland SE, Monks CP, Tsermentseli S (2016) Cool and hot executive function as predictors of aggression in early childhood: Differentiating between the function and form of aggression. Brit J Dev Psychol 34:181–197
Polgar Z, Blackwell EJ, Rooney NJ (2019) Assessing the welfare of kennelled dogs—a review of animal-based measures. Appl Anim Behav Sci 213:1–13
Puig M, Rose J, Schmidt R, Freund N (2014) Dopamine modulation of learning and memory in the prefrontal cortex: insights from studies in primates, rodents, and birds. Front Neural Circ 8:93
Raaijmakers MA, Smidts DP, Sergeant JA, Maassen GH, Posthumus JA, Van Engeland H, Matthys W (2008) Executive functions in preschool children with aggressive behavior: impairments in inhibitory control. J Abnorm Child Psychol 36:1097
Reese ED, Zielinski MJ, Veilleux JC (2015) Facets of mindfulness mediate behavioral inhibition systems and emotion dysregulation. Personal Individ Differ 72:41–46
Rooney N, Gaines S, Hiby E (2009) A practitioner’s guide to working dog welfare. J Vet Behav 4:127–134
Sallinen J, Haapalinna A, Viitamaa T, Kobilka BK, Scheinin M (1998) Adrenergic α2C-receptors modulate the acoustic startle reflex, prepulse inhibition, and aggression in mice. J Neurosci 18:3035–3042
Schipper LL, Vinke CM, Schilder MB, Spruijt BM (2008) The effect of feeding enrichment toys on the behaviour of kennelled dogs (Canis familiaris). Appl Anim Behav Sci 114:182–195
Sherman EM, Brooks BL (2010) Behavior rating inventory of executive function–preschool version (BRIEF-P): test review and clinical guidelines for use. Child Neuropsychol 16:503–519
Sonntag Q, Overall K (2014) Key determinants of dog and cat welfare: behaviour, breeding and household lifestyle. Rev Sci Tech Off Int Epiz 33:213–220
Szabo B, Noble DW, Whiting MJ (2019) Context-specific response inhibition and differential impact of a learning bias in a lizard. Anim Cogn. 22(3):317–329. https://doi.org/10.1007/s10071-10019-01245-10076
Thompson WR, Heron W (1954) The effects of restricting early experience on the problem-solving capacity of dogs. Can J Psychol/Rev Can Psychol 8:17–31
Toplak ME, Bucciarelli SM, Jain U, Tannock R (2008) Executive functions: performance-based measures and the behavior rating inventory of executive function (BRIEF) in adolescents with attention deficit/hyperactivity disorder (ADHD). Child Neuropsychol 15:53–72
Troisi CA, Mills DS, Wilkinson A, Zulch HE (2019) Behavioral and cognitive factors that affect the success of scent detection dogs. Comp Cogn Behav Rev 14:51–76
Tucker AM, Whitney P, Belenky G, Hinson JM, Van Dongen HP (2010) Effects of sleep deprivation on dissociated components of executive functioning. Sleep 33:47–57
Udell MA, Dorey NR, Wynne CD (2008) Wolves outperform dogs in following human social cues. Anim Behav 76:1767–1773
Udell MA, Dorey NR, Wynne CD (2010) The performance of stray dogs (Canis familiaris) living in a shelter on human-guided object-choice tasks. Anim Behav 79:717–725
Vas J, Topal J, Pech E, Miklosi A (2007) Measuring attention deficit and activity in dogs: a new application and validation of a human ADHD questionnaire. Appl Anim Behav Sci 103:105–117
Vaterlaws-Whiteside H, Hartmann A (2017) Improving puppy behavior using a new standardized socialization program. Appl Anim Behav Sci 197:55–61
Vernouillet AA, Stiles LR, McCausland JA, Kelly DM (2018) Individual performance across motoric self-regulation tasks are not correlated for pet dogs. Learn Behav 46:522–536
von Bastian CC, Oberauer K (2013) Distinct transfer effects of training different facets of working memory capacity. J Mem Lang 69:36–58
Wallis LJ, Range F, Muller CA, Serisier S, Huber L, Viranyi Z (2014) Lifespan development of attentiveness in domestic dogs: drawing parallels with humans. Front Psychol 5:71
Wells DL (2004) A review of environmental enrichment for kennelled dogs Canis familiaris. Appl Anim Behav Sci 85:307–317
Welsh M, Gorman Barry P, Jacobs AA, Beddes LA (2018) Homeless men living in transitional housing: the brainwise program and improvements in executive functions and coping self-efficacy. SAGE Open 8:2158244018769138
Williams LE, Huang JY, Bargh JA (2009) The scaffolded mind: Higher mental processes are grounded in early experience of the physical world. Eur J Soc Psychol 39:1257–1267
Willie A (2011) Does emotion dysregulation mediate the relationship between behavioural inhibition and psychopathological symptoms? University of Southampton, Southampton
Wright HF, Mills DS, Pollux PM (2012) Behavioural and physiological correlates of impulsivity in the domestic dog (Canis familiaris). Physiol Behav 105:676–682
Zaine I, Domeniconi C, Wynne CD (2015) The ontogeny of human point following in dogs: when younger dogs outperform older. Behav Proc 119:76–85
Ziv G (2017) The effects of using aversive training methods in dogs—a review. J Vet Behav 19:50–60
Funding
This research is supported by a Research Training Program (RTP) Stipend Scholarship and a Full Fee Research Scholarship (LTUFFRS) awarded to Maike Foraita by La Trobe University.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that no conflict of interest or competing interests exist.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Foraita, M., Howell, T. & Bennett, P. Environmental influences on development of executive functions in dogs. Anim Cogn 24, 655–675 (2021). https://doi.org/10.1007/s10071-021-01489-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-021-01489-1