
Abstract
Teaching is a powerful form of social learning, but there is little systematic evidence that it occurs in species other than humans. Using long-term video archives the foraging behaviors by mother Atlantic spotted dolphins (Stenella frontalis) were observed when their calves were present and when their calves were not present, including in the presence of non-calf conspecifics. The nine mothers we observed chased prey significantly longer and made significantly more referential body-orienting movements in the direction of the prey during foraging events when their calves were present than when their calves were not present, regardless of whether they were foraging alone or with another non-calf dolphin. Although further research into the potential consequences for the naïve calves is still warranted, these data based on the maternal foraging behavior are suggestive of teaching as a social-learning mechanism in nonhuman animals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Although one of the most potent forms of social learning in humans, there has been little evidence to suggest that teaching occurs in nonhuman animals. Some theorists have suggested that this may be because teaching requires advanced social-cognitive skills, including the ability to take the perspective of another and theory of mind, the ability to appreciate that an individual’s behavior is based on its knowledge and its desires (Boesch and Tomasello 1998; Tomasello 1996, 2000; Tomasello et al. 1993). For example, previous examples of suggested teaching behavior by meerkats, cheetahs, and domestic cats seem to benefit the prey-handling abilities of the young, but do not require the use of higher cognitive mechanisms (Thornton and McAuliffe 2006; Caro and Hauser 1992). However, Caro and Hauser (1992) provided a definition of teaching that may be more inclusive for nonhuman animals, defining it as, “An individual actor A can be said to teach if it modifies its behavior only in the presence of a naive observer, B, at some cost or at least without obtaining an immediate benefit for itself. A’s behavior thereby encourages or punishes B’s behavior, or provides B with experience, or sets an example for B. As a result, B acquires knowledge or learns a skill earlier in life or more rapidly or efficiently than it might otherwise do, or that it would not learn at all” (p. 153).
Previous studies that suggested teaching in primates and cetaceans, although promising, lacked systematic measurement of the behavior. Probably the best evidence of teaching in nonhuman primates to date is Boesch’s studies of mother chimpanzees (Pan troglodytes) in the Tai National Park of the Ivory Coast (Boesch 1991, 1993; Greenfield et al. 2000). Boesch suggested that the chimpanzee mothers facilitated the development of their offspring’s nut-cracking skills by means of stimulation, facilitation, and active teaching. Nut cracking is observed in only a few populations of chimpanzees, despite the availability of nuts and appropriate tools (i.e., rocks appropriate for use as anvils and hammers), qualifying, by some definitions, as an example of culturally-transmitted behavior (Whiten 2005; Whiten et al. 1999). The interpretation of such episodes as “teaching” has been questioned, however (Bering 2001; Bering and Povinelli 2003). Moreover, such episodes are rarely observed, suggesting that direct teaching is not a common form of cultural transmission in chimpanzees.
Although evidence of social learning is easier to document in these terrestrial great apes than it is in marine mammals, nongenetic transmission of behavior across generations has also been observed for cetaceans (Kruetzen et al. 2005; Rendell and Whitehead 2001), suggesting that, like the great apes, these large-brained, slow-developing, and socially complex species (Bjorklund and Bering 2003) have evolved powerful social-learning mechanisms.
Similar to research with great apes, little is known about the actual mechanism of transmission across generations in cetaceans. Herzing (2005) described some potential scenarios and mechanisms observed for a group of free-ranging Atlantic spotted dolphins (Stenella frontalis), including implications of vertical, horizontal, and oblique directions of transmission of information during various behavioral contexts. Recently, Spininelli et al. (2006) described prey-transfer between mother and calf in the marine tucuxi dolphin (Sotalia fluvialis). Observations such as these suggest that the mother-calf relationship may be one of the most important sources of information in the young calf’s life.
There is some evidence of presumed maternal teaching behavior associated with stranding behavior as a foraging specialization used by part of the population of killer whales (Orcinus orca) in the Crozet Islands and off Punta Norte, Argentina to capture seal pups on pinniped breeding beaches. Adult females demonstrated a modification of their strand foraging behavior in the presence of naïve juvenile observers (presumably their calves), suggesting that teaching may be involved in the development and the rate of success of calves in mastering these behaviors (Guinet and Bouvier 1995). This comparison between purported teaching in chimpanzees and killer whales is interesting because any commonalities would have been derived through convergent evolution, as the last common ancestor of primates with cetaceans is estimated to have lived over 90 million years ago (Marino et al. 2007). However, despite the multitude of observations of chimpanzees and killer whales in the wild, incidences of mother–infant teaching are scarce and anecdotal in nature.
Foraging behavior is a likely candidate for social learning among wild dolphins, particularly between mother and calf. The mother/calf relationship is the strongest association that Atlantic spotted dolphins have in their lifetimes (Herzing and Brunnick 1997). The prolonged developmental period provides both ample time and situational possibilities for a calf to learn foraging strategies from its mother (Herzing 1996). The majority of daytime feeding behavior of Atlantic spotted dolphins is benthic foraging, in which dolphins use echolocation to locate fish in the sandy bottom and then dig prey items out of the sand in order to catch and eat them (Herzing 1996).
Much of what we know about marine mammal social learning comes from research with captive animals due, in part, to the difficulty of studying such phenomena in the wild. Attempts to assess teaching among captive animals often involve contrived situations, which may affect the animals’ success or failure (Kuczaj et al. 2005). Studies that assess social learning in wild populations of marine mammals are needed to validate and supplement the findings from captive animals and to better understand the spontaneous occurrence of social learning in natural settings.
In the present study, using video archives from a long-term naturalistic study, we investigated social learning and possible teaching behavior by the Atlantic spotted dolphin. Unlike most previous research examining social learning in nonhuman mammals in which the focus is on the observer/learner (e.g., Bjorklund et al. 2002; Guinet and Bouvier 1995; Herman 2002; Tomasello et al. 1993), the present study shifted the focus from the observer (in our case, the calf observing the foraging behavior) to the model (the mother performing the foraging behavior) to explore the possibility of teaching behavior. We examined the foraging behavior by mother dolphins when foraging in the presence of their young (less than 3-year-old) calves in comparison to when the calves were not present. Additionally, comparisons were then made between mothers foraging alone and when they were foraging with non-calf conspecifics. Should the mothers alter their foraging behavior in the presence of their young, it would be suggestive of teaching and provide a possible mechanism for cultural transmission within this dolphin species.
Methods and analysis
Natural history
This study was performed using underwater video recordings of the Atlantic spotted dolphin collected by the Wild Dolphin Project (Herzing 1997) in the study area north of Grand Bahama Island, Bahamas during summer field seasons between 1991 and 2004 (Fig. 1). Unlike many other marine mammal habitats, this area is optimal for behavioral observation with clear, warm waters that allow for excellent visibility up to 90 ft and long observational periods (Herzing 1996). The dolphins in this study have been observed since 1985, and include over 200 dolphins that have been individually recognized and sexed. Atlantic spotted dolphins can be categorized into four age classes, based on the pigmentation of the individual (Herzing 1996). The number of spots on the individual is correlated with age, with a newborn having no spots. Although most individuals in the population are tracked from birth, this allows for approximating the individual’s age when it is not known from previous sightings. Calves in the sampled foraging events 3-years-old or younger were still observed to be nursing during the encounter year. The year of the calf’s birth was determined from previous sightings of the pregnant female followed by a sighting with a closely associated calf or a sighting of the mother with a suckling calf (Herzing 1997).

Map of study area for the wild dolphin project underwater video recordings collected over sandbanks north of Grand Bahama Island Study population of Atlantic spotted dolphins ranges over an area of approximately 500 km2
Apparatus and procedure
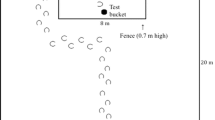
Video was recorded using various underwater cameras (Sony CCDV9 8-mm, Yashica KXV Hi8-mm with attached Labcore 76 hydrophone, Sony DCR-SC100 NTSC, or Sony DCR-PC110 NTSC Digital Video). Underwater video sequences were analyzed using focal follow (of the mother with and without the calf) as a sampling rule and continuous sampling as a recording rule. Video sequences from the long-term video archives from the Wild Dolphin Project® between 1991 and 2004 were assessed for the presence of benthic foraging events by individual mothers, either with or without their calves present, based on a behavioral ethogram designed to measure the individual benthic foraging behavior of the mother dolphins (see Fig. 2). Each benthic fish catch was broken down into a series of behaviors that made up a typical foraging event.

Visual ethogram of select foraging behavioral events (Miles and Herzing 2003). The visual ethogram includes the possible positions in which calves were observed during foraging events in relation to their mother and the sequence of behaviors for a benthic foraging event
For the purpose of this study, a foraging event was defined as the series of behaviors performed by the dolphin in order to catch the prey animal. For benthic feeding, the series of events was as follows:
Scanning is observed when the dolphin moves its head horizontally or vertically repeatedly while performing a directional swim, and usually occurs near the sea floor and can be followed by a dig or fish catch (Miles and Herzing 2003). Rooting or digging is observed when the dolphin inserts the rostrum into the sea floor or sandy bottom to dig the prey out of the substrate in the attempt to capture the prey. Chasing is observed as swimming in the direction of the prey object, as in pursuit of the prey. For our purposes, ingestion was defined as the food going into mouth and never being seen again. The categories of foraging behaviors measured were chase latency and number of body-orienting movements during pursuit. Chase latency, the length of time the prey was pursued, was operationally defined as the period of time when the fish appeared out of the sand (was rooted out) until the time of ingestion by the dolphin, or the dolphin no longer pursued prey (lost interest in prey). Body-orienting movements were measured to examine the dolphin’s attention to the prey object. A body-orienting movement was measured in foraging events as a movement of the body reorienting in direction of prey object, often seen during pursuit of prey, from the time the prey was rooted out of the sand. Body-orienting movements were particularly interesting as they appeared to be exaggerated movements in the direction of the prey, which may be an attention-directed referential behavior similar to the spontaneous pointing observed by dolphins during experiments in captivity (Xitco et al. 2001). Figure 2 is a visual ethogram describing the sequence of the benthic foraging behavior (Miles and Herzing 2003).
The segments from the video archives in which mothers were observed engaged in foraging behavior with or without their calves were then viewed and then examined for further criteria. Thirty-eight video segments were used in the study based on the selection criteria: fourteen video segments of mothers foraging with their calves present and 24 video segments of those mothers foraging without their calves present. Videos were selected based on the presence of a target female performing an individual foraging event, as well as on the following designated video acceptance criteria: (a) the individuals were identifiable; (b) it was possible to identify the beginning and end of the chase sequence; (c) the prey was visible, or if the prey was not completely visible, it was possible to identify the position of the prey based on the behavior of the dolphin; (d) if the calf was present, the calf was in a nearby position (within a proximity of two body lengths) from which it was capable of observing its mother; and (e) if the calf was present, it was possible to identify the position of the calf relative to the mother during the foraging event. The position of the calf relative to the mother was recorded as Infant, Head-under-head, Echelon, Observation, or Other, with comments where necessary, as depicted in Fig. 2.
The mothers’ foraging behaviors were then individually measured for the variables of chase latency and body-orienting movements, both when foraging alone (or with other juvenile or adult dolphins) and when foraging in the presence of their calves. Other variables, such as types of play involved in that foraging encounter, position of calf relative to the mother, directionality of the calf, age of the mothers and their calves, whether the mother eventually ate the prey; the prey species, when identifiable, were also recorded for each foraging event.
Participants
The foraging behaviors of nine mother dolphins were recorded both with (n = 14) and without (n = 22) their calves present. Ten different calves were observed with the nine mothers in the 14 foraging events, with one mother observed during separate events with two different calves. Calves ranged in age from neonate to 3 years old. All calves were observed to be nursing within the same field season as the foraging event. Ages of the mothers were known, or were estimated based on their age class. It is important to note that some foraging events without calves present (10 of the 22) occurred while the target female was still a juvenile (prior to sexual maturity); however, during those events, the “mothers to be” were already past the age of weaning and were independently foraging. The minimum age of any female during foraging events over the 12-year period was 10 years for mothers foraging with their calves and 4 years for mothers-to-be foraging without calves present. Of the observations without calves present, four of the females were observed foraging as juveniles, prior to becoming mothers. Of the nine mothers observed, foraging events were observed for one female both when she was a calf with her mother, and later as a mother herself with her own calf.
The video segments of the foraging events were shortened to within 1 min of the beginning and end of the foraging event and labeled with the foraging event number and the individuals involved. Segments were then watched by the first author and two independent observers to measure the desired behaviors. Of the 38 total foraging events measured, 32 were measured by the first author and two independent observers; the other six were measured by the first author and only one independent observer. For the measurement of chase latencies of foraging events, there was significant correlation between the author and the first independent observer, r 36 = 1.0, P < 0.001, between the author and the second independent observer, r 30 = 0.999, P < 0.001, and between the first and second observer, r 30 = 0.998, P < 0.001. For the measurement of number of body-orienting movements, there was significant correlation between the author and the first independent observer, r 36 = 1.0, P < 0.001, between the author and the second independent observer, r 30 = 0.988, P < 0.001, and between the first and second observer, r 30 = 0.988, P < 0.001.
The videos were watched and timed using Windows Media Player version 10 on a Hewlett Packard laptop computer and a projector in order to enlarge the viewing area. Due to the restrictions of the media software used for editing and playing the video, a “second or less” rule was instituted for measurement of latencies in which the latencies appearing to be less than 1 s were rounded up to 1 s in duration.
Results
Chase latencies
Mean chase latencies for each of the nine mothers, both when foraging with and without their calves, is presented in Table 1. The mothers chased the prey significantly longer when their calves were present (M = 22.24 s, SD = 9.36) than when their calves were not present (M = 2.74 s, SD = 1.47), t 8 = 6.57, P < 0.001, d = 1.14. Mean chase latencies were longer when foraging with their calves than without their calves for each of the nine mothers (Fig. 3).

Mean chase latencies of mothers foraging with and without their calves present
Body-orienting movements
Mean number of body-orienting movements for each of the nine mothers, both when foraging with and without their calves, is presented in Table 1. The mothers made significantly more body-orienting movements when their calves were present (M = 1.26, SD = 1.04) than when their calves were not present (M = 0.28, SD = 0.31), t 8 = 2.46, P = 0.04, d = 2.31. Six of nine mothers made more body-orienting movements when foraging in the presence of their calves than when not foraging with their calves.
Cross-generational comparisons
Two consecutive generations of the mother/calf pairs in the study showed the same altered foraging behavior. In the first generation, one mother, Nippy, and her calf, Nassau, demonstrated the presumed teaching behavior. In the second generation, Nassau, now a mother, showed the same altered foraging behavior as her mother with her calf, Neptune.
Foraging with non-calf individuals
Of the foraging events without calves, three mothers were observed foraging both alone and with other individuals that were not calves, at least juveniles or older. The chase latencies and body-orienting movements for these mothers were compared for the two conditions (alone and with non-calf individuals). There were too few subjects (n = 3) to perform a statistical test; however, the chase latencies of these three mothers foraging alone (M = 1.33, SD = 0.58) were comparable to the chase latencies of these same mothers foraging with non-calf individuals (M = 3.44, SD = 1.50), and both were much lower than the chase latencies of those mothers foraging with their calves present (M = 17.56, SD = 6.26). Additionally, the number of body-orienting movements of these three mothers foraging alone (M = 0.17, SD = 0.29) was comparable to the number of body-orienting movements when they were foraging with non-calf individuals (M = 0.33, SD = 0.58), and both were less than the number of body-orienting movements of those mothers foraging with their calves present (M = 1.94, SD = 0.42).
Additional analyses
The differences observed for chase latencies and number of body-orienting movements could not be attributed to prey type, as there were no significant differences in the foraging behaviors between prey species. Prey species were identified for 14 observations. When foraging with calves present, mothers were observed foraging for Snakefish (family Synodontidae), n = 5, flounder (family Bothidae), n = 2, and razorfish (family Clinidae), n = 1. The mean chase latencies, M Snakefish = 19.0 (SD = 11.03), M Flounder = 20.5 (SD = 14.85), M Razorfish = 25.0 (SD = 0.0), and number of body-orienting movements, M Snakefish = 0.6 (SD = 0.89), M Flounder = 25.0 (SD = 2.12), MRazorfish = 3.0 (SD = 0.0), were comparable for all three species of prey when foraging with calves present. Both snakefish, n = 9, and flounder, n = 4, were observed as prey types when the females were observed foraging without calves present. The mean chase latencies, M Snakefish = 2.22 (SD = 1.09), M Flounder = 3.83 (SD = 3.57), and number of body-orienting movements, M Snakefish = 0.5 (SD = 0.76), M Flounder = 0.5 (SD = 0.58), were comparable for both species when foraging without calves present, and both were lower than when foraging with calves present.
Additionally, individual foraging events of dolphins not included in the study observed foraging for either snakefish or flounder were collected and analyzed. Ten foraging events each for catches of snakefish and flounder using both dolphins in the present study and dolphins not used in the study were randomly selected, and their corresponding chase latencies and number of body-orienting movements were compared between the two types of prey using an independent samples Student’s t-test. There were no significant differences found between the two types of prey for either chase latencies, (M flounder = 4.60, SDflounder = 3.86, M snakefish = 2.80, SDsnakefish = 1.62) t 18 = −1.36, P = 0.19, or number of body orienting movements for dolphins foraging without calves present, (M flounder = 0.50, SDflounder = 0.71, M snakefish = 0.37, SDsnakefish = 0.48) t 18 = −0.49, P = 0.63. This supports the conclusion that the observed differences in this study for mothers foraging with calves present would not likely be due to the type of prey, as chase latencies and the number of body-orienting movements do not normally vary significantly between snakefish and flounder, the two main types of prey observed being caught by mother dolphins in this study.
To assess possible effects of age of calf and age of mother on the dependent measures, correlations with each dependent measure were computed separately with the age of the calf and the age of the mother for the foraging events when the calf was present. Correlations between age of the calf and chase latency, r 13 = 0.37, P = 0.10, and number of body-orienting movements, r 13 = −0.30, P = 0.15, were both nonsignificant. Correlations between mother’s age and number of body-orienting movements were significant, r 13 = −0.47, P = 0.046, with older mothers making fewer body-orienting movements than younger mothers. The correlation between mother’s age and chase latencies was not significant, r 13 = 0.01, P = 0.48. When comparing the foraging behaviors to the age of the mothers when the calves were not present, correlations with both chase latencies r 23 = −0.07, P = 0.73, and number of body-orienting movements, r 23 = −0.12, P = 0.58, were nonsignificant. The foraging behaviors of eight of the nine mothers without calves were compared between those females observed as juveniles and those observed as adults, with three mothers being observed foraging as juveniles and five mothers observed foraging as adults.Footnote 1 There were no significant differences between chase latencies, t (6) = −0.10, P = 0.92, or number of body-orienting movements, t (6) = −0.12, P = 0.91, of females observed foraging when juveniles or adults.
Additionally, of the nine mothers observed in this study, three of the mothers, Little Gash, Mugsy, and PR2, were observed foraging both with and without their calves present during the same year, and each of those three mothers being observed foraging both with and without calves during the same field encounter. The mean chase latencies for these three mothers are compared in Table 2. The chase latencies were much longer foraging with calves than foraging alone for the mothers at the same age and comparable to the mean chase latencies of foraging alone for the mothers at an earlier age.
The position of the calf during the chase period was indicated for each of the foraging events (N = 14) in which the calf was present. Although some foraging events involved multiple calf positions, the observation position was found to be the most common, taking place in 11 (79%) of the foraging events. Head-under-head position was present in one of the foraging events, Echelon was present in two foraging events, and Infant position was seen in one foraging event. Only Infant, Echelon, and Observation positions were seen solely in an individual foraging event, with one, one, and nine occurrences, respectively.
For the foraging events collected in which the ingestion by the mother was known (as seen by video or first-hand account), there were five events, one event each for five of the nine mothers, in which the prey was not eaten. Each of these foraging events occurred when the mothers’ calves were present (36% of the total events with calves). In addition, the calves were allowed to pursue the prey in each of these events and were confirmed to have eaten the prey in three of the foraging events, despite the fact that they were still nursing and not dependent upon fish for food. Four of the nine mothers were observed eating the prey in seven of the fourteen events in which calves were present. In two of the events in which calves were present it was not known whether the prey was eaten.
Discussion
The nine mother dolphins observed in this study displayed significantly longer chase latencies and made significantly more body-orienting movements when foraging in the presence of their calves than when foraging alone. The chase latency data are particularly impressive. Mean chase latencies were eight times longer for the female dolphins when foraging with their calves (22.24 s) than without them (2.74 s). As illustrated in Fig. 3, the distributions were non-overlapping, with every mother having a longer latency when foraging with her calf than when foraging without her calf.
Although differences were not as robust for the body-orienting movements, the overall number of body-orienting movements was significantly greater when the mothers were foraging with their calves than when foraging alone, with six of nine mothers displaying this pattern. Previous research has shown that dolphins are capable of understanding the human gesture of pointing (Herman et al. 1999; Pack and Herman 2004), and of producing spontaneous referential gestures in artificial experimental contexts (Xitco et al. 2001). Although these body-orienting movements were not demonstrated by all sampled mothers, the present results may be evidence of referential gesturing in a more ecologically valid context.
In addition to the elongated chase and presumed referential body-orienting movements in the direction of the prey, the altered foraging behavior in the presence of the calf appeared more exaggerated when compared to the mothers foraging alone or with another non-calf dolphin. Mothers seemed to toy with their prey, making it more like play behavior and less like the typical foraging behavior of mothers observed foraging without their calves present, similar to descriptions of possible teaching of predatory behavior from other species such as cats and killer whales (Caro and Hauser 1992; Guinet and Bouvier 1995). Some of the mothers were observed letting the prey swim away and then digging them back out of the sand, sometimes multiple times, after initially digging the fish out of the sand, and also make jawing motions in the direction of the prey. Mothers also allowed the seemingly attentive calves to participate in the chase, and calves were observed eating the prey in three of the events. Although 4 of the 9 mothers were observed eating the prey in 7 of the 14 events, all of the mothers were observed allowing the calves to participate and made little to no effort in these events to consume the prey, only seemingly facilitating the calves’ experience in chasing the prey. Despite the altered foraging behavior, mothers were never observed losing the prey. In all events in which ingestion of the prey was observed, either the mother or the calf ate the prey, or the mother made no further attempts to ingest the prey and left the prey to the calf to chase. This altered foraging behavior can be observed in the online supplemental movies S1 and S2 which exemplify the markedly different foraging behavior of an adult female foraging alone (S1) with the foraging behavior of a mother foraging in the presence of her calf (S2).
There was a potential cost of the alteration of the mothers’ foraging behavior in their calves’ presence. The elongated chase time allowed more opportunity for prey to escape, although this did not occur in any of the events observed, as well as taking time away from catching additional fish or other activities. Instead of the typical grab and ingestion, these mothers toyed with their prey as their calves watched, and, as mentioned above, sometimes even avoided ingestion in these altered foraging events. Often in teaching, the model will provide an exaggerated or elongated version of the typical behavior in front of the naïve observer, much as the mother chimpanzees observed in Boesch’s (1991) work. These exaggerated foraging behaviors may provide a window of opportunity for the calves to observe, and possibly learn from, the example provided by their mothers, and thus be worthy of the extra time and energy put forth by the mothers at a cost to themselves in order to help ensure the success of their offspring.
An alternative explanation for the altered foraging behavior may be that the mothers were distracted by their calves, resulting in longer chase latencies and exaggerated movements to compensate for their divided attention. However, we do not believe this to be the cause of the altered foraging behavior for a few reasons. First, three of the nine mothers observed in the present study demonstrated the ability of dolphin mothers to forage without distraction even when their calves were in the nearby vicinity, but not directly observing the mothers. These three mothers, Little Gash, Mugsy, and PR2, were observed foraging both with and without their calves present during the same field encounter. These mothers only altered their behavior when the calves were observing the mother, but not during the fish catches in which the calves were within the vicinity and not observing. Table 2 compares the means for those events, in which the mean chase latencies were much longer foraging with calves than foraging alone during the same field encounter and comparable to the mean chase latencies of foraging alone during separate field encounters. These data suggest that the calves’ being merely nearby is not a distraction for the mothers, but instead the altered foraging behavior only occurred when the calves were directly observing the mothers’ behavior. If the calves were a distraction for the mothers, the altered foraging behavior should have occurred whenever the calves were within the vicinity, regardless of whether the calves were directly observing the mothers.
Second, Atlantic spotted dolphins participate in alloparenting, which permits mothers to frequently forage separately from their calves (Herzing 1996). If the mother’s nutritional needs are being met in the absence of their calf with the help of alloparenting, this would enable more time and energy on the part of the mother for altered foraging events for teaching when the calf is present. Additionally, it would not be advantageous for the mother to forage with the calf present solely for her nutritional purposes if there is an alloparent available. In this study, there were potential alloparents, juvenile or adult female dolphins, available in all 14 of the calf-present events that would have permitted the mothers to forage without distraction, which is often observed in Atlantic spotted dolphins (Herzing 1996), including the events for the previously mentioned three mothers observed foraging with and without calves during the same field encounter.
Third, young dolphin calves are very precocious and the calves in the observed foraging events appeared attentive and interactive. Some of the evidence for the calves’ attentiveness to the mothers’ foraging behavior was observed through the calves’ positions relative to the mothers, which was indicated during the chase period for each of the foraging events in which the calf was present. Although previous research (Mann and Smuts 1999) has shown that dolphin calves in the wild, and in captivity spend the majority of their time in infant or echelon position until the time of weaning, calves were most commonly found during the chase period in the observation position relative to their mothers in which the calf was potentially exposed to both visual and acoustic information and in which calves appeared attentive to both the mother and the prey Footnote 2 (Figure 2).
Additionally, the calves were allowed to pursue the prey in five of the foraging events and were confirmed to have eaten the prey in three of the events, despite the fact that they were still nursing and not dependent upon fish for food. It is important to note that the only time when the fish was not ingested was when the mothers were foraging with their calves as part of their altered foraging behavior. The prey was always consumed when foraging alone or in the presence of a non-calf dolphin. This is potentially a costly behavior for the mother as she is depriving herself of calories needed to nourish her still nursing calf by allowing the prey to escape or allowing the calf to eat the prey, as well as the energy to play with the prey rather than just eating it quickly and efficiently herself. It would be more efficient and less costly for the mothers to have simply caught and consumed the fish, or perhaps to have left the calf in the care of an alloparent, as opposed to this altered foraging behavior observed in these events.
Furthermore, the chase latencies and foraging behaviors were drastically different with their calves present, not just with the longer latencies but also with the presumed referential behaviors toward the prey objects and allowing the still nursing calf to participate, which would be more indicative of social learning. If the calves were merely a distraction, the mothers could have either immediately consumed the prey or disciplined the calf, as previously observed by mothers in the population for undesirable behaviors (Miles and Herzing 2003), rather than allowing the calf to participate. Therefore, because the calves appeared attentive, interacted with the prey and the mother, and appeared to need little care during the event, they were not likely a distraction to the mother.
Reciprocally, the mothers may have been altering their foraging behavior because their calves were attentive, so that a calf’s attention may have stimulated the altered maternal foraging behavior. Further analysis into the calves’ behavior is warranted and may also elucidate the exact learning mechanism at play on behalf of the calf.
For the majority of the mothers, there was a difference in maternal age between the events observed foraging without their calves compared to the events foraging with their calves. It is not likely that the age differences, or resulting level of experience would have resulted in the observed differences in chase latencies and number of body-orienting movements. Rather, an older or more experienced dolphin should be expected to have quicker chase latencies and fewer body-orienting movements due to increased efficiency, not the longer latencies and increased number of body-orienting movements as observed here. Additionally, of the nine mothers observed in this study, three of the mothers, Little Gash, Mugsy, and PR2, were observed foraging both with and without their calves present during the same year, and each of these three mothers were observed foraging both with and without calves during the same field encounter. For these three mothers, mean chase latencies were much longer foraging with calves than foraging alone at the same age and comparable to the mean chase latencies of foraging alone at an earlier age (Table 2). This suggests that there was not a difference in foraging behavior due to difference in age or experience, but rather due to the presence of their calves.
There were also no significant differences for chase latencies or number of body-orienting movements for the prey species observed in this study: thus the altered foraging behaviors were also not likely due to the difficulty of catching a specific type of prey.
The three episodes in which a target dolphin was observed foraging with a non-calf individual, juvenile in age or older, produced chase latencies and number of body-orienting movements comparable to when these dolphins foraged alone, and both substantially different from the levels of behavior observed when these dolphins foraged with their calves present. This suggests that the change in behavior was not merely a social phenomenon seen in the presence of other individuals, but instead reflects teaching behavior targeted at a naïve observer. However, the naïve observers in this study were presumably the calves of the observed mothers. Future research is needed to explore if these same mothers or other alloparents also exhibit similar altered foraging behavior when in the presence of other naïve observers that are not their offspring.
This altered foraging behavior on behalf of the mothers may be a valuable social learning mechanism for Atlantic spotted dolphins. The mothers in the study clearly altered their typical foraging behavior in the presence of their calves at a potential cost to themselves due to the exaggerated behavior, as well as sometimes foregoing ingestion of the prey. However, as per Caro and Hauser’s definition of teaching, the observer must benefit from the model’s altered behavior, in this case through more rapid or skillful acquisition of foraging behavior. In dolphin society this skill would be essential, as mothers give birth approximately every 3 years, at which time the older calf would become weaned and rely more on independently caught fish. It is advantageous for the mother to ensure the success of her offspring, presumably by investing her time and energy into the calf’s foraging capabilities before the calf is weaned. Weaning is a gradual process in spotted dolphins, and although the calves were not dependent upon fish for food, consuming the fish may have been an important part of the development of the calves’ foraging behavior, perhaps because it reinforced the social-learning process.
Future research is warranted to explore the development and skillfulness of the young calf’s pre-weaning foraging capabilities in order to examine the full effect of the mother’s presumed teaching efforts. If this is true teaching behavior, the calf will derive some benefit from observing the mother’s altered foraging behavior. Additionally, a comparison is needed to compare the calves of teaching and non-teaching or less attentive mothers. However, because all of the mothers observed in this study demonstrated the altered foraging behavior, it may be difficult to assess the consequences of naturally occurring individual differences in the mothers’ foraging behavior on the calves. Although the full benefit for the calf is unknown, it seems that the calf is indeed the target of this altered behavior, which was observed only in the presence of the calf.
Additionally, further research is needed into the calf’s behavior and attention to the mother. Future research can hopefully clarify if the calf is indeed attentive to the mother’s potential teaching behavior to gain something from the experience, as well as clarify what mechanism the calves may be using to learn from the mothers, such as imitation, stimulus enhancement, or local enhancement. Data from this study, such as the calves being predominantly in the observation position relative to the mother during the events, some of the calves chasing and ingesting the fish, and the calves’ interactions with both the mothers and prey objects, may be evidence that the calves were attentive to the mothers’ altered foraging behavior and support the argument for teaching. Future data from the calf behavior will also hopefully strengthen our argument that the altered maternal foraging behavior may be an example of teaching. The data from the calf behavior are currently being collected and analyzed as part of a separate ongoing project and will be available for future publication. Despite this drawback, we believe that this study detailing the altered foraging behavior of the mothers is a significant finding in the area of animal cognition, even without data on the calf behavior.
Despite previous research that has shown that dolphins pass mirror self-recognition tests (Reiss and Marino 2001), understand referential pointing (Herman et al. 1999), and spontaneously use referential gesturing in captive situations (Xitco et al. 2001), further evidence is needed before attributing theory of mind to dolphins, which some researchers argue is required for true teaching (Tomasello et al. 1993). Regardless of the lack of conclusive evidence supporting theory of mind in dolphins, the perspective-taking abilities by dolphins supported by previous research (Herman et al. 1999; Xitco et al. 2001) might be sufficient for the presumed teaching behavior shown here. Although the cognitive abilities behind the clear alteration of foraging behavior of the mother dolphins in the presence of their calves are yet to be determined, the observed teaching behaviors in dolphins are nonetheless remarkable. Mother dolphins provide opportunities for calves to observe foraging behaviors, and sometimes even provide opportunities for calves to practice foraging. Teaching, then, may be an important way in which aspects of cetacean social learning and possibly culture are transmitted from one generation to the next.
Notes
One mother, Little Gash, was observed foraging without her calf present both as an adult and as a juvenile. When group means were compared including Little Gash there were no significant differences between adults and juveniles for either chase latencies, t (8) = 0.05, P = 0.96, or body-orienting movements, t (8) = 0.41, P = 0.69. Both the mean chase latencies, M A = 3.67 and M J = 6.89, and mean number of body-orienting movements, M A = 0.5 and M J = 1.0, for Little Gash were comparable as an adult and as a juvenile.
It is important to note that research in captivity (Pack and Herman 1995) has demonstrated the ability of dolphins to perceive and recognize objects through either vision or echolocation. In addition, their perceptions are readily shared or integrated across the senses, regardless of which modality the dolphin originally perceived the external stimuli. However, the sounds emitted from the dolphins were not measured in the present study, but should be looked at in future research in order to determine what sensory information the calf is receiving.
References
Bering JM (2001) Theistic percepts in other species: can chimpanzees represent the minds of non-natural agents? J Cogn Cult 1:107–137
Bering JM, Povinelli DJ (2003) Comparing cognitive development. In: Maestripieri D (ed) Primate psychology. Harvard University Press, Cambridge, pp 205–233
Bjorklund DF, Bering JM (2003) Big brains, slow development, and social complexity: the developmental and evolutionary origins of social cognition. In: Brüne M, Ribbert H, Schiefenhövel W (eds) The social brain: evolutionary aspects of development and pathology. Wiley, New York, pp 133–151
Bjorklund DF, Yunger JL, Bering JM, Ragan P (2002) The generalization of deferred imitation in enculturated chimpanzees (Pan troglodytes). Anim Cogn 5:49–58
Boesch C (1991) Teaching in wild chimpanzees. Anim Behav 41:530–532
Boesch C (1993) Toward a new image of culture in chimpanzees. Behav Brain Sci 16:514–515
Boesch C, Tomasello M (1998) Chimpanzee and human cultures. Curr Anthropol 39:591–614
Caro TM, Hauser MD (1992) Is there teaching in nonhuman animals? Q Rev Biol 67:151–174
Greenfield P, Maynard A, Boehm C, Schmidtling EY (2000) Cultural apprenticeship and cultural change: tool learning and imitation in chimpanzees and humans. In: Parker ST, Langer J, McKinney ML (eds) Biology, brains, and behavior: the evolution of human development. School of American Research Press, Santa Fe, pp 237–277
Guinet C, Bouvier J (1995) Development of intentional stranding hunting techniques in killer whale (Orcinus orca) calves at Crozet Archipelago. Canadian Journal of Zoology 73:27–33
Herman LM (2002) Vocal, social, and self-imitation by bottlenosed dolphins. In: Nehaniv C, Dautenhahn K (eds) Imitation in animals and artifacts. MIT Press, Cambridge, pp 63–108
Herman LM, Abichandani SL, Elhajj AN, Herman EYK, Sanchez JL, Pack AA (1999) Dolphins (Tursiops truncatus) comprehend the referential character of the human pointing gesture. J Comp Psychol 113:347–364
Herzing DL (1996) Vocalizations and associated underwater behavior of free-ranging Atlantic spotted dolphins, Stenella frontalis, and bottlenose dolphins, Tursiops truncatus. Aquat Mamm 22:61–79
Herzing DL (1997) The life history of free-ranging Atlantic spotted dolphins (Stenella frontalis): age classes, color phases an female reproduction. Mar Mamm Sci 13:576–595
Herzing DL (2005) Transmission mechanisms of social learning in dolphins: underwater observations of free-ranging dolphins in the Bahamas. In: Delfour F (ed) Autour de L’ethologie et de la cognition animale. Presses Universitaires de Lyon, Lyon, pp 185–194
Herzing DL, Brunnick BJ (1997) Coefficients of association of reproductively active female Atlantic spotted dolphins, (Stenella frontalis). Aquat Mamm 23:155–162
Kruetzen M, Mann J, Heithaus M, Connor R, Bejder L, Sherwin WB (2005) Cultural transmission of tool use in bottlenose dolphins. Proc Natl Acad Sci USA 105:8939–8943
Kuczaj SAII, Paulos RD, Ramos JA (2005) Imitation in apes, children and dolphins: implications for the ontogeny and phylogeny of symbolic representation. In: Namy L (ed) Symbol use and symbolic development. MIT Press, Cambridge, pp 221–243
Mann J, Smuts BB (1999) Behavioral development in wild bottlenose dolphin newborns (Tursiops sp.). Behaviour 136:529–566
Marino L, Connor RC, Fordyce RE, Herman LM, Hof PR, Lefebvre L, Lusseau D, McCowan B, Nimchinsky EA, Pack AA, Rendell L, Reidenberg JS, Reiss D, Uhen MD, Van der Gucht E, Whitehead H (2007) Cetaceans have complex brains for complex cognition. PLoS Biol 5:966–972
Miles JA, Herzing DL (2003) Underwater analysis of the behavioural development of free-ranging Atlantic spotted dolphin (Stenella frontalis) calves (birth to 4 years of age). Aquat Mamm 29:363–377
Pack AA, Herman LM (1995) Sensory integration in the bottlenosed dolphin: immediate recognition of complex shapes across the senses of echolocation and vision. J Acoust Soc Am 98:722–733
Pack AA, Herman LM (2004) Dolphins (Tursiops truncatus) comprehend the referent of both static and dynamic human gazing and pointing in an object choice task. J Comp Psychol 118:160–171
Reiss D, Marino L (2001) Self-recognition in the bottlenose dolphin: a case of cognitive convergence. Proc Natl Acad Sci USA 98:5937–5942
Rendell L, Whitehead H (2001) Culture in whales and dolphins. Behav Brain Sci 24:309–382
Spininelli LHP, Jesus AH, Nascimento LF, Yamamoto ME (2006) Prey-transfer in the marine tucuxi dolphin, Sotalia fluviatilis, on the Brazilian coast. JMBA2, Biodiversity Records (published online)
Thornton A, McAuliffe K (2006) Teaching in wild meerkats. Science 313:227–229
Tomasello M (1996) Do apes ape? In: Heyes C, Galef B (eds) Social learning in animals: the role of culture. Academic Press, San Diego, pp 319–346
Tomasello M (2000) Culture and cognitive development. Curr Dir Psychol Sci 9:37–40
Tomasello M, Kruger AC, Ratner HH (1993) Cultural learning. Behav Brain Sci 16:495–552
Whiten A (2005) The second inheritance system of chimpanzees and humans. Nature 437:52–55
Whiten A, Goodall J, McGrew WC, Nishida T, Reynolds V, Sugiyama Y, Tutin CEG, Wrangham RW, Boesch C (1999) Cultures in chimpanzees. Nature 399:682–685
Xitco MJ, Gory JD, Kuczaj SAII (2001) Spontaneous pointing by bottlenose dolphins (Tursiops truncatus). Anim Cogn 4:115–123
Acknowledgments
We would like to thank Stan Kuczaj and Jesse Bering for their helpful comments on earlier drafts of this manuscript, and John Bender, Miley Fishero, Megan Rothrock, Melissa Ingui, Wisline Shepherd, Sheryl Spencer, and the Wild Dolphin Project for their assistance on the project.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Movie clip S1 (MPG 5925 kb)
Movie clip S2 (MPG 9443 kb)
Rights and permissions
About this article
Cite this article
Bender, C.E., Herzing, D.L. & Bjorklund, D.F. Evidence of teaching in atlantic spotted dolphins (Stenella frontalis) by mother dolphins foraging in the presence of their calves. Anim Cogn 12, 43–53 (2009). https://doi.org/10.1007/s10071-008-0169-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-008-0169-9