
Abstract
There is increasing evidence for cultural variations in behaviour among non-human species, but human societies additionally display elaborate cumulative cultural evolution, with successive generations building on earlier achievements. Evidence for cumulative culture in non-human species remains minimal and controversial. Relevant experiments are also lacking. Here we present a first experiment designed to examine chimpanzees’ capacity for cumulative social learning. Eleven young chimpanzees were presented with a foraging device, which afforded both a relatively simple and a more complex tool-use technique for extracting honey. The more complex ‘probing’ technique incorporated the core actions of the simpler ‘dipping’ one and was also much more productive. In a baseline, exploration condition only two subjects discovered the dipping technique and a solitary instance of probing occurred. Demonstrations of dipping by a familiar human were followed by acquisition of this technique by the five subjects aged three years or above, whilst younger subjects showed a significant increase only in the elements of the dipping technique. By contrast, subsequent demonstrations of the probing task were not followed by acquisition of this more productive technique. Subjects stuck to their habitual dipping method despite an escalating series of demonstrations eventually exceeding 200. Supplementary tests showed this technique is within the capability of chimpanzees of this age. We therefore tentatively conclude that young chimpanzees exhibit a tendency to become ‘stuck’ on a technique they initially learn, inhibiting cumulative social learning and possibly constraining the species’ capacity for cumulative cultural evolution.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Social learning offers a potentially adaptive way to avoid the costs of individual (e.g. trial and error) learning, especially where complex tasks or changing environments may be involved (Laland 2004). However, social learning may also carry risks. In a species where social learning plays a particularly important role one can postulate situations in which a maladaptive, or at the very least a suboptimal strategy, will spread and be maintained throughout a community (Galef 1995, Laland 1996, 2004; Giraldeau et al 2002). A capacity to recognize instead that a modification of a known behaviour being used by another individual is more productive or effective in obtaining results than one’s own, and the flexibility to switch to this alternative behaviour, may be at the core of what Tomasello (1994) calls the ‘ratchet effect’, whereby incremental improvement in behaviour occurs in succeeding generations. Incremental ratcheting is evident in humans (Mesoudi et al. 2006) but its existence in non-humans remains controversial (McGrew 2004).
Chimpanzees have been shown to learn a variety of behaviours by observing either a conspecific or a human model in captivity (see Whiten et al. 2004, for a recent review) and there is evidence for socially transmitted behavioural variations in the wild (Whiten et al 1999, 2001). Together these lines of evidence suggest that chimpanzees qualify as a species particularly inclined to learn from observation, potentially exposing them to the risk of socially acquiring and maintaining maladaptive as well as adaptive behaviours.
In the wild, use of differently productive behaviours to obtain the same food source have been observed, with the Gombe ant-dipping technique reported to be more productive than the Tai technique (McGrew 1974; Boesch and Boesch 1990; Boesch 1996; Humle and Matsuzawa 2002). However in a wild population it is difficult to exclude the possibility that environmental factors are responsible for differentially productive foraging strategies (Galef 1990; Tomasello 1990; Whiten and Ham 1992). There is evidence that this is the case for some of the differing ant-dipping techniques observed in the wild (Humle and Matzsuawa 2002).
Recent research in a controlled setting has highlighted chimpanzees’ inclination to conform to local ‘cultural norms’ (Whiten et al. 2005) and their tendency to maintain arbitrary, culturally spread conventions (Bonnie et al. 2007). Conformity to local norms may go some way to explaining why neighbouring communities sharing the same environment may maintain different social conventions (McGrew et al 2001; Nakamura and Uehara 2004) and foraging techniques (Humle and Matzusawa 2002). Conformity could also represent a powerful mechanism in the maintenance of a maladaptive or suboptimal strategy. On the other hand, the cognitive sophistication shown by chimpanzees in the adaptive use of social learning mechanisms including imitation and emulation (see Horner and Whiten 2005) and the flexibility shown in the use and modification of tools in natural foraging contexts, such as the use of multiple tool sets for termite fishing (Sanz et al. 2004) and the use of stones as props for anvils in nut cracking (Matsuzawa 1994; Sugiyama 1997), may hint at the possibility of simple forms of cumulative culture.
To our knowledge, no experimental study has as yet directly addressed the question of chimpanzees’ capacity for cumulative social learning. In the current study, a tool-based foraging task, which could be solved using either a relatively simple technique, or a more elaborate and more productive technique, was introduced to young chimpanzees. After a number of baseline exploration trials participants witnessed a model performing the simple, suboptimal strategy. We measured whether chimpanzees’ subsequent performance of this technique exceeded baseline levels, and occurred more frequently than the alternative, more complex, technique. Subjects that acquired the first, simple technique, then witnessed the more rewarding technique, built cumulatively by adding an extra element to the existing one. The question we then addressed was: Would the chimpanzees switch to the more elaborate and successful technique or would they stick to what they had previously learned?
Methods
Subjects
The study was carried out in the Ngamba Island Chimpanzee Sanctuary in Uganda, which houses orphaned chimpanzees confiscated by the Ugandan Wildlife authorities following illegal poaching activities (see Horner and Whiten 2005 for more details).
Eleven chimpanzees comprised the experimental group, five females and six males, aged 2–7 years (Table 1). The age of the chimpanzees was determined on first arrival by the sanctuary veterinarian based largely on dental eruption.
Materials
The dipping task
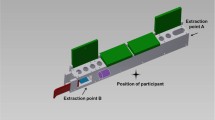
In all baseline and experimental trials the participants were presented with a ‘honey-dip’ array and two identical tools (rods) appropriate for performing both a dipping and a probing technique. The honey-dip array (HD) consisted of nine separate units fixed to two wooden trays (thus forming one array of five honey pots and another of four honey pots). All the honey-pots were opaque. Each honey pot measured 9 × 6 × 6 cm.
During the social learning trials, participants witnessed a familiar human gaining the food using one of two methods:
Dipping-technique: The model used the index finger to slide open a small trap door and, whilst holding it open, used the other hand to insert a rod to reach the honey, a small amount of which could be licked off the tool after withdrawal (Fig. 1a).

Foraging techniques. a Dipping technique demonstrated by the human model, b probing technique, removal of the bolt, c probing technique, levering open of the lid
Probing-technique: Using the same rod, the model poked a small, recessed bolt in the side of each honey-pot inwards, thus allowing the lid (in which the smaller trap door is set) to be opened (Fig. 1b). After having poked the bolt, the model opened the trap door with a finger and, whilst holding it open, used the other hand to insert the rod and lever the larger lid open (Fig. 1c), making all the contents available. Thus to lever the lid open, a variation of the dipping technique was incorporated, making the probing technique one that could be built cumulatively once the dipping technique is already in an animal’s repertoire.
The apparatus contained both honey and peanuts; thus whereas the dipping-technique did not allow participants to obtain the peanuts and numerous insertions were required to gain all the honey content, the probing-technique allowed rapid access to both honey and peanuts. Both peanuts and honey are highly prized food sources with which the chimpanzees were already familiar prior to testing.
Procedure
The procedure consisted of (a) three baseline trials, followed by (b) four demonstration/test trials of the dipping-technique, then (c) four demonstration/test trials of the probing-technique and (d) further and more extensive demonstrations of the probing-technique and subsequent testing (details below). Only the chimpanzees who had successfully acquired the dipping-technique were exposed to the remaining conditions (Table 2). All chimpanzees were tested individually, out of sight of their peers.
Baseline trials
Three individual exploration trials lasting 1 h each were conducted with each participant on consecutive days. In these trials the tool was placed within 1.5 m of the apparatus. On entering the testing room, the researcher simulated food-grunts next to the baited apparatus, thus drawing attention to the novel object.
Demonstration of the dipping technique
Demonstration trials of the dipping technique were presented on four consecutive days. Daily sessions lasted 1 h with a set of demonstrations being carried out on first entering the room and then every 15 min. A set of demonstrations consisted of the model dipping into all the five honey pots on the tray. Demonstrations were given by the researcher (S.M-P) to one chimpanzee at a time and, to mimic as far as possible the naturalistic setting in which social learning of tool-use normally occurs, participants were allowed to approach and watch from as close a position as s/he wished. The researcher ate some of the honey and occasionally shared it with the chimpanzee. After each set of demonstrations, the researcher positioned herself 1 m from the apparatus and video recorded the chimpanzees’ interactions with the apparatus.
Demonstration of the optimal ‘probing’ technique to ‘dipping’ chimpanzees
Chimpanzees who acquired the dipping technique were subsequently exposed to demonstration trials of the probing technique (Phase 1) presented on four consecutive days. Daily sessions lasted 1 h, with a set of demonstrations being carried out on first entering the room and then every 15 min. A set of demonstrations consisted of the model using the probing-technique to open an array of five honey pots. The model performed the probing-technique on a separate honey-dip tray (placed next to the chimpanzees’ tray) to allow refilling of the apparatus between demonstrations. The researcher simulated pant-grunts on opening the lids and exposing the entire honey and nut content, and shared part of the food with the chimpanzee. In all other respects the procedure was identical to the demonstration trials described above.
If chimpanzees failed to acquire the probing technique, they were further exposed to a human model carrying out two sets of demonstrations of the probing technique every 30/40 min for a total of 30 sets of demonstrations a day (Phase 2). The demonstrations were carried out in front of the whole group, but the chimpanzees were not permitted to approach the apparatus. Demonstrations were carried out on three consecutive days and on the fourth day the chimpanzees were tested individually. A further cycle of three consecutive days of demonstrations, was followed by a second individual testing session on the fourth day.
Behavioural coding
Behaviours were coded as separate actions only if participants broke contact with the device before beginning a new action. A detailed coding of all the participants’ actions on the HD was carried out. For the purpose of analysis, discrete behaviours were allocated to two major categories corresponding to either the ‘dipping technique’ or the ‘probing technique’ and two further categories comprising ‘elements’ of the two techniques (Table 3). Because of the physical separation of target areas, coding of the two methods was straightforward and unambiguous and was thus carried out by a single observer.
Statistical analysis
Given the sample size, non-parametric statistical tests were used to analyse the data and within group comparisons between conditions were conducted (Wilcoxon tests). We followed the recommendation of Mundry and Fischer (1998) to avoid reliance on Z values when N < 15, instead using the raw Wilcoxon T + values and associated probabilities. Reported P values are one-tailed (Siegel and Castellan 1988) since predictions are clearly uni-directional (that social learning will enhance success rates utilising the method witnessed).
Results
Baseline trials: dipping versus probing
In baseline trials 2 of the 11 chimpanzees (Mawa and Ikuru) discovered the dipping technique, doing so in the first trial. Both chimpanzees performed a single continuous movement in which the rod was used to open the trap door then inserted into the hole to obtain the honey. One of these chimpanzees, Mawa, also discovered the probing technique in the second trial, although he performed the whole technique only once.
Demonstration trials of the dipping technique
After demonstrations, three further chimpanzees above three years of age acquired the dipping technique successfully and did so either during the first trial (two chimpanzees: Nkumwa and Cindy) or the second (Kalema).
A within subject analysis for the five older chimpanzees (including Mawa and Ikuru who had mastered a one-handed version of the dipping technique during baseline trials) showed a significant increase in the frequency of dipping technique from baseline (median 7.2) to demonstration trials (median 88.28) (Wilcoxon test: N = 5, T + = 0, P = 0.03) (Fig. 2). The frequency of the dipping technique was significantly greater than that of probing (median dipping 83.4 vs. median probing 0; Wilcoxon test: N = 5, T + = 0, P = 0.03). However, the behaviour adopted to carry out the dipping technique varied amongst the chimpanzees. Cindy immediately adopted a non-demonstrated, one-handed technique, where the tool was used to open the slide door by directly inserting it into the pot. This is the same variant of the dipping technique used by the two chimpanzees who independently discovered this method of accessing the honey during baseline trials (Mawa and Ikuru). Mawa and Ikuru continued performing the one-handed technique they adopted during baseline trials, despite demonstrations of the two-handed method. The other two chimpanzees (Kalema and Nkumwa) used the two-handed dipping method as demonstrated. However, Nkumwa rapidly changed to using the one-handed variant, whereas Kalema continued using the observed bi-manual technique to the exclusion of the other method.

Increase in the frequency of dipping technique for the older subjects from baseline to demonstration trials
The younger chimpanzees did not successfully acquire the dipping technique. However, there was a significant increase in the occurrence of elements of the dipping technique (i.e. opening the trap door with finger or tool or tapping the tool on the closed trap door) between baseline (median 0) and demonstration trials (median 13.2) (Wilcoxon test: N = 6, T + = 0, p = 0.016) (Fig. 3).

Increase in the frequency of elements of the dipping technique for younger subjects (who did not acquire the whole dipping technique) from baseline to demonstration trials
Demonstration trials of the optimal ‘probing’ technique for ‘dipping’ chimpanzees
Only the older group of chimpanzees who had successfully acquired the dipping technique were shown the more rewarding probing technique. Of the five chimpanzees showing the dipping technique only one (Mawa) successfully switched to the optimal probing behaviour, and did so after the first session of demonstrations. However, he had performed this technique once in the baseline trials. None of the remaining four chimpanzees switched to the more rewarding probing method.
A within-subjects analysis comparing dipping behaviour in dipping (median 88.3) and probing (median 103.2) demonstration trials revealed no decrease in the frequency of dipping behaviour (Wilcoxon test: N = 5, T + = 3, P = 0.16).
Four of the five chimpanzees never performed the probing technique either in baseline or in the last two trials, negating further statistical testing. This was not true of the frequency of elements of the probing technique, where a comparison of baseline versus the last two trials revealed a small, non-significant increase (from a median of 0 to a median of 3.5; Wilcoxon test: N = 5, T + = 1, P = 0.06).
Discussion
We investigated whether chimpanzees would observationally learn a new foraging technique, and whether they would then persist in its use after repeated exposure to a model performing a substantially more efficient technique built cumulatively on the first, or would instead ‘upgrade’ to this improved technique. We found evidence of social learning in the acquisition of the dipping technique, but overall, once the chimpanzees had learned the first technique they failed to switch to the more efficient strategy, despite extensive demonstrations.
A plausible explanation for this conservatism is that as the dipping technique became habitual, it inhibited these young chimpanzees’ perception and assimilation of the more advanced technique. However, an alternative hypothesis that must be considered is that the probing technique was simply too difficult for chimpanzees of this age to master, through either individual or social learning. Fortunately, after the main study was completed, a further three chimpanzees, above 3-years of age became available for testing. Although not providing a statistically robust sample, these youngsters provided the opportunity to examine whether different outcomes would emerge if the simpler technique was made to be non-functional, so only the use of the more advanced technique would allow chimpanzees to obtain the food. We engineered this by simply presenting the task containing only peanuts and no honey, following the same design of baseline and social learning phases. However, two of these three chimpanzees explored the task fully, discovering both the dipping technique and the probing technique during the baseline trials, thus rendering the presentation of the social learning trials redundant. Acquisition of the probing technique occurred in the first trial for one chimpanzee and in the third trial for the other. These responses demonstrate that without the prior habit of dipping for honey, discovery of the probe technique is not so very challenging for chimpanzees of this age.
That chimpanzees did not switch because they were simply occupied by dipping and did not watch demonstrations of the probing technique can also be dismissed, because during probing demonstrations the task was not made available.
Aside from Mawa who did carry out a complete switch, eventually exclusively performing the probing technique to obtain the combined honey and nut resource, in all the other chimpanzees there was only an appearance of a few elements of the probing technique (i.e. attempts at inserting the rod in the recessed bolt placed on the side of the honey-pots) during the latter two trials (thus after over 200 demonstrations by the model). As highlighted by the lack of statistically significant increase in this behaviour, these attempts were infrequent and overshadowed by the persistence in the use of the dipping technique. It is of course possible that with yet more demonstrations and more experience with the apparatus, the chimpanzees would have eventually upgraded to the more efficient probing task as Mawa did. However, what is highlighted by this study is the chimpanzees’ difficulty in doing so as compared to their relative ease in socially acquiring a novel tool-using behaviour.
We suggest our results are best interpreted in relation to two of the scenarios that Laland (2004) considered in an analysis of the conditions that might motivate animals to adopt a social learning as opposed to an individual learning strategy. One of these would be a copy-if-better approach, whereby an individual would be inclined to copy a behaviour, which is perceived as more successful than ones own. As noted earlier this is considered the basis of the ratchet effect (Tomasello 1994), whereby incremental improvements in behaviour may occur from one generation to the next. The copy-if-better strategy however, is cognitively demanding insofar as it requires a comparison between the outcome of another’s and one’s own actions. Laland postulates a simpler yet still potentially efficient alternative: the copy-when-dissatisfied scenario. In this case the strategy requires reference only to one’s own internal state. Our results seem to lend support to the use of only a copy-when-dissatisfied strategy in young chimpanzees, in that whereas social learning of the dipping technique did take place when the chimpanzees were unable to discover the technique individually and thus were not obtaining a reward at all (presumably accompanied by dissatisfaction), social learning of the more productive method did not occur when at least some reward was obtainable by the chimpanzees’ existing strategy. Of course this first study does not show that chimpanzees are unable to use the copy-if-better strategy. A potentially interesting development of our study, to dissect the motivation issue more closely, would be to modify the reward and/or tool being used to reduce the effectiveness/value of the dipping method, and thus monitor if and when the switch to the more rewarding probing strategy might occur.
The one chimpanzee (Mawa) who did switch techniques was one of the two chimpanzees who had learned the dipping behaviour during the individual exploration trials, then also successfully showed the probing technique, albeit only once. It is thus likely that the actions he eventually observed when the model performed the probing technique were easier to assimilate and reproduce because already familiar to him (‘response facilitation’, Byrne 1994).
The difficulty in switching behaviours observed in this study is in line with research in the wild suggesting a degree of conservativism in chimpanzees (Kummer and Goodall 1985; Takahata et al 1986; Boesch 1995). A particularly suggestive example evidenced in a study by Matsuzawa (1996) highlights the apparent difficulties in incorporating new elements into an already familiar behaviour. A new species of nut (Coula edulis) was presented to a wild chimpanzee community already adept at cracking open oil palm nuts. Although most of the chimpanzees examined the Coula nuts, only one female (Yo) carried out the appropriate behaviour. Authors suspected Yo may have originated from a neighbouring Coula-nut-cracking community, although this could not be confirmed. During the following days, only two juvenile chimpanzees (aged 5 and 6) acquired the behaviour and, whereas the adults tended to ignore Yo’s behaviour, youngsters were seen to observe her more closely. Over the following ten year period, the cracking of Coula nuts did eventually spread throughout the entire community, with the juveniles being more proactive in this process and the adults showing a slower acquisition of the novel food source (Biro et al. 2003). A more rapid switch was seen in a study carried out in captivity where chimpanzees adept at dipping straws to obtain milk from a pot, switched to using the straw as a drinking tool after observing a model (Spiezio, unpublished data). As in the wild (Biro et al 2003), juveniles were the first to acquire this new behaviour. Results from this study contrast with our own, since the chimpanzees upgraded to a more rewarding method relatively quickly, despite previous familiarity with the less efficient technique. A possible reason for this discrepancy is that in Spiezio’s study the new technique was not directly building on an existing one; rather, the two behaviours might be perceived as distinctly different strategies.
Conclusion
Chimpanzees in our experiment displayed a striking lack of acquisition of an optimal and cumulatively built technique they witnessed, once they were habitually using a simpler technique that worked. Our supplementary experiment, even though with only three chimpanzees, suggested that the more elaborate technique was comfortably within the chimpanzees’ grasp if they had been motivated to assimilate it. The conservatism implied by our results suggests one explanation for the lack of evidence for cumulative cultural evolution in our nearest relative that contrasts so markedly with human cultural history. However, ours is but a single, exploratory study of this phenomenon. Further studies are needed employing other kinds of contrast between simpler and more cumulatively elaborated versions of functional techniques before the generality of the conclusions appropriate to our study can be established.
References
Biro D, Inoue-Nakamura N, Tonooka R, Yamakoshi G, Sousa C, Matsuzawa T (2003) Cultural innovation and transmission of tool use in wild chimpanzees: evidence from field experiments. Anim Cogn 6:213–223
Boesch C (1995) Innovation in wild chimpanzees (Pan troglodytes). Int J Primatol 16(1):1–15
Boesch C (1996) The emergence of culture among wild chimpanzees? Behav Brain Sci 16:514–515
Boesch C, Boesch H (1990) Tool use and tool making in wild chimpanzees. Folia Primatol 54:86–99
Bonnie KE, Horner V, Whiten A, de Waal FBM (2007) Spread of arbitrary customs among chimpanzees: a controlled experiment. Proc R Soc B 274:367–372
Byrne RW (1994) The evolution of intelligence. In: Slater JB, Halliday TR (eds) Behaviour and evolution. Cambridge University Press, Cambridge
Galef BG (1990) Traditions in animals: field observations and laboratory analyses. In: Bekoff M, Jamieson D (eds) Interpretation and explanation in the study of animal behaviour. Westview Press, Boulder, pp 74–95
Galef BG (1995) Why behaviour patterns that animals learn socially are locally adaptive. Anim Behav 49:1325–1334
Giraldeau L-A, Valone TJ, Templeton JJ (2002) Potential disadvantages of using socially acquired information. Phil Trans R Soc 357:1559–1566
Horner V, Whiten A (2005) Causal knowledge and imitation/emulation switching in chimpanzees (Pan troglodytes) and children. Anim Cog 8:164–181
Humle T, Matsuzawa T (2002) Ant dipping among chimpanzees of Bossou, Guinea and some comparisons with other sites. Am J Primatol 58(3):133–148
Kummer H, Goodall J (1985) Conditions of innovative behaviour in primates. Phil Trans R Soc 308:203–214
Laland K (1996) Is social learning always locally adaptive? Anim Behav 52:637–640
Laland K (2004) Social learning strategies. Learn Behav 32(1):4–14
Matsuzawa T (1994) Field experiments on use of stone tools by chimpanzees in the wild. In: Wrangham RW, Mcgrew WC, de Waal FBM, Heltne P (eds) Chimpanzee cultures. Harvard University Press, Cambridge, pp 351–370
Matsuzawa T (1996) Chimpanzee intelligence in nature and in captivity: isomorphism of symbol use and tool use. In: McGrew WC, Marchant LF, Nishida T (eds) Great ape societies. Cambridge University Press, Cambridge
McGrew WC (1974) Tool use by wild chimpanzees in feeding upon driver ants. J Hum Evol 3:501–508
McGrew WC, Marchant LF, Scott SE, Tutin CEG (2001) Intergroup differences in a social custom in wild chimpanzees. The grooming handclasp of the Mahale Mountains. Curr Anthropol 42:148–153
McGrew WC (2004) The cultured chimpanzee: reflections on cultural primatology. Cambridge University Press, Cambridge
Mundry R, Fischer J (1998) Use of statistical programs for nonparametric tests of small samples often leads to incorrect P values: examples from Animal Behaviour. Anim Behav 56:256–259
Mesoudi A, Whiten A, Laland KN (2006) Towards a unified science of cultural evolution. Behav Brain Sci 29:329–383
Nakamura M, Uehara S (2004) Proximate factors of different types of grooming hand-clasp in Mahale chimpanzees: implications for chimpanzee social customs. Curr Anthropol 45:108–114
Sanz C, Morgan D, Glick S (2004) New insights into chimpanzees, tools and termites from the Congo Basin. Am Nat 164:567–581
Siegel S, Castellan NJ (1988) Nonparametric statistics for the behavioral sciences. McGraw-Hill, New York
Sugiyama Y (1997) Social tradition and the use of tool-composites by wild chimpanzees. Evol Anthropol 6:23–27
Takahata Y, Hiraiwa-Hasegawa M, Takasaki H, Nyundo R (1986) Newly acquired feeding habits among the chimpanzees of the Mahale Mountians National Park, Tanzania. Hum Evol 1:277–284
Tomasello M (1990) Cultural transmission in the tool use and communicatory signalling of chimpanzees? In: Parker S, Gibson K (eds) Language and intelligence in monkeys and apes: comparative developmental perspectives. Cambridge University Press, Cambridge
Tomasello M (1994) The question of chimpanzee culture. In: Wrangham R, McGrew W, de Waal F, Heltne P (eds) Chimpanzee cultures. Harvard University Press, Cambridge
Whiten A, Goodall J, Mc Grew WC, Nishida T, Reynolds V, Sugiyama Y, Tutin CEG, Wrangham RW, Boesch C (1999) Cultures in chimpanzees. Nature 399:682–685
Whiten A, Goodall J, Mc Grew WC, Nishida T, Reynolds V, Sugiyama Y, Tutin CEG, Wrangham RW, Boesch C (2001) Charting cultural variation in chimpanzees. Behaviour 138:1481–1516
Whiten A, Ham R (1992) On the nature and evolution of imitation in the animal kingdom: reappraisal of a centuary of research. In: Slater PJ, Rosenblatt JS, Beer C, Milinski M (eds) Advances in the study of behaviour. Academic Press, San Diego
Whiten A, Horner V, Litchfield CA, Marshall-Pescini S (2004) How do apes ape? Learn Behav 32:36–52
Whiten A, Horner V, de Waal FBM (2005) Conformity to cultural norms of tool use in chimpanzees. Nature 437:737–740
Acknowledgments
We gratefully acknowledge the Trustees of the Chimpanzee Sanctuary and Wildlife Conservation Trust (Born Free, International Foundation for Animal Welfare, Jane Goodall Institute Germany, Ugandan Wildlife Education Centre, Zoological Parks Board of New South Wales, Australia), the Directors, Debby Cox and Cherie Montgomery for their support throughout. Research was in compliance with the Uganda National Council for Science and Technology, whom we also thank. Particular thanks are due to the staff on Ngamba Island, without whose help this research could not have been carried out. We thank BBSRC for financial support of the study and Emanuela Prato-Previde for comments on earlier versions of the manuscript. AW was supported by a Royal Society Leverhulme Trust Senior Research Fellowship during the writing of this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Marshall-Pescini, S., Whiten, A. Chimpanzees (Pan troglodytes) and the question of cumulative culture: an experimental approach. Anim Cogn 11, 449–456 (2008). https://doi.org/10.1007/s10071-007-0135-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-007-0135-y