
Abstract
Forested wetlands are an important carbon (C) sink. Fine roots (diameter < 2 mm) dominate belowground C cycling and can be functionally defined into absorptive roots (order 1–2) and transport roots (order ≥ 3). However, effects of microtopography on the function-based fine root dynamics in forested wetlands are poorly understood. We studied fine root biomass allocation and biomass, necromass, mass loss rate, production, mortality and decomposition of absorptive and transport roots in hummocks and hollows in a coastal plain freshwater forested wetland (FFW) in the southeastern USA using dynamic-flow method. Biomass ratios of first- to second-order roots and absorptive to transport roots and the biomass and necromass of absorptive and transport roots were significantly higher in top 0–10 cm organic peat layer than in 10–20 cm muck and mineral layer, and were significantly higher in hummocks than in hollows. The mass loss rate, production, mortality and decomposition were significantly higher in hummocks than in hollows. Absorptive roots did not have a lower mass loss rate than transport roots. Microtopography significantly affected the contributions of absorptive and transport roots to the total production, mortality and decomposition. Production, mortality and decomposition of absorptive roots were higher than those of transport roots in hummocks but lower than those of transport roots in hollows. Total (hummocks plus hollows) fine root production, mortality and decomposition were 455 ± 106 g m−2 y−1, 475 ± 79 g m−2 y−1 and 392 ± 60 g m−2 y−1, respectively. Greater mortality than decomposition resulted in net fine root C input to soil. The observed microtopographic controls on fine root dynamics have great implications for soil C cycling. As sea level rises, the relative area of hollows in coastal plain FFWs will increase, causing a decrease in fine root mass loss rate, biomass, production, mortality and decomposition and it is the balance of these processes that will determine future soil C storage and cycling.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Highlights
-
Absorptive (AR) and transport root (TR) dynamics in a coastal forested wetland were studied.
-
Soil depth and microtopography significantly affected biomass allocation, biomass and necromass.
-
Microtopography significantly changed AR and TR production, mortality and decomposition.
Introduction
Freshwater forested wetlands (FFWs), which have a broad distribution along US Gulf and Atlantic coasts (Ensign and Noe 2018), coastlines of Europe, Central America and the Amazon (Duberstein and others 2014), are important for mitigating rising atmospheric CO2 due to their large carbon (C) storage (Mitsch and Gosselink 1993; Bridgham and others 2006) and C sequestration capacity (Krauss and others 2018). Hummocks and hollows are a unique feature in most of the FFWs (Belyea and Clymo 2001; Miao 2013). Hummocks are slight microtopographic variation and usually above water table, while hollows are submerged for most of the year (Miao 2013). Differences in this microtopography dramatically change local hydrology and soil aeration, leading to differences in soil C cycling processes such as soil respiration (Miao 2013, Miao and others 2017; Krauss and others 2012), CH4 production (Minick and others 2019a, b), and leaf litter and fine root decomposition (Schilling and Lockaby 2005; Stagg and others 2017).
Fine roots (diameter < 2 mm) play a key role in regulating belowground C cycling processes (Jackson and others 1997; Norby and Jackson 2000). Quantifying fine root production, mortality and decomposition is thus essential for constructing C budgets in terrestrial ecosystems. Studies on FFWs show that fine roots in hummocks have higher biomass and production (Jones and others 2000; Baker and others 2001) than those in hollows. However, fine roots in those studies were assessed based on arbitrary diameter limits rather than branching orders, which limits our ability to quantify the functional role in the belowground C cycle. The hierarchical root system is morphologically, chemically and functionally heterogeneous (Pregitzer and others 2002; Guo and others 2008) and can be functionally defined into a short-lived absorptive root pool (order 1–2) and a long-lived transport root pool (order 3–5) (McCormack and others 2015). Absorptive roots have been found to be more responsive to a changing environment (Kou and others 2018) and have a faster turnover rate (McCormack and others 2015) than transport roots. Sun and others (2012) found that estimates of fine root production based on diameter were lower than those based on the branching order. For this reason, a study based on the functional groups could better reflect fine root dynamics in this ecosystem.
The low mass loss rate (that is, decomposition rate) of fine roots is a dominant factor controlling C accumulation rate in wetlands (Bridgham and others 2006; Mitsch and others 2013). However, existing studies on fine root mass loss rates in FFWs and other wetlands have been diameter-based rather than function-based (Day and Megonigal 1993; Crawford and others 2007; Stagg and others 2017). This classification scheme would impact the results because absorptive roots have been found to decompose much more slowly than transport roots (Fan and Guo 2010; Xiong and others 2013). Fine root mass loss rate is closely related to soil water content and temperature (Day and Megonigal 1993; Ostertag and Hobbie 1999). Fine roots died in growing season would have a different mass loss pattern than those died in non-growing season because of differences in hydrology and temperature, and therefore soil microbial activities. So far, it is still unclear whether the microtopography has the same effects on the mass loss rates of absorptive and transport roots and whether seasonal changes in hydrologic regime and temperature have been confound with the microtopographic effect in FFWs.
Root traits including structure, life span and chemistry vary greatly between absorptive and transport roots (Kou and others 2015, 2018). In some tree species, there are even distinct differences in resource uptake and transport between first- and second-order roots (McCormack and others 2015). Root traits exhibit species-specific plasticity in response to changes in soil conditions such as soil water content (Megonigal and Day 1992), nutrient availability (Prieto and others 2015) and soil depth (Makita and others 2010). Increase in nitrogen (N) availability can promote the growth of absorptive roots but may have no effect on transport roots (Kou and others 2018). Assessing biomass allocation in root systems is thus important for understanding plant resource allocation strategy in response to a changing environment (Poorter and Nagel 2000; Prieto and others 2015; McCormack and others 2014). Compared with hollows, hummocks are more aerobic and have greater nitrification fluxes, and soil C and N concentrations (Noe and others 2013). In addition, soils in FFWs have two distinct layers, an organic peat layer and a highly decomposed muck and mineral sediment layer (Riggs 1996). Soil in muck and mineral layer has lower C and N concentrations and is saturated for much longer time than peat layer (Minick and others 2019a, b). These differences in nutrient and hydrologic conditions between hummocks and hollows and between organic peat layer and muck and mineral sediment layer could significantly affect fine root biomass allocation pattern at the ecosystem scale. Unfortunately, research on the effects of microtopography and soil depth on the root biomass allocation and biomass of different orders of fine roots in FFWs is almost completely lacking.
To address these questions, we investigated a FFW situated in the coastal plain of the southeastern USA (Mitsch and Gosselink 1993). Fine root biomass allocation patterns, and biomass and necromass of absorptive and transport roots were assessed using a soil coring method, while the mass loss rates of absorptive and transport roots in growing and non-growing seasons were determined using a litterbag method. We used a dynamic-flow model (Li and Lange 2015) to quantify production, mortality and decomposition (that is, amount of roots decomposed) of absorptive and transport roots. This model can give more reliable estimates than traditional soil coring methods as the concurrent fine root growth, death and decomposition processes can be better assessed by combining soil core data with results of a modified decomposition experiment. The objectives of this study were (1) to estimate fine root production, mortality and decomposition by functional group; (2) to test whether microtopography significantly affects the production, mortality and decomposition of absorptive and transport roots; (3) to test whether the effect of microtopography on mass loss rates of absorptive and transport roots is cofounded by seasonal changes in soil temperature and water contents; (4) to determine standing biomass and necromass of absorptive and transport roots in hummocks and hollows; and (5) to assess effects of microtopography and soil depth on root biomass allocation pattern in FFWs.
Materials and Method
Study Site
The study was conducted at the Alligator River National Wildlife Refuge on the Albemarle-Pamlico Peninsula of North Carolina, the USA (35°47′ N, 75°54′ W). This peninsula differs from coastlines to the north and south because of the specific combination of geomorphic features and lagoonal environment, which results in the absence of astronomic tides and rainfall being the main source of water input (Moorhead and Brinson 1995). The mean annual precipitation and temperature for the period 1981–2018 were 1272 mm and 17.0°C, respectively. The forest type is a mixed hardwood swamp forest. Black gum (Nyssa sylvatica), swamp tupelo (Nyssa biflora), and bald cypress (Taxodium distichum) are dominant species in the overstory, with occasional red maple (Acer rubrum), water oak (Quercus laurifolia), white cedar (Chamaecyparis thyoides), sweet gum (Liquidambar styraciflua) and loblolly pine (Pinus taeda). The understory is dominated by fetterbush (Lyonia lucida), bitter gallberry (Ilex galbra), sweet bay (Magnolia virginiana) and red bay (Persea borbonia). Canopy heights ranged from 15 to 20 m, and peak leaf area index was 3.5 ± 0.3 (Miao 2013, Miao and others 2017). Tree stand density was 2320 ± 800 stems ha−1, and aboveground live biomass was allometrically estimated at 37.5 ± 12.5 MgC ha−1 (Miao and others 2017). Soils at this site are acidic with a pH of 4.2–4.8 in the surface horizons. Hummocks are usually above water table, and the non-vegetated hollows are submerged for more than 70% of the year (Miao 2013). On average, hummocks and hollows account for 60% and 40% of the total area, respectively (Miao and others 2017).
Precipitation, soil temperature (Ts) at 5 cm and water table depth (WTD) were measured at an eddy covariance flux tower in this FFW (Figure 1). Meteorological data from an adjacent meteorological station, in Manteo, NC, the USA, approximately 20 km away, were analyzed to gap fill missing data. For more information, refer to Miao (2013), Miao and others (2017).

Soil temperature at 5 cm (10-day mean) and soil water table depth (WTD) (10-day mean) from Jan 2016 to Feb 2018 and starting and ending dates of the first (DE1) and second (DE2) decomposition experiments. The dark gray and black dots stand for the litterbag sampling dates in DE1 and DE2, respectively.
Soil Profile and Root Sampling
Five plots, 25 m to 300 m apart, were randomly selected in November 2015. Each plot had one hummock and one hollow, covering an approximate area of 25 m2. The soil profile consisted of a peat horizon underlain by a highly decomposed muck horizon and a highly reduced mineral sediment layer of Pleistocene origin. Our previous study showed that over 90% of fine roots (diameter < 2 mm) are distributed in the top 0–20 cm layer (unpublished data). Fine root standing biomass and necromass were assessed using a sequential soil coring method. Root samples were collected on 18 February, 15 May, 11 July, 28 September, 30 November in 2016, on 21 January, 12 April, 25 June, 8 December in 2017 and on 7 February in 2018. On each sampling occasion, three soil cores (22 mm in diameter and 20 cm in depth) were collected from each hummock and hollow, yielding 30 soil cores in total. Each core was separated into two 10-cm layers. The upper layer is mostly composed of less decomposed organic peat, and the lower layer consists of highly decomposed muck and mineral sediment. As a result, the soil cores were classified into four types: peat layer in hummocks and hollows and muck and mineral layer in hummocks and hollows.
Biomass, Necromass and Morphology
Collected samples were rinsed with clear tap water through a 0.5-mm mesh sieve to isolate roots. Live and dead fine roots (diameter < 2 mm) were separated out based on the cohesion of stele and periderm, root elasticity, and color. Generally, fine roots with light color, intact stele and periderm were regarded as live roots (Persson 1980; Hertel and Leuschner 2002).
To estimate biomass allocation factor among different types of live roots and the ratio of absorptive to transport root necromass, one or two soil cores were randomly selected from the three cores collected from both hummocks and hollows at each plot on 18 February, 15 May, 28 September in 2016, on 21 January, 25 June in 2017 and on 7 February in 2018. Live roots were separated into first-order, second-order, and transport fine roots according to the functional classification of tree fine roots (McCormack and others 2015). The combination of the first- and second-order roots was defined as absorptive roots, whereas third-order roots to roots with diameter below 2 mm were regarded as transport roots. First-order roots are the most distal, unbranched roots, and second-order roots begin at the junctions of two first-order roots, and so on (Pregitzer and others 2002). The biomass ratios of first-order to second-order roots (R1st/R2nd) and absorptive to transport roots (AR/TR) were determined.
Dead fine roots were separated into dead absorptive roots and dead transport roots using the following means. For the dead fine roots with intact rooting branches, we distinguished ARs from TRs using the method mentioned above. Some dead intact ARs were off the root branches. We identified them based on their unique morphology. First-order roots are distal and unbranched, whereas second-order roots are the next order up. For partly decomposed dead fine roots, we classified these roots based on the diameter limit separating absorptive and transport roots developed from intact dead fine roots. The small partly decomposed roots with diameters below half of the mean diameter threshold were regarded as absorptive roots. Similarly, the big partly decomposed roots having diameters below 1 mm were classified as transport roots. The measure may not be completely accurate but could substantially exclude the partly decomposed roots which did not originate from absorptive roots or transport roots. These partly decomposed fine roots did not greatly impact the results as they accounted for less than 20% of the total dead roots. All fine root materials were dried at 50°C to a constant weight and weighed. The standing biomass of ARs and TRs was calculated based on live AR/TR ratio and standing fine root biomass, while the standing necromass of ARs and TRs was calculated based on dead AR/TR ratio and standing fine root necromass. The unit of fine root biomass in the soil cores was calculated as g m−2 at 0–20 cm depth.
Decomposition Experiment
The root mass loss pattern was assessed using litterbag method. To provide input parameters for the dynamic-flow model, we used four types of fine roots including live and dead absorptive and transport roots as decomposing materials to do in situ decomposition experiments. Approximately 0.15 g root material was put into each 15 cm × 3.5 cm litterbag (0.05 mm mesh). Each litterbag was filled with in situ collected organic matter and then inserted vertically into the 0–15 cm of soil. We conducted two root decomposition experiments to better quantify fine root mass loss dynamics during the different seasons. One began on March 31, 2016 (DE 1), and a second began on October 15, 2016 (DE 2) (Figure 1). After installation, two bags of each of four root types (60 bags per type) were retrieved from each plot on each sampling occasion. In DE 1, litterbags were collected after 50, 135 and 300 days of incubation. In DE 2, litterbags were collected after 90, 220 and 365 days of incubation. Roots from the bags were rinsed with clear tap water, carefully sorted, dried at 50°C to a constant weight and then weighed.
Estimation of Production, Mortality and Decomposition
Production, mortality and decomposition of the absorptive and transport roots were determined using a dynamic-flow model we developed (Li and Lange 2015).
GI-i and GII-i are the fine roots that died before the start of interval i and in interval i (1 ≤ i), respectively. The mass loss dynamics of GI-i and GII-i were determined by the litterbag method with dead and live roots used as decomposing materials, respectively.
Root mass loss pattern was simulated using the double exponential equation:
where y(t) and y0 are root mass at time t and the start, respectively. The two parameters λ and k were calculated based on the fine root mass remaining in litterbags collected on all sampling occasions using nonlinear regression.
The fine root mortality rate in interval i (μi) was assumed to be constant. The production (gi) and mortality (mi) in interval i were calculated by the following equations,
where Bi(0) and Bi represent the fine root biomass in soil cores sampled at the start and the end of interval i, respectively. Ni(0) and Ni represent the fine root necromass in soil cores collected at the start and the end of interval i. NII-i and NI-i are the mass remaining of GII-i and GI at the end of interval i, respectively. T was time length of interval i
where
was an exponential integral function (Abramowitz and Stegun 1964, ch. 6).
Bi, Bi(0), Ni, Ni(0), NII-i and NI-i had the unit g m−2. λI-i, kI-i, λII-i and kII-i were decomposition parameters for GI-i and GII-i, respectively, which were calculated as mentioned above. Bi, Bi(0), Ni, and Ni(0) were measured in the soil cores, NII-i was calculated by Eq. (5) and mi(t) = μi T. Thus, gi and di were calculated by Eqs. (2) and (3).
Modeling Decomposition Dynamics
We used data in DE 2 to infer the root decomposition dynamics from November 2016 to January 2017 and December 2017 to February 2018 as these two intervals occurred in the non-growing season, whereas the decomposition dynamics in other intervals were inferred using data in DE1 as these intervals occurred in the growing season.
Root decomposition dynamics in each interval were simulated using the procedures below. First, we developed double exponential decay models (Eq. 1) based on the percent mass remaining data in DE 1 and DE 2. Second, we calculated the percent mass remaining at different decomposition time points using the decay models previously developed. Third, we transformed the estimated percent remaining data with a natural function and used the cumulative soil temperature (10-day average) at the depth of 5 cm (Ts) and cumulative WTD (10-day average) to predict the natural log-transformed percent mass remaining. Lastly, we simulated root mass loss dynamics in each interval by inputting the cumulative Ts and cumulative WTD for that interval into the regression models.
Data Analysis
Effects of microtopography and soil depth on biomass allocation factors in each sampling occasion were assessed using a two-way factorial ANOVA. Simple repeated ANOVAs were used to test whether root biomass allocation factors change significantly over time, with sampling months as a within-subject variable. A post hoc test was conducted to assess which sampling months had significantly different biomass allocation factors. Two-way factorial ANOVAs were also used to test the effects of microtopography and soil depth on biomass of first-order, second-order, absorptive and transport roots, and necromass of absorptive and transport roots. A two-way ANOVA was used to test significant difference in absorptive and transport root production, mortality and decomposition estimates between hummocks and hollows. Raw data were natural log-transformed when necessary to establish homogeneity of variance. Significant differences between means were compared using Turkey’s significant test at the 0.05 level. All data were analyzed using SPSS (IBM SPSS Statistics 19.0) and MATLAB software (version 7.8; MathWorks; Natick, Massachusetts, US).
Results
Biomass Allocation
R1st/R2nd ratios did not change significantly over time, while AR/TR ratios showed significant seasonal change (Table 1). A post hoc test showed that AR/TR ratio was significantly higher in May, June, and September (growing season) compared to January and February (non-growing season) (Figure 2). Microtopography and soil depth significantly affected R1st/R2nd ratio and AR/TR ratio. Mean R1st/R2nd ratio and mean AR/TR ratio were significantly higher in hummocks than in hollows and significantly higher in the peat layer than in the muck and mineral layer (Figure 2).

Temporal changes in fine root biomass allocation factors in peat organic layer and muck and mineral layer at hummocks and hollows (n = 6, 1 ± SE). Different capital letters among columns within the same month stand for significant difference, while different bold italicized letters above column groups stand for significant difference among months (P < 0.05); H-OP and L-OP stand for peat organic payer in hummocks and hollows, respectively, while H-MM and L-MM stand for muck and mineral layer in hummocks and hollows, respectively.
Biomass and Necromass
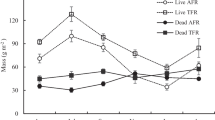
Total root mass at the peat layer and the muck and mineral layer was 965 ± 25 g m−2 and 308 ± 24 g m−2, respectively, in hummocks and 396 ± 16 g m−2 and 198 ± 15 g m−2, respectively, in hollows. After making area correction, total fine root mass was 995 ± 80 g m−2, with 76% in hummocks and 24% in hollows. The peat layer and the muck and mineral layer stored 74% and 26% of the total mass, respectively. Fine root biomass and necromass accounted for 43% and 57% of the total fine root mass, respectively. Both microtopography and soil depth significantly affected biomass and necromass of absorptive and transport roots (Table 2; Figure 3). The biomass and necromass of both types of roots were significantly higher in the hummocks than in the hollows, and significantly higher in the peat layer than in the muck and mineral layer (Table 2; Figure 3). Fine root mass showed highly seasonal variation. Absorptive and transport root biomass reached the highest values during growing season and the lowest values during dormant season, whereas absorptive and transport root necromass were higher in the dormant season than in the growing season (Figure 3). Microtopography and soil depth did not significantly affect the coefficients of variation (CV) of both types of roots (Table 3). The CVs of the live absorptive roots were higher than those of the dead absorptive roots at all positions (Table 3).

Temporal changes in biomass and necromass of absorptive and transport roots in peat layer (10 cm depth) in hummocks (H-OP) and hollows (L-OP) and muck and mineral layer (10 cm depth) in hummocks (H-MM) and hollows (L-MM) (1 SE, n = 5) from February 2016 to February 2018.
Mass Loss Dynamics
Both absorptive and transport roots decomposed significantly faster in the hummocks than in the hollows, but no significant difference in mass loss rate between absorptive and transport roots was found (Figure 4) in both growing and non-growing seasons. The double decay exponential equation fitted the observed percent root mass remaining data well in both the hummocks and the hollows (Table 4; Figure 4).

Mass loss patterns of absorptive (AR) and transport (TR) fine roots at hummocks and hollows in the experiments starting at the growing and the non-growing seasons (1SE, n = 5). H-AR and L-AR stand for absorptive root tissues buried in hummocks and hollows, respectively, while H-TR and L-TR stand for transport root tissues buried in hummocks and hollows, respectively. Different letters stand for significant difference in means. The coarse and thin short dash lines represent the simulated percent mass remaining of absorptive and transport roots in hummocks, respectively, while the coarse and thin long dash lines represent the simulated percent mass remaining of absorptive and transport roots in hollows, respectively.
Production, Mortality and Decomposition
The production, mortality and decomposition of absorptive and transport roots was significantly higher in the hummocks than in the hollows (Table 5). Absorptive roots had higher production, mortality and decomposition estimates than transport roots in the hummocks but lower estimates than transport roots in the hollows (Table 5). After making an area correction, mean annual fine root production, mortality and decomposition were 455 ± 106 g m−2, 475 ± 79 g m−2 and 392 ± 60 g m−2, respectively. The production, mortality and decomposition in the hummocks accounted for 77%, 76% and 75% of the total, respectively.
Discussion
Root Biomass Allocation Factors
R1st/R2nd ratio did not show evident temporal changes in any microtopographic location, indicating that first- and second-order roots have a similar biomass response to seasonal variability in Ts and WTD. By contrast, AR/TR ratios were significantly higher in the growing season compared to the non-growing season, and were positively related to seasonal change in Ts, showing that absorptive roots have a different response to temporal changes in Ts and WTD from third- and higher-order fine roots. This justifies our functional classification of treating first- and second-order roots as the ARs and third- and higher-order roots as TRs. The significant seasonal change in AR/TR ratios may be explained by eco-physiological traits between absorptive and transport roots. Absorptive roots have a faster growth rate and a shorter life span than transport roots and are more sensitive to seasonal changes in Ts (McCormack and Guo 2014; McCormack and others 2015). The faster growth rate of absorptive roots compared to transport roots during the growing season increased the relative proportion of live absorptive root biomass in live fine root pool, while the higher mortality of absorptive roots in the non-growing season decreased the relative live absorptive root proportion. As a result, the AR/TR ratio was higher in the growing season than in the non-growing season.
The R1st/R2nd ratio and the AR/TR ratio were significantly lower in the hollows compared to the hummocks and significantly lower in the muck and mineral layer compared to the peat layer. Shifts in biomass distribution among different root orders or functional groups in response to soil depth and microtopography may be due to several factors. First, trees may reduce photosynthate allocation to resource-acquiring roots in adaption to the poorly drained, waterlogged soil conditions and/or lower soil fertility (Day and Megonigal 1993; Makita and others 2010). Second, some tree species with higher R1st/R2nd ratio and AR/TR ratio may restrict roots to more aerobic and organic-rich layers. Third, longer waterlog and/or lower soil fertility can lead to morphological changes in fine roots (Day and Megonigal 1993; Krauss and others 2006), which may significantly increase the mass density of higher-order roots.
Root Biomass and Necromass
Microtopography significantly affected the biomass and necromass of absorptive and transport roots, causing higher biomass and necromass in the hummocks than in the hollows. Many studies also found higher fine root biomass and necromass in the hummocks compared to the hollows (Powell and Day 1991; Jones and others 1996; Baker and others 2001; Clawson and others 2001). However, Jones and others (1996, 2000) reported that fine root biomass values were comparable between the hummocks and the hollows in a FFW located in Alabama, the USA. Such an inconsistency may be ascribed to differences in fine root diameter limits among studies. Fine root diameter limit adopted in Jones and others (1996, 2000) was 5 mm rather than 2 mm. This may have confounded the microtopographic effects on fine root biomass because the transport roots accounted for a higher proportion in hollows than in hummocks. Setting such a larger diameter limit increased the proportion of transport roots in the total and therefore reduced the microtopographic effect. In this study, over 70% of absorptive and transport root biomass and necromass was distributed in the peat layer. Similar result was also found in a forested floodplain in which 74% of fine roots (≤ 3 mm) were restricted to the top 15 cm of soil (Baker and others 2001). Additionally, we found that the difference between the peat layer and the muck and mineral layer in root biomass was greater in hummocks compared to hollows probably due to smaller variability in soil water content between the two layers in the hollows as they were both under water for most of the year.
Jones and others (2000) found that fine root biomass was more temporally variable in hollows compared to hummocks in a FFW in floodplain of low order stream, while a study conducted in another floodplain site showed the opposite (Clawson and others 2001). Different from these reports, our study showed that neither absorptive roots nor transport roots had greater temporal variability in the hummocks than in the hollows, as indicated by comparable CV values. We found that CVs of absorptive root biomass were on average 10% higher than those of absorptive root necromass. The small seasonal variability in absorptive root necromass may be driven by slow decomposition rates of this flooded ecosystem, which results in greater dead absorptive root accumulation as expressed by little seasonal variation in absorptive root necromass.
Fine Root Mass Loss Rate
The mass loss rates of absorptive and transport roots at both microsites were within the ranges of fine root mass loss rates in FFWs reported by Stagg and others (2017) but lower than those in Crawford and others (2007). A significant microtopographic effect on the mass loss rates of both absorptive and transport roots was found in this study. However, seasonal changes in Ts and WTD did not confound the microtopographic effect as both absorptive and transport roots decomposed significantly faster in the hummocks than in the hollows during all seasons. The consistent microtopographic effect on fine root mass loss rates was in agreement with soil and coarse woody debris respiration in the same site, which showed significantly higher respiration rates in hummocks compared to hollows (Miao 2013, Miao and others 2017). The aerobic and organic-rich conditions in hummocks increase microbial activity (Minick and others 2019a, b). This together with greater fine root-derived organic matter inputs results in a higher organic matter turnover rate. We therefore concluded that hummocks are biogeochemical hot spots of soil C cycling in FFWs. Absorptive roots had slightly faster mass loss rates than transport roots in both the hummocks and the hollows. However, previous studies conducted in upland forests reported the opposite (Fan and Guo 2010; Xiong and others 2013; Kou and others 2018). This apparent contradiction may be due to the unique hydrologic conditions in FFWs. The frequent flooding in FFWs reduces soil microbial activity (Miao and others 2017; Minick and others 2019a) but may magnify leaching. Absorptive roots with a higher-surface-area-to-volume ratio would be more susceptible to leaching than transport roots (Beidler and Pritchard 2017), leading to a greater mass loss rate in absorptive roots.
Production, Mortality and Decomposition
Reported diameter-based fine root production estimates varied greatly among FFWs, ranging from 886 to 56 g m−2 y−1 due to the differences in study sites, fine root diameter limits and estimation methods (Symbula and Day 1988; Powell and Day 1991; Baker and others 2001; Clawson and others 2001; Pierfelice and others 2015). Most of these studies reported a higher fine root production in hummocks compared to hollows. Fine root production in this study was in the middle of this range, and fine root production, mortality and decomposition were significantly higher in hummocks compared to hollows. More importantly, the root function-based study could provide a more accurate fine root-derived C budget and a more comprehensive understanding of fine root dynamics including root responses to a changing environment. Our results showed that absorptive roots made greater contributions to the total production, mortality and decomposition than did transport roots at the ecosystem scale. However, the microtopography significantly affected the relative contributions of absorptive and transport roots to the total. The production, mortality and decomposition of absorptive roots were higher than those of transport roots in hummocks but lower than those of transport roots in hollows. This clearly demonstrated that root C cycling in hummocks and hollows was predominantly regulated by different types of fine roots, with absorptive roots in hummocks and transport roots in hollows. Compared with transport roots, absorptive roots were more responsive to microtopographic effect as indicated by the fact that the standing biomass in the hummocks were 1.6 and 1.8 times greater than those in the hollows for absorptive roots but 1.0 and 0.7 times greater than those in the hollows for transport roots. The mean decomposition estimates were lower than the mortality estimates, which means a net C input through fine roots. After making an area correction, net fine root mass sequestration was 92.6 g m−2 y−1, representing 22% of total C sequestration in a FFW (Krauss and others 2018). C sequestration through fine root dynamics mainly occurred in hummocks as fine roots made 82% of the contribution to the total root mass sequestration in the soil.
Implications
This two-year consecutive soil coring study has revealed that microtopography significantly affected fine root dynamics, with a greater biomass, production, mortality and decomposition occurring in hummocks compared to hollows. As the US Atlantic Coast is projected to have some of the highest rates of sea level rise globally (Sallenger and others 2012; IPCC 2013), the relative area of hummocks to hollows would decrease due to rising water table (Day and Templet 1989; Rotzoll and Fletcher 2013). This has potential to alter ecosystem level fine root dynamics and soil C sequestration by changing the relative rates of fine root production, mortality and decomposition in hummocks vs. hollows in these FFWs. In current study, we found AR/TR ratio and fine root biomass varied greatly over time and space due to highly temporal variability in soil temperature and hydrology. To better understand fine root dynamics, reliable and efficient methods are needed. One potential method to monitor fine root growth and death dynamics in wetlands is minirhizotrons (Iverson and others 2012). Unfortunately, this method has been found problematic in FFWs (unpublished research work; Baker and others 2001; Rodgers and others 2004). There are two reasons for this shortfall. First, frequently fluctuating water table in FFWS shifts soil materials around the minirhizotron tubes and obscures images. Second, some fine roots living in the peat layer in the hollows cannot be tracked over time as they drift with changing water table. In contrast, the dynamic-flow model we used here also had some difficulties with the confounding of spatial–temporal variation, but it is the only method that can give accurate estimates of fine root C dynamics in FFWs based on careful analysis of fine root functional morphology. Increasing sample size and sampling frequency can help to reduce uncertainties. Our data clearly show that absorptive roots are more responsive to the microtopographic effect than are transport roots, and therefore, future studies must carefully characterize both the relative distributions of hummocks vs. hollows and the very fine-scale distributions of functionally distinct absorptive and transport fine roots. Thus, models that hope to project future changes in belowground C cycling in coastal FFWs must therefore account for both absorptive and transport root dynamics, the demographic parameters of which vary continuously as a function of microtopography and hydrology.
References
Abramowitz M, Stegun I. 1964. Pocketbook of Mathematical Functions. Abridged edn. USA: National Bureau of Standards.
Baker TT, Conner WH, Lockaby BG, Stanturf JA, Burke MK. 2001. Fine root productivity and dynamics on a forested floodplain in South Carolina. Soil Sci Soc Am J 65:545–56.
Belyea LR, Clymo RS. 2001. Feedback control of the rate of peat formation. Proc R Soc London B 268:1315–21.
Beidler KV, Pritchard SG. 2017. Maintaining connectivity: understanding the role of root order and mycelial networks in fine root decomposition of woody plants. Plant Soil 420:19–36.
Bridgham SD, Megonigal JP, Keller JK, Bliss NB, Trettin C. 2006. The carbon balance of north American wetlands. Wetlands 26:889–916.
Clawson RG, Lockaby BG, Rummer B. 2001. Changes in production and nutrient cycling across a wetness gradient within a floodplain forest. Ecosystems 4:126–38.
Crawford ER, Day FP, Atkinson RB. 2007. Influence of environment and substrate quality on root decomposition in naturally regenerating and restored Atlantic white cedar wetlands. Wetlands 27:1–11.
Day FP, Megonigal JP. 1993. The relationship between variable hydroperiod, production allocation, and belowground organic matter turnover in forested wetlands. Wetlands 13:115–21.
Day JJ, Templet P. 1989. Consequences of sea level rise: implications from the Mississippi Delta. Coast Manage 17:241–57.
Duberstein JA, Conner WH, Krauss KW. 2014. Woody vegetation communities of tidal freshwater swamps in South Carolina, Georgia and Florida (US) with comparisons to similar systems in the US and South America. J Veg Sci 25:848–62.
Ensign SH, Noe GB. 2018. Tidal extension and sea-level rise: recommendations for a research agenda. Front Ecol Environ 16:37–43.
Fan PP, Guo DL. 2010. Slow decomposition of lower order roots: a key mechanism of root carbon and nutrient retention in the soil. Oecologia 163:509–15.
Guo D, Mitchell RJ, Withington JM, Fan PP, Hendricks JJ. 2008. Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: root branch order predominates. J Ecol 96:737–45.
Hertel D, Leuschner C. 2002. A comparison of four different fine root production estimates with ecosystem carbon balance data in a Fagus-Quercus mixed forest. Plant Soil 239:237–51.
IPCC. 2013. Climate change 2013: the physical science basis. In: Stocker D, Qin G-K, Plattner M, Tignor SK, Allen J, Boschung A, Nauels Y, Xia V Bex, Midgley PM, Eds. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press.
Iversen CM, Murphy MT, Allen MF, Childs J, Eissenstat DM, Lilleskov EA, Sarjala TM, Sloan VL, Sullivan PF. 2012. Advancing the use of minirhizotrons in wetlands. Plant Soil 352:23–39.
Jackson RB, Mooney HA, Schulze ED. 1997. A global budget for fine root biomass, surface area, and nutrient contents. Proc Natl Acad Sci USA 94:7362–6.
Jones RH, Lockaby BG, Somers GL. 1996. Effects of microtopography and disturbance on fine-root dynamics in wetland forests of low-order stream floodplains. Am Midl Nat 136:57–71.
Jones RH, Henson KO, Somers GL. 2000. Spatial, seasonal, and annual variation of fine root mass in a forested wetland. J Torrey Bot Soc 127:107–14.
Kou L, Jiang L, Fu X, Dai X, Wang H, Li S. 2018. Nitrogen deposition increases root production and turnover but slows root decomposition in Pinus elliottii plantations. New Phytol 218:1450–61.
Kou L, Guo DL, Yang H, Gao WL, Li SG. 2015. Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China. Plant Soil 391:207–18.
Krauss KW, Noe GB, Duberstein JA, Conner WH, Stagg CL, Cormier N, Jones MC, Bernhardt CE, Lockaby BG, From AS, Doyle TW, Day RH, Ensign Pierfelice K N, Hupp CR, Chow AT, Whitbeck JL. 2018. The role of the upper tidal estuary in wetland blue carbon storage and flux. Global Biogeochem Cycles 32:817–39.
Krauss KW, Whitbeck JL, Howard RJ. 2012. On the relative roles of hydrology, salinity, temperature, and root productivity in controlling soil respiration from coastal swamps (freshwater). Plant Soil 358:265–74.
Krauss KW, Doyle TW, Twilley RR, Rivera-Monroy VH, Sullivan JK. 2006. Evaluating the relative contributions of hydroperiod and soil fertility on growth of south Florida mangroves. Hydrobiologia 569:311–24.
Li X, Lange H. 2015. A modified soil coring method for measuring fine root production, mortality and decomposition in forests. Soil Biol Biochem 91:192–9.
Makita N, Hirano Y, Mizoguchi T, Kominami Y, Dannoura M, Ishii H, Finer L, Kanazawa Y. 2010. Very fine roots respond to soil depth: biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecol Res 26:95–104.
McCormack ML, Adams TS, Smithwick EAH, Eissenstat DM. 2014. Variability in root production, phenology, and turnover rate among 12 temperate tree species. Ecology 95:2224–35.
McCormack LM, Dickie IA, Eissenstat DM, Fahey TJ, Fernandez CW, Guo D, Helmisaari H-S, Hobbie EA, Iversen CM, Jackson RB, Leppälammi-Kujansuu J, Norby RJ, Phillips RP, Pregitzer KS, Pritchard SG, Rewald B, Zadworny M. 2015. Redefining fine roots improves understanding of belowground contributions to terrestrial biosphere processes. New Phytol 207:505–18.
McCormack LM, Guo D. 2014. Impacts of environmental factors on fine root lifespan. Front Plant Sci 5:205.
Megonigal JP, Day FP. 1992. Effects of flooding on root and shoot production of bald 5 cypress in large experimental enclosures. Ecology 73(4):1182–93.
Miao G. 2013. A multi-scale study on respiratory processes in a lower coastal plain forested wetland in the southeastern United States. Dissertation, North Carolina State University.
Miao G, Noormets A, Domec J-C, Fuentes M, Trettin CC, Sun G, McNulty SG, King JS. 2017. Hydrology and microtopography control carbon dynamics in wetlands: implications in partitioning ecosystem respiration in a coastal plain forested wetland. Agric For Meteorol 247:343–55.
Mitsch WJ, Gosselink JG. 1993. Wetlands. 2nd edn. New York: Van Nostrand Reinhold.
Mitsch WJ, Bernal B, Nahlik AM, Mander Ü, Zhang L, Anderson CJ, Jørgensen SE, Brix H. 2013. Wetlands, carbon, and climate change. Landsc Ecol 28:583–97.
Minick KJ, Kelley AM, Miao G, Li X, Noormets A, Mitra B, King JS. 2019a. Microtopography alters hydrology, phenol oxidase activity and nutrient availability in organic soils of a coastal freshwater forested wetland. Wetlands 39:263–73.
Minick JK, Mitra B, Li X, Noormets A, King J. 2019b. Water table drawdown alters soil and microbial carbon pool size and isotope composition in coastal freshwater forested wetlands. Front Glob Change . https://doi.org/10.3389/ffgc.2019.00007.
Moorhead KK, Brinson MM. 1995. Response of wetlands to rising sea level in the lower coastal plain of North Carolina. Ecol Appl 5:261–71.
Noe G, Hupp C, Rybicki N. 2013. Hydrogeomorphology influences soil nitrogen and phosphorus mineralization in floodplain wetlands. Ecosystems 16:75–94.
Norby RJ, Jackson RB. 2000. Root dynamics and global change: seeking an ecosystem perspective. New Phytol 147:3–12.
Ostertag R, Hobbie SE. 1999. Early stages of root and leaf decomposition in Hawaiian forests: effects of nutrient availability. Oecologia 121:564–73.
Persson H. 1980. Spatial distribution of fine-root growth, mortality and decomposition in a young scots pine stand in Central Sweden. Oikos 34:77–87.
Prieto I, Roumet C, Cardinael R, Dupraz C, Jourdan C, Kim JH, Maeght JL, Mao Z, Pierret A, Portillo N et al. 2015. Root functional parameters along a land-use gradient: evidence of a community-level economics spectrum. J Ecol 103:361–73.
Pierfelice K, Graeme LB, Krauss K, Conner W, Noe G, Ricker M. 2015. Salinity influences on aboveground and belowground net primary productivity in tidal wetlands. J Hydrol Eng 22:D5015002.
Poorter H, Nagel O. 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Aust J Plant Physiol 27:1191.
Powell SW, Day FP. 1991. Root production in four communities in the great dismal swamp. Am J Bot 78:288–97.
Pregitzer KS, DeForest JL, Burton AJ, Allen MF, Ruess RW, Hendrick RL. 2002. Fine root architecture of nine north American trees. Ecol Monogr 72:293–309.
Riggs SR. 1996. Sediment evolution and habitat function of organic-rich muds within the Albemarle estuarine system, North Carolina. Estuaries 19:169–85.
Rodgers HLR, Day FP, Atkinson R. 2004. Root dynamics in restored and naturally regenerated Atlantic white cedar wetlands. Restor Ecol 16:401–11.
Rotzoll K, Fletcher CH. 2013. Assessment of groundwater inundation as a consequence of sea-level rise. Nat Clim Change 3:477–81.
Sallenger AH Jr, Doran KS, Howd PA. 2012. Hotspot of accelerated sea-level rise on the Atlantic coast of North America. Nat Clim Change 2:884–8.
Schilling EB, Lockaby BG. 2005. Microsite influences on productivity and nutrient circulation within two southeastern floodplain forests. Soil Sci Soc Am J 69:1185–95.
Stagg CL, Schoolmaster DR, Krauss KW, Cormier N, Conner WH. 2017. Causal mechanisms of soil organic matter decomposition: deconstructing salinity and flooding impacts in coastal wetlands. Ecology 98:2003–18.
Symbula M, Day FP. 1988. Evaluation of two methods for estimating belowground production in a freshwater swamp forest. Am Midl Nat 120:405–15.
Sun JJ, Gu JC, Wang ZQ. 2012. Discrepancy in fine root turnover estimates between diameter-based and branch-order-based approaches: a case study in two temperate tree species. J For Res 23:575–81.
Xiong YM, Fan PP, Fu SL, Zeng H, Guo DL. 2013. Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees. Plant Soil 363:19–31.
Acknowledgement
Primary support was provided by USDA NIFA (Multi-agency A.5 Carbon Cycle Science Program) award 2014-67003-22068. Additional support was provided by DOE NICCR award 08-SC-NICCR-1072, the USDA Forest Service Eastern Forest Environmental Threat Assessment Center award 13-JV-11330110-081, and Carolinas Integrated Sciences and Assessments award 2013-0190/13-2322. The USFWS Alligator River National Wildlife Refuge provided helpful scientific discussions, the forested wetland research sites, and valuable in-kind support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author’s Contribution
X.L. and J. K. conceived the ideas and designed the research, X.L. analyzed the data; X.L., J.L. and K.M. collected the data; X.L., S.N., K.M., A.N., G.M., B.M., J.C., G.S. and J.K. led the writing of the manuscript.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Li, X., Minick, K.J., Luff, J. et al. Effects of Microtopography on Absorptive and Transport Fine Root Biomass, Necromass, Production, Mortality and Decomposition in a Coastal Freshwater Forested Wetland, Southeastern USA. Ecosystems 23, 1294–1308 (2020). https://doi.org/10.1007/s10021-019-00470-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-019-00470-x