
Abstract
Variation in aboveground net primary production (ANPP) is usually studied across wide environmental gradients focusing on spatial averages of zonal natural communities. We studied the spatial and temporal variation of ANPP of upland sown pastures and lowland natural grasslands across a narrow gradient of precipitation and temperature. The Flooding Pampa (Argentina) encompasses an 850–1000 mm range of mean annual precipitation and a 13.8–16.0°C range of mean annual temperature. For 15 100 × 100 km cells, we obtained mean monthly precipitation, temperature, and paddock-level ANPP of upland pastures and lowland grasslands during 8 years. Mean annual ANPP of lowland grasslands and upland sown pastures was positively related to mean annual precipitation. ANPP of upland pastures was 60–80% larger and increased more steeply with mean annual precipitation. ANPP seasonality also changed across the gradient. In lowland grasslands, as mean annual precipitation increased, ANPP monthly maximum increased, minimum decreased, and the duration of the growing season shortened. In contrast, in upland pastures, ANPP monthly maximum was constant, minimum increased, and the growing season lengthened with increasing precipitation. ANPP was more stable across years for lowland grasslands than for upland pastures. The response of annual ANPP to current-year precipitation decreased across the gradient, while the importance of the previous-year precipitation increased. In summary, we found strong spatial and temporal patterns of ANPP across a narrow environmental gradient. In addition, landscape position and species composition heavily influenced those patterns.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Aboveground net primary production (ANPP) integrates several aspects of ecosystem functioning (McNaughton and others 1989; Costanza and others 1997). In grazing systems, herbivore carrying capacity correlates with the spatial and temporal variation of ANPP (Oesterheld and others 1992; Oesterheld and others 1998; Irisarri and others 2013). As 80% of the energy required by livestock worldwide is provided by rangelands and sown pastures (Wheeler and others 1981; Oltjen and Beckett 1996), quantifying the patterns and environmental controls of ANPP is critical to understand and define grazing strategies. Previous studies have focused on natural vegetation gradients. However, approximately one-quarter of temperate and tropical grasslands worldwide have been converted to agriculture (Mock 2000). In those areas, sown pastures become key forage resources due to their positive effects on soil fertility, crop production (Franzluebbers and others 2011; Lemaire and others 2014), and economic stability (Pacín and Oesterheld 2014). Thus, it is important to evaluate the patterns and environmental controls of both natural grasslands and sown pastures.
Spatial and temporal correlation analyses have revealed the most important patterns and environmental controls of ANPP in natural grasslands. Mean annual precipitation and potential evapotranspiration usually explain a large fraction of mean annual ANPP across sites (Walter 1939; Rosenzweig 1968; Lauenroth 1979). Interestingly, most of these studies encompassed wide gradients from arid to wet extremes. Less is known about subregional gradients bounded to shorter precipitation ranges and finer spatial scales. The temporal (inter-annual) variability of annual ANPP in a given site also correlates with annual precipitation, although this association is weaker than for spatial gradients of mean annual precipitation (Le Houérou and others 1988; Lauenroth and Sala 1992). Vegetation constraints (associated with drought resistance syndromes of species) in drier sites, and nutrient and light constraints in wetter sites may account for this weakness (Paruelo and others 1999; Epstein and others 2006; La Pierre and others 2016). In addition, ANPP is correlated with the precipitation or ANPP of previous years (Lauenroth and Sala 1992; Jobbágy and Sala 2000; Oesterheld and others 2001; Fabricante and others 2009; Sala and others 2012). This “memory” is explained by soil water holding capacity, plastic responses of individual plants and population, or biogeochemical processes that affect nutrient availability (Schimel and Parton 1986; Wiegand and others 2004; Reichmann and others 2013).
Land-use change is also a major control of ANPP (Lauenroth and others 1999; Guerschman and others 2003; Bradford and others 2006; Baldi and Paruelo 2008, Di Bella and others 2009). However, the response of the inter-annual variability of ANPP to land-use change is less clear and contradictory in the literature (Lauenroth and others 2000; Verón and others 2002; Bradford et al. 2006; Grigera and others 2007a). Because land-use change is unevenly distributed across the landscape, crops and sown pastures often occupy the most favorable conditions, whereas natural grasslands remain restricted to soils with strong limitations for cropping (Burke and others 1994; Suttie and others 2012). As a result, contrasting forage resources become key landscape elements: high-input sown pastures are in the most favorable sites (Viglizzo and others 2011; Tribouillois and others 2015) and low-input natural grasslands in the least favorable sites (Baldi and others 2006).
Our objective was to reveal the spatio-temporal patterns and controls of the ANPP of upland sown pastures and lowland natural grasslands of the Flooding Pampa region. Natural grasslands and sown pastures in rotation with crops are distributed in a fine-grained pattern across the region. Natural grasslands occupy lowland areas with hydro-halomorphic soils (hereafter lowland grasslands), dominated by native and exotic grasses and forbs that progressively gain C4 grasses from SW to NE, following an increase of precipitation and temperature (León and others 1984; Sala and others 1986; Damario and Pascale 1988; Burkart and others 1990; Perelman and others 2001; Epstein and others 2002; Batista and others 2005). Sown pastures occupy upland areas with better drained soils (Hall and others 1992) in a 4 × 4 year no-till pasture-crop rotation. In contrast to lowland natural grasslands, they are often fertilized and weeded. They are typically composed by Festuca arundinacea, Dactylis glomerata and Lolium multiflorum grasses, and Medicago sativa, Trifolium pratense, and T. repens legumes (Grigera et al. 2007b).
We addressed four questions concerning the spatial and temporal patterns of ANPP:
- Question 1:
-
What is the relationship between mean annual ANPP and water availability for upland sown pastures and lowland natural grasslands?
- Question 2:
-
What is the relationship between ANPP seasonality and water availability, for upland sown pastures and lowland natural grasslands?
- Question 3:
-
What is the inter-annual variability of ANPP for upland sown pastures and lowland natural grasslands? Is it related to precipitation?
- Question 4:
-
What is the relationship between ANPP and current and previous-year precipitation across a regional gradient?
Methods
Data Source
The same data source was used to address the four questions. ANPP was derived from a monitoring system (http://www.agro.uba.ar/laboratorios/lart, Grigera et al. 2007b) which produces ANPP data at a monthly step. We analyzed data from eight growing seasons (July 2000–June 2001 through July 2007–June 2008). The number of paddocks monitored every year ranged between 840 and 1100 for upland pastures and between 950 and 1760 for lowland natural grasslands, and the total area was around 400,000 ha.
The ANPP monitoring system combines meteorological, satellite, and land-use data under the framework of the radiation use efficiency logic (Monteith 1972). ANPP results from the product of the incoming photosynthetically active radiation (PAR), the fraction absorbed by the canopy (fPAR), and the efficiency with which that energy was transformed into aboveground dry matter (that is, radiation use efficiency, RUE). PAR was obtained from two meteorological stations in the region. Because monthly PAR varied little among years (between 3 and 12% according to the month, https://eosweb.larc.nasa.gov/), average values were used. fPAR of each paddock was derived from satellite data: MODIS NDVI layer from the MOD13Q1 product (250 m and 16 days spatial and temporal resolutions, Grigera et al. 2007b; Caride and others 2012). Finally, RUE was estimated through a calibration that involved ANPP field estimations and remote sensing data (for more details see Grigera et al. 2007b).
Monthly precipitation and temperature between July 1999 and June 2008 were obtained from 42 meteorological stations located within 100 km of the study area (source: private farms and Ministry of Agriculture database www.minagri.gob.ar). For each meteorological station, we estimated potential evapotranspiration from temperature and latitude (Thornthwaite 1948), and water balance as the difference between precipitation and potential evapotranspiration. Annual precipitation, calculated as the sum of precipitation events across a growing season ranged from 850 mm.y−1 in the SW to 1000 mm.y−1in the NE. It was concentrated in spring and summer (70% of annual precipitation occurs between September and February). Mean annual precipitation correlated positively with mean minimum winter temperature (Jun.–Aug., range 3.6–4.5°C, R 2 = 0.58) and negatively with mean maximum summer temperature (Dec.–Feb., range 28.2–26.7°C, R 2 = 0.64).
Data Analysis
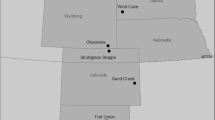
We used Quantum GIS (2.0.1-Dufour, http://qgis.osgeo.org) and the R (R Core Team 2013) software to process and analyze all the data. To merge ANPP derived from the monitoring system and meteorological data at a consistent spatial resolution, we divided the region into 15 100 × 100 km cells (Figure 1). Monthly ANPP of each paddock was averaged by landscape element (that is, upland sown pasture or lowland natural grassland) and cell. The minimum number of paddocks for a landscape element, cell, and month was 10 (mean 65 and 102 and maximum 330 and 350 for upland pastures and lowland grasslands, respectively). Monthly precipitation and water balance for each cell were interpolated from the 12 closer stations by the inverse of the squared distance to the cell centroid.

Study region with the 15 100 × 100 km cells overlapped. Paddocks with ANPP data are in black. The Flooding Pampa region is shown in dark gray. Other lines correspond to provincial borders. Bottom-right map location of the study region within Argentina (dark gray) and the rest of South America (light gray).
The first two questions were addressed by linear regression models between several descriptors of ANPP (dependent variables) and climatic variables (independent variables) across cells. The Durbin-Watson test (performed with the lmtest R package) showed no autocorrelation in linear models (1.5 < d < 2.5). For the first question, we regressed ANPP against precipitation and water balance. We considered both annual and quarter windows (spring: September–November, summer: December–February, autumn: March–May, and winter: June–August). However, we discarded annual-level water balance, because it was highly correlated with annual precipitation (R 2 = 0.97). Quarterly precipitation and water balance were considered only when they significantly outperformed mean annual precipitation, which is the explanatory variable most often found in the literature.
To address the second question, we regressed mean annual values of six variables that describe the seasonality of ANPP against mean annual precipitation, which, as shown before, covaried with temperature. The variables describing the seasonality of ANPP were mean maximum monthly ANPP (that is, the month of maximum ANPP of each growing season was averaged across growing seasons), mean minimum monthly ANPP, mean standard deviation of monthly ANPP within each growing season, and the mean start, end, and duration of the growing season. Although both lowland grasslands and upland pastures are active all year round (Semmartin and others 2007), about 70% of annual ANPP is concentrated in a comparatively short period. Thus, we defined each year’s growing season as the most productive months that concentrate at least 70% of annual ANPP. To obtain the months included in the growing season, we sorted the 12 months by decreasing ANPP. Then, we accumulated monthly ANPP from the most productive month until at least 70% of annual ANPP was reached. In average, these so defined growing seasons accumulated 73% (standard deviation 2% and maximum 80%) of annual ANPP. The earliest month (starting from July) was considered the growing season start, the latest was considered the growing season end, and the number of months was considered the duration of the growing season. Notice that occasionally the growing season duration was shorter than the difference between the last and first month in the list.
To address the third and fourth questions, we assessed the inter-annual variability of ANPP. For the third question, we compared the inter-annual coefficient of variation of annual ANPP of both landscape elements for each cell through Welch’s test, which is better suited than the t test to compare the means of two populations when they may have unequal variances (Welch 1947). For the fourth question, temporal multiple linear regression models between annual ANPP (dependent variable) and annual precipitation of the current and previous year (independent variables) for each cell of the grid were performed. Then, the relative weights of the regression parameters were compared through Welch’s test.
Results
Mean Annual ANPP and Water Availability
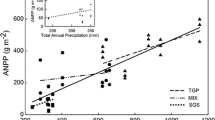
Mean annual ANPP of upland pastures and lowland grasslands increased linearly across the gradient, but the specific models for each landscape element differed (Figure 2). Mean annual precipitation explained a 12% higher proportion of mean annual ANPP variations in lowland grasslands than in upland pastures (Figure 2). In addition, ANPP of upland pastures was better explained by summer water balance than by mean annual precipitation (R 2 = 0.58 vs 0.46). The ANPP of upland pastures ran above the ANPP of lowland grasslands with a higher slope (Figure 2). As a consequence, precipitation-use-efficiency (the ratio between mean annual ANPP and precipitation) was 60% higher for upland pastures than for lowland grasslands at the driest end of the gradient and 80% higher at the wettest end.

Relationship between mean annual ANPP and mean annual precipitation for upland pastures and lowland grasslands. Solid lines represent the best fit models: upland pastures ANPP = −10883 + 18.2 mean annual precipitation (R 2 = 0.46); lowland grasslands ANPP = −3162 + 8.0 mean annual precipitation (R 2 = 0.70). Both models were statistically significant (p < 0.05).
ANPP Seasonality and Water Availability
Seasonality of upland pastures and lowland grasslands varied differently across the gradient (Figure 3). Mean maximum monthly ANPP (Figure 3a) increased with mean annual precipitation in lowland grasslands, whose species composition is enriched in C4 species at the wet end of the gradient, but did not change significantly in upland pastures, which are composed of C3 species across the gradient. As expected from these community patterns, mean minimum monthly ANPP (Figure 3b) decreased with mean annual precipitation in lowland grasslands and increased in upland pastures. Seasonality, measured as the intra-annual standard deviation of ANPP (Figure 3c), increased with mean annual precipitation in lowland grasslands but did not change significantly in upland pastures. Regarding the timing of the growing season, the start became earlier as mean annual precipitation increased in upland pastures and later for lowland grasslands (Figure 3d). The end of the growing season did not change across the gradient for upland pastures and was anticipated 1 month for lowland grasslands (Figure 3e). Consequently, as mean annual precipitation increased, the duration of the growing season was extended for upland pastures and shortened for lowland grasslands (Figure 3f).

Relationship between six variables describing ANPP seasonality and mean annual precipitation for upland pastures and lowland grasslands: maximum A and minimum B mean monthly ANPP, mean intra-annual ANPP standard deviation C, and mean start D, end E, and duration F of the growing season. Only statistically significant (p < 0.05) best fit lines are shown.
Inter-Annual Variability of ANPP
The inter-annual relative variability of ANPP (coefficient of variation, CV) was larger for the upland pastures than for the lowland grasslands in 13 of the 15 cells (mean values = 0.10 and 0.08, Figure 4a). The difference was significant even when the cell with extremely high inter-annual CV for upland pastures was excluded (p = 0.016). Given the higher ANPP of upland pastures compared to lowland grasslands, the slight difference in the inter-annual CV implies a proportionally higher difference in the standard deviation. Except for the upland pastures of one cell, the inter-annual CV of ANPP was lower than the inter-annual CV of precipitation, (Figure 4b). Moreover, there was no association between the inter-annual CV of mean annual precipitation and that of ANPP.

A Comparison of the inter-annual relative variability of ANPP between upland pastures and lowland grasslands. B Relationship of the inter-annual relative variability of ANPP and precipitation for lowland grasslands and upland pastures. The diagonal line represents the 1:1 relationship.
Annual ANPP vs Current and Previous-Year Precipitation
Inter-annual variability of ANPP was explained by linear models based on current and previous-year precipitation. The coefficient of determination (R 2) of the models varied across the region and for different landscape elements between 0.12 and 0.92 (mean = 0.60). The response of annual ANPP to current-year precipitation decreased across the regional gradient (Figure 5a). This decrease was significantly steeper for upland pastures than for lowland grasslands (marginal responses were −0.026 and −0.009, respectively). In contrast, the response of annual ANPP to previous-year precipitation did not vary across the precipitation gradient for lowland grasslands but increased for upland pastures (Figure 5b). Consequently, for both landscape units, drier sites were more responsive to current-year precipitation and wetter sites were more responsive to previous-year precipitation.

Effect of current-year A and previous-year B precipitation on ANPP across the regional precipitation gradient for lowland grasslands and upland pastures. Lines represent the statistically significant (p < 0.05) best fit models. Lowland grasslands current-year: Y = 9.66. −0.009 X (R 2 = 0.83), Upland pastures current-year: Y = 26.56. −0.026 X (R 2 = 0.50). Upland pastures previous year: Y = −11.88. 0.014 X (R 2 = 0.31).
Discussion
Mean Annual ANPP and Water Availability
The regional patterns of ANPP were strongly associated with precipitation and water balance, even in a narrow gradient of just 150 mm, and differed between the two landscape elements. Upland pastures presented higher ANPP and, as a consequence, higher precipitation-use-efficiency than lowland grasslands. Upland pastures occupy the best drained soils, which contrast with the hydro-halomorphic limitations of lowland grassland soils (León et al. 1984). They are composed of species selected for high growth rate and are subjected to more intense agricultural management (that is, seeding, fertilization, weed control) than lowland natural grasslands.
The relationship between ANPP and precipitation of both landscape elements differed with that observed for other grassland systems of the world (McNaughton 1985; Sala and others 1988; Bai and others 2008). These studies performed on different zonal natural grasslands presented a similar relationship between ANPP and precipitation (ANPP = −603 + 6.5 mean annual precipitation; standard deviation 365 and 0.5). For lowland grasslands, the lower increase of the precipitation-use-efficiency across the precipitation gradient resulted on an ANPP 1000 and 1500 kg.ha−1.y−1 lower than reported by previous studies. Likely, soil hydro-halomophic limitations account for this lower ANPP. In contrast, upland pastures presented a higher increase of precipitation-use-efficiency across the precipitation gradient compared to previous studies, which resulted in similar ANPP at the dry end of the gradient, and 1500 kg.ha−1.y−1 higher at the wet end. This steeper increase of ANPP for upland pastures may be accounted for by fertilization and the combination of C3 grasses and legume species, which may remove the increasing nitrogen limitation as mean annual precipitation increases (Epstein et al. 2006). Contrary to what we observed in upland pastures, Lauenroth et al. (2000) observed a decrease of precipitation-use-efficiency in cropped systems across a precipitation gradient. As they studied wheat systems, this opposing pattern could be explained by the differences in the period of occupation between pastures and wheat (perennial and seasonal, respectively).
ANPP Seasonality and Water Availability
The regional precipitation and temperature gradients also explained the variation in seasonality of both landscape elements. In lowland grasslands, the higher abundance of C4, warm-season species at the wet end of the gradient with less severe winters (Perelman et al. 2001; Epstein et al. 2002) is consistent with our findings of a later beginning of the growing season and higher seasonality of ANPP, concentrated during spring and summer. In contrast, the upland pastures, which are C3-dominated and with a constant floristic composition across the gradient is consistent with an early start of the growing season, an increase of winter ANPP and a longer growing season across the gradient, all likely influenced by temperature. These opposite responses of lowland grasslands and upland pastures across the gradient resulted in lowland grasslands having a 1-month longer growing season than upland pastures at the dry and cold end of the gradient and a 1-month shorter growing season at the wet and warm end. The importance of temperature as a driver of ANPP is observed in the higher goodness of fit of the spatial model explaining annual ANPP for upland pastures based on summer water balance compared to annual precipitation. This pattern also highlights the importance of precipitation and temperature seasonality as ANPP drivers (Guo and others 2012).
Inter-Annual Variability of ANPP
The inter-annual variability of ANPP was higher for upland pastures than for lowland grasslands. The different inter-annual variability of both landscape elements may result from two factors: the intensity of climatic events in each forage resource and the sensitivity of each landscape element to a given event. Due to the differences in soil properties and topographic position, precipitation fluctuations should result in more intense modifications in the lowland soils, because they are less permeable and more susceptible to flooding and drought than the upland soils, which are better drained and have higher water retention capacity (Bork and others 2001; Perelman and others 2001; Swemmer and others 2007). However, ANPP varied more in upland pastures than in lowland grasslands, likely as a result of the larger niche complementarity in the more diverse system (Tilman and others 1997), the lowland grasslands. For both landscape elements, the relative variation of ANPP was lower than that of precipitation, a pattern that contrasts with the relative variation of wheat yield in South America, which was higher than that of precipitation (Verón and others 2002). Similarly, to Oesterheld and others (1999)and opposite to other studies (Knapp and Smith 2001; Sanderson and others 2005), the decrease of ANPP during dry years was similar to the increase during wet years (data not shown).
Annual ANPP vs Current and Previous-Year Precipitation
Current and previous-year precipitation explained a large proportion of annual ANPP. The response of ANPP to current-year precipitation decreased across the gradient for lowland grasslands and upland pastures. As the direct response of ANPP to precipitation occurs in the short-term, the lower frequency of years where water is a limiting factor at the wet end of the gradient may explain the decreasing response of ANPP to current-year precipitation (Sala and others 2012; Petrie and others 2016). In contrast, the response of ANPP to previous-year precipitation increased across the gradient for upland pastures. Previous-year precipitation may affect nutrient availability in the subsequent year and hence ANPP (Yahdjian and others 2011). This response will be higher in wet habitats since nutrient limitation increases with humidity (Paruelo and others 1999; Epstein and others 2006). Species turnover and plant cover change, expected to explain differences between both landscape elements probably act at longer time scales not covered in our work (Reichmann and others 2013; Wilcox and others 2016).
The relationship between previous-year precipitation and ANPP provides evidence to generate scenarios of ANPP in the next growing season, although current-year precipitation will eventually define ANPP. According to the model obtained for upland pastures, an average increase (+1 standard deviation) of precipitation at the dry end of the gradient will increase annual ANPP by 550 kg.ha−1.y−1 for the current year and 90 kg.ha−1.y−1 for the following year. For the wet end of the gradient, the corresponding figures would be ANPP 150 and 380 kg.ha−1.y−1. For lowland grasslands, the corresponding variations of annual ANPP would be 240 and 90 kg.ha−1.y−1 for the dry end, and 10 and 100 kg.ha−1.y−1 for the wet end.
Conclusion
We described the patterns and controls of ANPP of two contrasting landscape elements across the same regional gradient. As a result, we observed that upland sown pastures, which occupied well-drained soils had a higher response to mean annual precipitation, lower stability, and was more sensitive to previous-year precipitation as mean annual precipitation increased than lowland natural grasslands. These findings also provide policy makers with a technological tool that could help them anticipate extreme weather conditions, something that will become more frequent under the ongoing climate change (Yahdjian and Sala 2008; Petrie and others 2016).
References
Bai Y, Wu J, Xing Q, Pan Q, Huang J, Yang D, Han X. 2008. Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau. Ecology 89:2140–53.
Baldi G, Guerschman JP, Paruelo JM. 2006. Characterizing fragmentation in temperate South America grasslands. Agric Ecosyst Environ 116:197–208.
Baldi G, Paruelo JM. 2008. Land-use and land cover dynamics in South American temperate grasslands. Ecol Soc 13:6.
Batista WB, Taboada MA, Lavado RS, Perelman SB, León RJC. 2005. Asociación entre comunidades vegetales y suelos en el pastizal de la Pampa Deprimida. In: Oesterheld M, Aguiar M, Ghersa C, Paruelo JM, Eds. La heterogeneidad de la vegetación de los agroecosistemas. Editorial Facultad de Agronomía. pp 113–29.
Bork EW, Thomas T, McDougall B. 2001. Herbage response to precipitation in Central Alberta boreal grasslands. J Range Manag 54:243–8.
Bradford JB, Lauenroth WK, Burke IC, Paruelo JM. 2006. The influence of climate, soils, weather, and land use on primary production and biomass seasonality in the US Great Plains. Ecosystems 9:934–50.
Burkart SE, León RJC, Movia CP. 1990. Inventario fitosociológico del pastizal de la Depresión del Salado (Prov. Bs. As.) en un área representativa de sus principales ambientes. Darwiniana 30:27–69.
Burke IC, Lauenroth WK, Parton WJ, Cole CV. 1994. Interactions of landuse and ecosystem function: a case study in the central Great Plains. In: Groffman PM, Likens GE, Eds. Integrated regional models: interactions between humans and their environment. New York: Chapman Hall. p 79–95.
Caride C, Piñeiro G, Paruelo JM. 2012. How does agricultural management modify ecosystem services in the argentine Pampas? The effects on soil C dynamics. Agric Ecosyst Environ 154:23–33.
Costanza R, d’Arge R, de Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, O’Neill RV, van den Belt M. 1997. The value of the world’s ecosystem services and natural capital. Nature 387:253–60.
Damario EA, Pascale AJ. 1988. Características agroclimáticas de la Región Pampeana Argentina. Rev Fac de Agron (UBA) 9:41–64.
Di Bella CM, Negri IJ, Posse G, Jaimes FR, Jobbágy EG, Garbulsky F, Deregibus VA. 2009. Forage production of the Argentine Pampa Region based on land use and long-term normalized difference vegetation index data. Rangel Ecol Manag 62:163–70.
Epstein HE, Gill RA, Paruelo JM, Lauenroth WK, Jia GJ, Burke IC. 2002. The relative abundance of three plant functional types in temperate grasslands and shrublands of North and South America: effects of projected climate change. J Biogeogr 29:875–88.
Epstein HE, Paruelo JM, Piñeiro G, Burke IC, Lauenroth WK, Barret JE. 2006. Interactions of water and nitrogen on primary productivity across spatial and temporal scales in grassland and shrubland ecosystems. In: D’Odorico P, Porporato A, Eds. Dryland ecohydrology. Netherland: Springer. p 201–11.
Fabricante I, Oesterheld M, Paruelo JM. 2009. Annual and seasonal variation of NDVI explained by current and previous precipitation across Northern Patagonia. J Arid Environ 73:745–53.
Franzluebbers AJ, Sulc RM, Russelle MP. 2011. Opportunities and challenge for integrating North-American crop and livestock systems. In: Lemaire G, Hodgson J, Chabbi A, Eds. Grassland productivity and ecosystem services. Wallingford: CAB Int. p 208–18.
Grigera G, Oesterheld MO, Durante M, Pacín F. 2007a. Evaluación y seguimiento de la productividad forrajera. Rev Argent de Prod Anim 27:137–48.
Grigera G, Oesterheld M, Pacín F. 2007b. Monitoring forage production for farmers’ decisión making. Agric Syst 94:637–48.
Guerschman JP, Paruelo JM, Burke IC. 2003. Land use impacts on the normalized difference vegetation index in temperate Argentina. Ecol Appl 13:616–28.
Guo Q, Hu Z, Li S, Li X, Sun X, Yu G. 2012. Spatial variations in aboveground net primary productivity along a climate gradient in Eurasian temperate grassland: effects of mean annual precipitation and its seasonal distribution. Glob Chang Biol 18:3624–31.
Hall AJ, Rebella CM, Ghersa CM, Culot JPH. 1992. Field-crop systems of the Pampas. In: Pearson CJ, Ed. Ecosystems of the Worlds. Field crop ecosystems. Amsterdam: Elsevier. p 413–50.
Irisarri JGN, Oesterheld MO, Paruelo JM, Oyarzábal M, Durante M. 2013. Monitoring the Ecosystem Service of Forage Production. In: Alcaraz-Segura D, DiBella CM, Straschnoy JV, Eds. Earth Observation of Ecosystem Services. Boca Raton: CRC Press—Taylor and Francis Group. p 87–103.
Jobbágy EG, Sala OE. 2000. Controls of grass and shrub aboveground production in the patagonian steppe. Ecol Appl 10:541–9.
Knapp AK, Smith MD. 2001. Variation among biomes in temporal dynamics of aboveground primary production. Science 291:481–4.
Lauenroth WK. 1979. Grasslands primary production. In: French NR, Ed. North American grasslands in perspective. New York: Springer. p 3–24.
Lauenroth WK, Sala OE. 1992. Long-term forage production of North American shortgrass steppe. Ecol Appl 2:397–403.
Lauenroth WK, Burke IC, Gutmann MP. 1999. The structure and function of ecosystems in the central North American grassland region. Gt Plains Res 9:223–59.
Lauenroth WK, Burke IC, Paruelo JM. 2000. Patterns of production and precipitation-use efficiency of winter wheat and native grasslands in the Central Great Plains of the United States. Ecosystems 3:344–51.
La Pierre KJ, Blumenthal DM, Brown CS, Klein JA, Smith MD. 2016. Drivers of variation in aboveground net primary productivity and plant community composition differ across a broad precipitation gradient. Ecosystems. doi:10.1007/s10021-015-9949-7.
Le Houérou HN, Bingham RL, Skerbek W. 1988. Relationship between the variability of primary production and the variability of annual precipitation in world arid lands. J Arid Environ 15:1–18.
Lemaire G, Franzluebbers A, Carvalho PC, de Faccio PC, Dedieu B. 2014. Integrated crop-livestock systems: strategies to achieve synergy between agricultural production and environmental quality. Agric Ecosyst Environ 190:4–8.
León RJC, Rusch GM, Oesterheld M. 1984. Pastizales pampeanos-impacto agropecuario. Phytocoenologia 12:201–18.
McNaughton SJ. 1985. Ecology of a grazing ecosystem: the Serengeti. Ecol Monogr 55:259–94.
McNaughton SJ, Oesterheld M, Frank DA, Williams KJ. 1989. Ecosystem-level patterns of primary productivity and herbivory in terrestrial habitats. Nature 341:142–4.
Mock G. 2000. Domesticating the world: conversion of natural ecosystems. WRI, UNEP, UNDP, World Bank (Eds.), World Resources 2000–2001, World Resources Institute, Washington. pp 24–25.
Monteith J. 1972. Solar radiation and productivity in tropical ecosystems. J Appl Ecol 9:747–66.
Oesterheld M, Sala OE, McNaughton SJ. 1992. Effect of animal husbandry on herbivore-carrying capacity at a regional scale. Nature 356:234–6.
Oesterheld M, DiBella CM, Kerdiles H. 1998. Relation between NOAA-AVHRR satellite data and stocking rate of rangelands. Ecol Appl 8:207–12.
Oesterheld M, Loreti J, Paruelo JM, Semmartin M, Walker L. 1999. Grazing, fire, and climate effects on primary productivity of grasslands and savannas. In: Walker LR, Ed. Ecosystems of disturbed ground. Amsterdam: Elsevier. p 287–306.
Oesterheld M, Loreti J, Semmartin M, Sala OE. 2001. Inter-annual variation in primary production of a semi-arid grassland related to previous-year production. J Veg Sci 12:137–42.
Oltjen JW, Beckett JL. 1996. Role of ruminant livestock in sustainable agricultural systems. J Anim Sci 74:1406–9.
Pacín F, Oesterheld M. 2014. In-farm diversity stabilizes return on capital in Argentine agro-ecosystems. Agric Syst 124:51–9.
Paruelo JM, Lauenroth WK, Burke IC, Sala OE. 1999. Grassland precipitation-use efficiency varies across a resource gradient. Ecosystems 2:64–8.
Perelman SB, León RJC, Oesterheld M. 2001. Cross-scale vegetation patterns of Flooding Pampa grasslands. J Ecol 89:562–77.
Petrie MD, Brunsell NA, Vargas R, Collins SL, Flanagan LB, Hanan NP, Litvak ME, Suyker AE. 2016. The sensitivity of carbon exchanges in Great Plains grasslands to precipitation variability. J Geophys Res Biogeosci. doi:10.1002/2015JG003205.
R Core Team. 2013. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Reichmann LG, Sala OE, Peters DPC. 2013. Precipitation legacies in desert grassland primary production occur through previous-year tiller density. Ecology 94:435–43.
Rosenzweig ML. 1968. Net Primary Productivity of Terrestrial Communities: prediction from Climatological Data. Am Nat 102:67–74.
Sala OE, Oesterheld MO, León RJC, Soriano A. 1986. Grazing effects upon plant community structure in subhumid grasslands of Argentina. Vegetatio 67:21–32.
Sala OE, Parton WJ, Joyce LA, Lauenroth WK. 1988. Primary production of the central grassland region of the United States. Ecology 69:40–5.
Sala OE, Gherardi LA, Reichmann L, Jobbágy EG, Peters DPC. 2012. Legacies of precipitation fluctuations on primary production: theory and data synthesis. Philos Trans R Soc B Biol Sci 367:3135–44.
Sanderson MA, Soder KJ, Muller LD, Klement KD, Skinner RH, Goslee SC. 2005. Forage mixture productivity and botanical composition in pastures grazed by dairy cattle. Agron J 97:1465.
Semmartin M, Oyarzábal M, Loreti J, Oesterheld M. 2007. Controls of primary productivity and nutrient cycling in a temperate grassland with year-round production. Austral Ecol 32:416–28.
Schimel DS, Parton WJ. 1986. Microclimatic controls of nitrogen mineralization and nitrification in shortgrass steppe soils. Plant Soil 93:347–57.
Suttie JM, Reynolds SG, Batello C. 2012. Grassland perspectives. In: Suttie JM, Reynolds SG, Batello C, Eds. Grasslands of the world. ROME: FAO. p 463–93.
Swemmer AM, Knapp AK, Snyman HA. 2007. Intra-seasonal precipitation patterns and above-ground productivity in three perennial grasslands. J Ecol 95:780–8.
Thornthwaite CW. 1948. An approach toward a rational classification of climate. Geogr Rev 38:55–94.
Tilman D, Lehman CL, Thomson KT. 1997. Plant diversity and ecosystem productivity: theoretical considerations. Proc Natl Acad Sci 94:1857–61.
Tribouillois H, Fort F, Cruz P, Charles R, Flores O, Garnier E, Justes E. 2015. A functional characterisation of a wide range of cover crop species: growth and nitrogen acquisition rates. Leaf traits and ecological strategies. PLoS One 10:3.
Verón SR, Paruelo JM, Sala OE, Lauenroth WK. 2002. Environmental controls of primary production in agricultural systems of the Argentine Pampas. Ecosystems 5:625–35.
Viglizzo EF, Frank FC, Carreño LV, Jobbágy EG, Pereyra H, Clatt J, Pincén D, Ricard MF. 2011. Ecological and environmental footprint of 50 years of agricultural expansion in Argentina. Glob Chang Biol 17:959–73.
Walter H. 1939. Grassland, savanne und busch der ariderenteileAfrikas in ihrerokologischenbedingtheit. Jahrb fur Wiss Bot 87:750–860.
Welch BL. 1947. The generalization of “Student’s” problem when several different population variances are involved. Biometrika 34:28–35.
Wheeler RD, Kramer GL, Young KB, Ospina E. 1981. The world Livestock product feedstuff, and food grain system. Morrilton: Winrock International. p 85.
Wiegand T, Snyman HA, Kellner K, Paruelo JM. 2004. Do grasslands have a memory: modeling phytomass production of a semiarid South African grassland. Ecosystems 7:243–58.
Wilcox KR, Blair JM, Smith MD, Knapp AK. 2016. Does ecosystem sensitivity to precipitation at the site-level conform to regional-scale predictions? Ecology 97:561–8. doi:10.1890/15-1437.1.
Yahdjian L, Sala OE. 2008. Climate change impacts on South American rangelands. Rangelands 30:34–9.
Yahdjian L, Gherardi L, Sala O. 2011. Nitrogen limitation in arid-subhumid ecosystems: a meta-analysis of fertilization studies. J Arid Environ 75:675–80.
Acknowlegments
We thank all CREA farmers for providing field data. Victor Suárez and LART personnel helped in the computer implementation and data processing. This research was funded by University of Buenos Aires, ANPCyT (PICT 2013-1785),CONICET (PIP 555), and IPCVA. Martín Durante was funded by a CONICET fellowship.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author contributions
M. Durante, M. Oesterheld, and G. Piñeiro conceived the study; M. Durante, M. Oesterheld, G. Piñeiro, and G. Irisarri performed the research; M. Durante analyzed the data; G. Irisarri contributed new models; and M. Durante and M. Oesterheld wrote the paper.
Rights and permissions
About this article

Cite this article
Durante, M., Piñeiro, G., Irisarri, J.G.N. et al. Primary Production of Lowland Natural Grasslands and Upland Sown Pastures Across a Narrow Climatic Gradient. Ecosystems 20, 543–552 (2017). https://doi.org/10.1007/s10021-016-0039-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-016-0039-2