
Abstract
A total of 26 proteolytic moderate halophiles were isolated and characterized. Most isolates were members of the genus Salinivibrio (16 strains), while others were identified as Bacillus (4 strains), Salinicoccus (2 strains), or members of the γ-Proteobacteria (4 strains). Strain CP76 was selected as the best producer of an extracellular protease, designated CP1, and was used for further studies. Sequence analysis of the 16S rRNA gene in addition to phenotypic tests led to the placement of this organism in the genus Pseudoalteromonas. Maximal protease production was detected at the end of the exponential growth phase. This CP1 protease was purified and biochemically characterized, showing optimal activity at 55 °C, pH 8.5, and high tolerance to a wide range of NaCl concentrations (0 to 4 M NaCl). The most interesting features of this enzyme are its moderate thermoactivity, its activity at a range of pH values (6–10), and, especially, its salt tolerance (optimal activity at 7.5% total salts). The purified protease has a molecular mass of 38 kDa, and the N-terminal amino acid sequence determined showed similarity to metalloproteases previously described. The protease activity was strongly inhibited by EDTA, PMSF, and Pefabloc. No significant inhibition was detected with E-64, bestatin, chymostatin, or leupeptin. These results suggest that Pseudoalteromonas sp. strain CP76 produces an extracellular metalloprotease moderately thermotolerant and stable at high salt concentrations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bacterial proteases are a well-studied group of hydrolases that catalyze the total hydrolysis of proteins. Besides their physiological importance, they constitute a class of enzymes of great application in commercial fields (Rao et al. 1998). While several proteases from mesophilic microorganisms have been purified and characterized, and the corresponding genes have been cloned (Rao et al. 1998), proteases from halophilic microorganisms have not been extensively studied. A few proteases from extreme halophiles, members of the archaeal phylogenetic branch, have been characterized (Norberg and Hofsten 1969; Kamekura and Seno 1990; Kamekura et al. 1992; Stepanov et al. 1992; Ryu et al. 1994; Giménez et al. 2000), but no proteases from moderately halophilic bacteria have been purified and studied in depth so far (Ventosa et al. 1998). Since moderately halophilic microorganisms are able to grow over a wide range of salt concentrations (optimal growth at 3–15% NaCl) (Ventosa et al. 1998), they constitute a very interesting group of organisms with great potential for use in biotechnology. The screening and isolation of moderate halophiles with protease activity will allow the characterization of the enzymes and the cloning of encoding genes. The possibility of using enzymes from halophilic bacteria in industrial processes has the advantage of optimal activities at high salt concentrations (Ventosa et al. 1998). The interest in these enzymes resides in both their biotechnological potential for novel applications and the need for a better understanding of their intrinsic salt-resistance properties and the stabilization mechanisms that enable extracellular enzymes of halophiles to have activity in the extreme environments in which these microorganisms are normal inhabitants.
In order to isolate moderately halophilic microorganisms able to produce proteases, we performed a screening. A total of 26 proteolytic, moderately halophilic bacteria were isolated from salterns and saline soils. Strain CP76 was selected as the best producer of an extracellular protease. The protease, designated protease CP1, was purified and biochemically characterized. The combination of salt and temperature tolerance made this enzyme a good candidate not only for industrial applications but also for elucidating the molecular basis of enzymes' osmostability from halophilic microorganisms. To our knowledge, this study constitutes the first report of the purification and characterization of a proteolytic enzyme from a moderately halophilic microorganism.
Materials and methods
Bacterial strains and culture conditions
Bacterial strains were routinely cultured on a saline medium (SW) with a final concentration of 10% (w/v) total salts (SW-10) supplemented with 0.5% (w/v) yeast extract (Ventosa et al. 1982). When necessary, different concentrations of salt mixture were added to the basal medium. Growth curves of strain CP76 were obtained by monitoring the culture absorbance at 600 nm by using a Perkin-Elmer spectrophotometer at different incubation times. For this purpose, 250-ml flasks containing 50 ml of SW medium were each inoculated with 500 μl of a stationary-phase culture and incubated at 37 °C and 250 rpm on a New Brunswick environmental incubator shaker.
The effects of culture conditions on protease production were assayed by growing the microorganism in SW-7.5 medium to which different carbohydrates such as lactose, sucrose, glycerol, maltose, mannitol, fructose, glucose (50 mM), NH4Cl (50 mM), or Casamino Acids (Difco) (1%) were added. Growth was monitored as previously described, and protease activity was determined after 24 h of incubation.
Screening of strains for proteolytic activity
A total of 310 moderately halophilic strains were isolated from water and sediment of solar salterns and hypersaline soils located in Isla Cristina (Huelva, Spain). Samples were collected in sterile plastic containers and were plated not later than 6 h after collection. Proteolytic activity of the cultures was screened qualitatively in a saline medium containing milk (50%) plus 10% (w/v) total salts (Ventosa et al. 1982) supplemented with 0.5% (w/v) yeast extract (Difco) and 1% peptone. The medium was solidified by adding 20 g/l Bacto-Agar (Difco). Clear zones around the colonies appearing over the next 48 h were taken as evidence of proteolytic activity.
Characterization of isolates
In order to characterize the strains taxonomically, several morphological, biochemical, physiological, and nutritional features were investigated. Gram staining, catalase, oxidase, growth under anaerobic conditions, and other biochemical tests were determined as previously reported (Ventosa et al. 1982). A total of 95 nutritional tests were performed by using the Biolog automatic identification system (Biolog Inc., Hayward, Calif., USA). Strains were grown on isolate medium, at 37 °C for 24 h, and suspended in pre-warmed sterile saline medium (3% NaCl), within the density range specified by the manufacturer with a Biolog photometer model 21101. Immediately after suspending the cells in the saline solution, the suspensions were transferred into sterile multi-channel pipetter reservoirs, and the Biolog GP (for Gram-positive) and GN (for Gram-negative) MicroPlates were inoculated with 125 μl of the cell suspension per well by means of an 8-channel repeating pipetter. The inoculated Biolog plates were incubated at 37 °C for 24 h and the results were read with a MicroPlate Reader using Microlog 3.59 computer software to perform automated reading.
Isolation of genomic DNA and 16S rDNA sequence analysis
DNA of strain CP76 was extracted and precipitated following the CTAB protocol for bacterial genomic DNA preparations (Wilson 1987).
PCR amplification of the 16S rRNA gene by using the forward primer 16F27 and the reverse primer 16R1488 was carried out by using methods that have been previously described in detail (Mellado et al. 1995). Direct sequence determination of the PCR-amplified DNA was carried out using automated DNA cycle sequencing (LiCor). Data analyses were performed using the ARB software package following the method detailed in Arahal et al. (2001).
Enzyme assay
Proteolytic activity was detected by a modified method of Kunitz (1947) as follows. Samples (in duplicates) containing 400 μl of 0.5% (wt/vol) casein (Hammarsten; Merck) in 50 mM Tris-HCl pH 8.5 buffer were incubated in a water bath with 100-μl enzyme sample at optimal temperature and pH for 30–60 min. The enzyme reaction was stopped by the addition of 500 μl of 10% (wt/vol) trichloroacetic acid and was kept at room temperature for another 10 min. The reaction mixture was centrifuged at 13,000 rpm (Heraeus Sepathec, Osterode, Germany) and the absorbance was measured against a blank (non-incubated sample) at 280 nm. One unit of protease was defined as the amount of the enzyme yielding the equivalent of 1 μmol of tyrosine per minute under the defined assay conditions. Protein concentration was determined according to Bradford (1976).
Gel electrophoresis
Native polyacrylamide gels containing a gradient of 4-20% polyacrylamide were purchased from Novex (MBI Fermentas, St. Leon Rot, Germany). Gels were run at 120 V for 16 h at 4 °C. High-molecular-weight marker proteins (Pharmacia Biotech) were used as standards. In order to examine the subunit composition of the protease, protein samples were also analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) as described by Laemmli (1970) after heating the samples at 60 °C for 10 min. Low-molecular-weight marker proteins (Pharmacia Biotech) were used as standards. Following native and SDS-PAGE, the proteins were stained with Commassie blue (0.1%). Zymogram staining for proteolytic activity was detected in situ according to Heussen and Dowdle (1980) with 0.1% (wt/vol) gelatin as the substrate. After electrophoresis the gels were rinsed in 0.25%(vol/vol) Triton X-100 at 4 °C for 1 h to remove SDS and were incubated (10–30 min) under optimal assay conditions to detect the proteolytic activity. The gels were stained in a solution of 0.6% (wt/vol) amido black in water-ethanol-acetic acid (60:30:10) for 1 h and distained in water-ethanol-acetic acid (60:30:10).
Purification of the protease CP1
After cultivation of strain CP76, the culture fluid was centrifuged at 12,000 g for 30 min at 4 °C, and the cell-free supernatant was dialyzed against 100 volumes of 20 mM Tris-HCl, pH 8.5 (buffer A). The supernatant was then loaded onto a Q-Sepharose column (2.5×20 cm) equilibrated in buffer A until no absorbance at 280 nm was detectable. The column was washed with buffer A and proteins were eluted with a linear 0–0.5 M NaCl gradient in buffer A. Active fractions were pooled, concentrated by ultrafiltration, and dialyzed overnight against 100 volumes of 20 mM Tris-HCl pH 8.5 containing 0.1 M NaCl (buffer B). The concentrated sample was applied to a Pharmacia fast protein liquid chromatography Superdex S-200 prep column (1.5×96 cm; Pharmacia Biotech) connected to a Pharmacia FPLC system equilibrated in buffer B. Elution was performed with the same buffer and 1 ml fractions were collected at a flow rate of 1 ml/min. Fractions containing enzyme were analyzed by SDS-12% PAGE.
Influence of pH, temperature, and NaCl
The influence of pH on protease activity was determined by using the same protocol described above, but replacing Tris-HCl buffer with 120 mM Universal buffer, to obtain values from pH 3.5 to 10.0. All of the assays were performed at 55 °C. To determine the influence of temperature on enzymatic activity, samples were incubated at temperatures from 25 °C to 75 °C for 30 min. Thermostability was investigated after incubation of the samples at different temperatures in 120 mM Universal buffer at pH 8.5. After various time intervals, samples were withdrawn and clarified by centrifugation, and the enzymatic activity was measured. The influence of NaCl was determined between 0 and 4 M NaCl using the standard protocol previously described.
Effect of metal ions and inhibitors on protease activity
The effect of various substances on protease activity was examined after incubation of the purified enzyme (0.2 U/ml final concentration) with metal ions and other reagents in various concentrations at 55 °C for 60 min. The protease inhibitors antipain, bestatin, chymostatin, E-64, EDTA-Na, leupeptin, pepstatin, phosphoramidon, Pefabloc, phenylmethylsulfonyl fluoride (PMSF), and aprotinin were added in different final concentrations to the enzymatic mixture. The mixture (450 μl) containing 0.5% casein in 50 mM Tris-HCl buffer pH 8.5 was incubated with 5 μl of enzyme solution for 1 h at room temperature and at 55 °C for 60 min. The residual protease activity was then measured as described before. For determining the influence of metal ions, the same procedure was used, except that the reaction mixture was incubated with the respective salt for 30 min at room temperature and then the activity was detected as described before. The activity without inhibitors or metal ions was considered as 100%.
Amino-terminal analysis
The NH2-terminal sequence determination was carried out with electroblotted samples on PDVF membrane (Matsudaira 1987) by automated Edman degradation on a pulsed liquid sequencer (model 473A, Applied Biosystems, Foster City, Calif.) connected online to an HPLC apparatus for phenylthiohydantoin-derivative identification, following the procedures suggested by the manufacturer.
Results
Screening and characteristics of strains
To test the proteolytic activity of the 310 moderately halophilic strains isolated, a plate assay was performed according to the protocol described in the Materials and methods section. Of the strains tested, 26 exhibited proteolytic activity in milk agar plates. All these proteolytic strains were identified by using the Biolog automatic identification system. Since the Biolog plates for the nutritional tests are different for Gram-negative and Gram-positive bacteria, Gram staining of cells was the first test performed. Of the 26 isolates, 6 were Gram positive and 20 were Gram-negative. According to their phenotypic features, including the ability to oxidize 95 different compounds, the isolates were identified as members of the following groups: 16 strains belonged to the genus Salinivibrio, 4 strains to Bacillus, 2 strains to Salinicoccus, and 4 strains are members of the γ-Proteobacteria (of which 3 strains are Halomonas sp.).
Proteolytic enzyme production
It has been established (Coolbear et al. 1991) that there is not necessarily a good correlation between zones of clearing around colonies on milk-agar plates and levels of proteinase activity produced. To assess proteinase production more quantitatively, the casein assay method was used on culture supernatant fluid of each strain. Results of the assay for proteolytic enzyme production showed that the most active strains were CP52, CP56, and CP76. A partial biochemical characterization of the supernatant of these strains led us to select strain CP76 as the most promising strain for further studies. The supernatant of strain CP76 showed the highest protease activity and exhibited reasonable activity at high salinity (up to 4 M NaCl). The enzyme sample in the supernatant showed highest activity at 1 M NaCl, pH 8.5, and 55 °C.
Characterization of strain CP76
This organism has an optimum growth at a NaCl concentration of around 10%. According to the definition of Kushner (1985), strain CP76 is a moderately halophilic strain. In order to determine the phylogenetic position of strain CP76, 16S rDNA sequencing was performed. Comparative sequence analyses of the 16S rRNA gene of strain CP76 and other 16S rDNA available in the database found the highest similarity (around 90–95%) to the 16S rDNA of strains belonging to the genus Pseudoalteromonas. This result, in addition to phenotypic tests, led us to place this organism in the genus Pseudoalteromonas, as Pseudoalteromonas sp. strain CP76.
Determination of growth conditions for optimal protease activity
To establish the optimal growth conditions yielding the highest extracellular protease activity, the influence of several factors was analyzed. Samples were taken at different times, and the caseinolytic activity of the cell-free medium was measured. No extracellular proteolytic activity was detected during exponential growth; however, during the stationary growth phase, high values were reached.
The highest protease activity was detected when cells were incubated in the saline medium with 7.5% NaCl, and slightly lower activity was detected in 3% or 10% NaCl; however, activity decreased approximately two fold when SW-15 (15% NaCl) was used. Moreover, the effects of different incubation temperatures on protease production were evaluated. Optimal growth was at 37 °C, and at this temperature the highest proteolytic activity was also measured.
To test the influence of different nutrients, strain CP76 was cultured in the presence and absence of these substrates, and both growth and proteolytic activity were determined (Table 1). Highest protease production was obtained when sucrose, fructose, and glycerol were used, whereas maltose repressed the proteolytic activity dramatically. The presence of glucose or lactose had no significant effect. Casamino acids, NH4Cl, mannitol, nutrient broth, and peptone, on the other hand, resulted in good growth but repressed the protease activity. The combination of casamino acids and NH4Cl decreased protease production but not the growth of the bacterium (Table 1).
Purification and properties of the protease
The protease produced by Pseudoalteromoas sp. strain CP76 was purified from a SW-7.5 culture supernatant as indicated in Materials and methods. The procedure of the protease purification is summarized in Table 2. After anion-exchange chromatography and gel filtration, the protease was purified 65-fold with a specific activity of 133 units/mg protein and a final yield of 2.9%. The purified protease showed activity with different substrates, including azocasein (data not shown). SDS-PAGE analysis of the molecular mass of the purified enzyme revealed a single band of 38 kDa (Fig. 1A). A zymogram, shown in Fig. 1B, revealed the high-level activity of the purified protease. A zymogram carried out under native conditions also showed a band of the same molecular mass, indicating that the protease is a monomer of 38 kDa. Kinetic studies using the protease CP1 in the presence of casein as substrate allowed the determination of Michaelis-Menten parameters with a K m value of about 7.1 μM measured at pH 8.5 and 55 °C. The V max was calculated to be 961 U/mg. These results indicate that CP1 is highly active.

A Gel electrophoretic analysis of various purification steps of the protease CP1 produced by Pseudoalteromonas sp. strain CP76: lane 1 molecular markers, lane 2 dialyzed culture supernatant preparation, lane 3 Q-Sepharose column chromatography, lane 4 purified protease after Superdex S-200 column. Proteins were detected with Coomassie Blue (0.1%). B Zymogram of the purified protease as described in Materials and methods; lane 2 purified protease after Superdex S-200 column. Molecular mass markers are indicated in kilodaltons, lane 1
Effects of temperature, pH, and NaCl concentration on the activity and stability of the protease
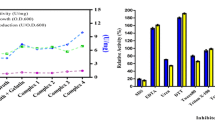
The protease CP1 was active between 25 °C and 75 °C. The optimum activity is at 55 °C, and the enzyme was slightly or moderately thermoactive, retaining 78% and 30% of residual activity at 65 °C and 70 °C, respectively (Fig. 2A).

Effect of temperature (A), pH (B), and NaCl concentration (C) on the activity of the protease CP1. The relative protease activity was defined as the percentage of activity detected with respect to the maximum protease activity detected in the enzymatic assay
The protease showed activity over a broad pH range with an optimum of pH 8.5. Interestingly, the enzyme activity was half-maximal at pH 9.0, and the enzyme retained 30% of initial activity at pH 10.0 (Fig. 2B).
Protease activity was measured at concentrations between 0 and 4 M NaCl, and the maximal activity was detected in the range of 0–1 M NaCl, although at higher salinities (4 M NaCl) activity still remained (Fig. 2C).
Stability of the protease under various conditions was also examined. Samples were incubated for 60, 120, 180, and 240 min in Tris-HCl buffer pH 8.5 and 1 M NaCl at 50 °C and 55 °C, then protease activity was measured under standard conditions using casein as substrate (Fig. 3A). At 50 °C, the purified protease retained 60% and 23% of initial activity after 60 and 240 min of incubation, respectively. Protease stability at different salt concentrations (0.5–4 M NaCl) was determined after pre-incubation of the enzyme at 4 °C for 24 h. As shown in Fig. 3B, the enzyme is completely stable at 1 M and 2 M NaCl, and 60% of activity was measured at 4 M NaCl.

A Thermal stability of protease CP1 at 50 °C (●) and 55 °C (❏). B Stability of protease CP1 at different NaCl concentrations. For thermal stability, the enzyme solution was incubated for 60, 120, 180, and 240 min in 50 mM Tris-HCl + 1 M NaCl pH 8.5. Samples of enzyme solution were removed and cooled on ice, and the residual activity was determined as described in Materials and methods. Stability of the enzyme to NaCl was monitored by incubating the enzyme solution in buffer containing various concentrations of NaCl up to 4 M for 24 h at 4 °C and measuring the residual activity
Effect of different reagents on the activity of the protease
Addition of 0.8 M urea to the protease incubation mixture slightly increased the activity (14%) in comparison to the control. However, a higher concentration (1.6 M) of urea decreased the activity significantly (82%). The stability of the protease was highly affected by the presence of 0.1% and 0.5% SDS, ammonium sulfate (0.2 M and 0.4 M), guanidine-HCl (0.4, 0.8, and 1.6 M), 2-mercaptoethanol (1 and 10 mM), and dithiothreitol (1 and 10 mM) (data not shown).
Influence of different inhibitors on the activity of the protease
The effects of various reagents on purified protease were studied by incubating the enzyme with inhibitors for 1 hour at room temperature (Table 3). Enzyme activity was inhibited to 40% by the metal-ion chelator ethylene diamine tetraacetic acid (EDTA); increasing concentrations of phenylmethylsulfonyl fluoride, a serine protease inhibitor, caused a strong inhibition (87% inhibition at 20 mM PMSF). Similarly, enzyme activity was inhibited by 20% with Pefabloc, another inhibitor of serine proteases. The cysteine protease inhibitors E-64 and leupeptin had no effect. An increase in protease activity was found with phosphoramidon, an inhibitor of metalloendopeptidases. On the other hand, protease activity was not enhanced when different ions were added to the reaction mixture (Table 4), as was expected according to the inhibitory effect caused by the chelating agent EDTA. On the contrary, divalent cations such as Zn2+, Ca2+, Mg2+, or Mn2+ had an inhibitory effect.
Substrate specificity
The purified protease from Pseudoalteromonas sp. strain CP76 showed the highest activity when casein was used as substrate. Lower activity was detected up to 16.2% when gelatin or bovine serum albumin was used as substrate (data not shown).
Amino terminal sequencing
The N-terminal sequence was determined to be A-D-A-T-G-P-G-G-N-Q-K-T-(G-Q-Y-N-Y). Comparison of this sequence with those available in the Swiss-Prot and Genbank databases revealed similarity to metalloproteases from different microorganisms. The highest identity was found to a metalloprotease from an extremophilic bacterium (strain 646) isolated from the Antarctic (AF272770). A high level of similarity was also found to other metalloproteases isolated from other microorganisms (David et al. 1992; Cheng et al. 1996; Chuang et al. 1997; Cascon et al. 2000; Jeong et al. 2000).
Discussion
Proteases from extreme halophiles have been more extensively studied than proteases from moderate halophiles; however, the ability of the latter group of microorganisms to grow in a very wide range of salinities (3–25%) make them very interesting from a biotechnological point of view. Aiming at the discovery of novel proteases, we isolated a total of 310 moderately halophilic strains from solar salterns and saline soils located in Isla Cristina (Huelva, Spain). Interestingly, almost 10% of the strains were found to secrete proteases.
Most strains were Gram-negative rods identified as Salinivibrio, a genus that includes a moderately halophilic species, S. costicola, that is able to produce proteolytic enzymes (García et al. 1987). Other Gram-positive isolates belong to the genera Bacillus or Salinicoccus, while very few are members of the genus Halomonas. This result is surprising if we consider that species of Halomonas are very common inhabitants of the hypersaline environments studied and constitute a large percentage of their microbial population. However, this finding agrees with some preliminary studies that reported the inability of most Halomonas culture collection strains to produce proteases (Coronado 1998).
Strain CP76 was selected for further studies since it appeared to be the best producer of extracellular protease. This organism has an optimum growth at 7.5% NaCl and does not grow without NaCl. According to the definition of Kushner (1985), strain CP76 is a moderately halophilic bacterium. As determined by 16S rDNA sequencing, strain CP76 was closely related to the species of the genus Pseudoalteromonas; however, it was not so similar to any of the species previously described. In addition, the nutritional tests performed are in accordance with the results obtained for other Pseudoalteromonas strains previously described, and thus we placed this organism in the genus Pseudoalteromonas, as Pseudoalteromonas sp. strain CP76. Further studies will be necessary to determine whether it constitutes a new bacterial species.
The production of the protease CP1 by Pseudoalteromonas sp. strain CP76 was strongly influenced by the composition of the culture medium: protease production was optimal in the saline medium supplemented with sucrose, fructose, and glycerol, whereas activity decreased drastically when grown in the presence of maltose. A catabolic repression mechanism for extracellular enzyme production has been reported for various proteases; however, an enhancement of protease activity caused by carbohydrates is not a common behavior.
The highest growth rate was obtained in saline medium supplemented with peptone, although the specific caseinolytic activity decreased by 22.3%. The presence of casamino acids in the culture media increased the growth rates but had negative effects on protease production, although this effect was less extensive than with maltose. Similar behavior, although more marked, has been reported for other proteolytic enzymes of fish pathogens (Secades and Guijarro 1999).
The addition of ammonium to the medium decreased the enzyme activity. A specific repression by ammonium has been reported for several proteases from different microorganisms, such as Aeromonas hydrophila (O'Reilly and Day 1983), A. salmonicida (Liu and Hsieh 1969), or Yersinia ruckeri (Secades and Guijarro 1999).
The salinity of the growth medium strongly influenced the protease activity of Pseudoalteromonas sp. strain CP76. There was a good correlation between the optimal salt concentration for growth and for protease production (7.5% NaCl). Similar behavior has been described for other moderate halophiles producing extracellular amylases (Coronado et al. 2000). However, this is not a general pattern, since in some cases there is no correlation between these two features.
Ion exchange and gel filtration chromatography were used to obtain the protease in its pure form. As judged by SDS-PAGE and native gel analysis (Fig. 1), protease CP1 is a single polypeptide of about 38 kDa. Similar molecular masses presented other halophilic proteases previously characterized (Kamekura et al. 1992; Stepanov et al. 1992; Giménez et al. 2000). Studies to determine temperature dependence revealed that protease CP1 is a moderately thermoactive enzyme (Fig. 2), showing optimal activity at 55 °C. On the other hand, the enzyme was active in a broad pH range (6.0–10.0), showing maximum activity at pH 8.5. Moreover, the protease retained activity in the presence of high salt concentrations (up to 4 M NaCl). Catalytic activity of most proteases from extremely halophilic microorganisms that normally live in a 4-M salt environment falls off dramatically and irreversibly when the enzyme is exposed to lower salt concentrations (Adams and Kelly 1995). The protease CP1, however, is active even without NaCl and in a wide range of NaCl concentrations (up to 4 M NaCl). These characteristics make protease CP1 interesting for application in biotechnological processes. The effect of denaturing and reducing agents on protease CP1 was examined. The proteolytic activity was inhibited by SDS, urea, and guanidine-HCl, thus indicating that hydrogen bonds may be important in maintaining enzyme activity. Furthermore, inhibition by dithiothreitol and 2-mercaptoethanol suggests that disulfide bonds are important in maintaining the molecular conformation required for activity. Enzyme activation by thiol-reducing agent has been reported for a number of extracellular proteases from halophilic bacteria (Giménez et al. 2000). Inhibition of protease CP1 by EDTA suggested that this protease is a metalloprotease. This result is in accordance with the high similarity found between the amino terminal sequence of the purified protein and the metalloproteases from other microorganisms previously described. The amino terminal sequence of protease CP1 displayed the high similarity to a metalloprotease from an extremophilic bacterium isolated from the Antarctic (GenBank accession no. AF272770). Likewise, a high similarity was found to other metalloproteases isolated from Vibrio vulnificus (Cheng et al. 1996; Chuang et al. 1997; Jeong et al. 2000), Vibrio proteolyticus (David et al. 1992), and Aeromonas hydrophila (Cascon et al. 2000). On the other hand, the two inhibitors of serine protease tested, Pefabloc and PMSF, caused a marked inhibition of the activity of the protease. The conjunction of both results suggests the possibility that the protease purified is a serine metalloprotease. The cloning of the protease-encoding gene will allow us to clarify the nature of this novel protease. The comparison of the properties and structures of proteases from the two main groups of microorganisms adapted to live in environments with high salt concentrations, halobacteria and moderately halophilic bacteria, will permit us to determine the different mechanisms of adaptation to salt of these extremophilic microorganisms.
References
Adams MWW, Kelly RM (1995) Enzymes from microorganisms in extreme environments. Chem Eng News 73:32–42
Arahal DR, García MT, Ludwig W, Schleifer KH, Ventosa A (2001) Transfer of Halomonas canadensis and Halomonas israelensis to the genus Chromohalobacter, as Chromohalobacter canadensis comb. nov. and Chromohalobacter israelensis comb. nov., respectively. Int J Syst Evol Microbiol 51:1443–1448
Bradford MM (1976) A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Cascon A, Yugueros J, Temprano A, Sánchez M, Hernanz C, Luengo JM, Naharro G (2000) A major secreted elastase is essential for pathogenicity of Aeromonas hydrophila. Infect Immun 68:3233–3241
Cheng JC, Shao CP, Hor LI (1996) Cloning and nucleotide sequencing of the protease gene of Vibrio vulnificus. Gene 183:255–257
Chuang YC, Chang TM, Chang MC (1997) Cloning and characterization of the gene (empV) encoding extracellular metalloprotease from Vibrio vulnificus. Gene 189:163–168
Coolbear T, Eames CW, Casey Y, Daniel RM, Morgan HW (1991) Screening of strains identified as extremely thermophilic bacilli for extracellular proteolytic activity and general properties of the proteinases from two of the strains. J Appl Bacteriol 71:252–264
Coronado MJ (1998) Transferencia genética en bacterias halófilas moderadas. Caracterización y clonación de una α-amilasa producida por Halomonas meridiana. Ph.D. thesis, University of Seville, Seville, Spain
Coronado MJ, Vargas C, Hofemeister J, Ventosa A, Nieto JJ (2000) Production and biochemical characterization of an -amylase from the moderate halophile Halomonas meridiana. FEMS Microbiol Lett 183:67-71
David VA, Deutch AH, Sloma A, Pawlyk D, Ally A, Durham DR (1992) Cloning, sequencing and expression of the gene encoding the extracellular neutral protease, vibriolysin, of Vibrio protelyticus. Gene 112:107–112
García MT, Ventosa A, Ruíz-Berraquero F, Kocur M (1987) Taxonomic study and amended description of Vibrio costicola. Int J Syst Bacteriol 37:251–256
Giménez MI, Studdert CA, Sánchez JJ, De Castro RE (2000) Extracellular protease of Natrialba magadii: purification and biochemical characterization. Extremophiles 4:181–188
Heussen C, Dowdle EB (1980) Electrophoretic analysis of plasminogen activators in polyacrylamide gels containing sodium dodecyl sulphate and copolymerized substrates. Anal Biochem 102:196–202
Jeong KC, Jeong HS, Rhee JH, Lee SE, Chung SS, Starks AM, Escudero GM, Gulig PA, Choi SH (2000) Construction and phenotypic evaluation of a Vibrio vulnificus vvpE mutant for elastolytic protease. Infect Immun 68:5096–5106
Kamekura M, Seno Y (1990) A halophilic extracellular protease from a halophilic archaebacterium 172 P1. Biochem Cell Biol 68:352–359
Kamekura M, Seno Y, Holmes ML, Dyall-Smith ML (1992) Molecular cloning and sequencing of the gene for a halophilic alkaline serine protease (halolysin) from an unidentified halophilic archaea strain (172P1) and expression of the gene in Haloferax volcanii. J Bacteriol 174:736–742
Kunitz M (1947) Crystalline soybean trypsin inhibitor. II. General properties. J Gen Physiol 30:291–310
Kushner DJ (1985) The Halobacteriaceae. In Woese CR, Wolfe RS (eds) The bacteria, vol 8. Academic, London, pp 171–214
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Liu PV, Hsieh MC (1969) Inhibition of protease production of various bacteria by ammonium salts: its effects on toxin production and virulence. J Bacteriol 99:406–413
Matsudaira P (1987) Sequence from picomole quantities of proteins electroblotted onto polyvinylidene difluoride membranes. J Biol Chem 262:10035–10038
Mellado E, Moore ERB, Nieto JJ, Ventosa A (1995) Phylogenetic inferences and taxonomic consequences of 16S ribosomal DNA sequence comparison of Chromohalobacter marismortui, Volcaniella eurihalina and Deleya salina, and reclassification of V. eurihalina as Halomonas eurihalina comb. nov. Int J Syst Bacteriol 45:712–716
Norberg P, Hofsten B (1969) Proteolytic enzymes from extremely halophilic bacteria. J Gen Microbiol 55:251–256
O´Reilly T, Day F (1983) Effects of culture conditions on protease production by Aeromonas hydrophila. Appl Environ Microbiol 45:1132–1135
Rao MB, Tanksale AM, Ghatge MS, Deshpande VV (1998) Molecular and biotechnological aspects of microbial proteases. Microbiol Mol Biol Rev 62:597–635
Ryu K, Kim J, Dordick JS (1994) Catalytic properties and potential of an extracellular protease from an extreme halophile. Enzyme Microbiol Technol 16:266–275
Secades P, Guijarro JA (1999) Purification and characterization of an extracellular protease from the fish pathogen Yersinia ruckeri and effect of culture conditions on production. Appl Environ Microbiol 65:3969–3975
Stepanov VM, Rudenskaya GN, Revina LP, Gryaznova YB, Lysogorskaya EN, Filippova IY, Ivanova II (1992) A serine proteinase of an archaebacterium, Halobacterium mediterranei homologue of eubacterial subtilisins. Biochem J 285:281–286
Ventosa A, Quesada E, Rodríguez-Valera F, Ruiz-Berraquero F, Ramos-Cormenzana A (1982) Numerical taxonomy of moderately halophilic Gram-negative rods. J Gen Microbiol 128:1959–1968
Ventosa A, Nieto JJ, Oren A (1998) Biology of moderately halophilic aerobic bacteria. Microbiol Mol Biol Rev 62:504–544
Wilson K (1987) Preparation of genomic DNA from bacteria. In: Ausubel FM, Brent R, Kingston RE, Moore DD Seidman JG, Smith JA, Struhl K (eds) Current protocols in molecular biology.Wiley, New York, pp 2.4.1.–2.4.2
Acknowledgments
We gratefully acknowledge Dr. T. Pugsley for his comments on the manuscript. This work was supported by grants from the Spanish Ministerio de Ciencia y Tecnología (PB98-1150 and 1FD97-1162) and Junta de Andalucia. Cristina Sánchez-Porro was supported by a fellowship from the Spanish Ministerio de Ciencia y Tecnología.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by W.D. Grant
Rights and permissions
About this article
Cite this article
Sánchez-Porro, C., Mellado, E., Bertoldo, C. et al. Screening and characterization of the protease CP1 produced by the moderately halophilic bacterium Pseudoalteromonas sp. strain CP76. Extremophiles 7, 221–228 (2003). https://doi.org/10.1007/s00792-003-0316-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00792-003-0316-9