
Abstract
There is evidence to suggest that low levels of magnesium (Mg) are associated with affective disorders, however, causality and central neurobiological mechanisms of this link are largely unproven. We have recently shown that mice fed a low Mg-containing diet (10% of daily requirement) display enhanced depression-like behavior sensitive to chronic antidepressant treatment. The aim of the present study was to utilize this model to gain insight into underlying mechanisms by quantifying amygdala/hypothalamus protein expression using gel-based proteomics and correlating changes in protein expression with changes in depression-like behavior. Mice fed Mg-restricted diet displayed reduced brain Mg tissue levels and altered expression of four proteins, N(G),N(G)-dimethylarginine dimethylaminohydrolase 1 (DDAH1), manganese-superoxide dismutase (MnSOD), glutamate dehydrogenase 1 (GDH1) and voltage-dependent anion channel 1. The observed alterations in protein expression may indicate increased nitric oxide production, increased anti-oxidant response to increased oxidative stress and potential alteration in energy metabolism. Aberrant expressions of DDAH1, MnSOD and GDH1 were normalized by chronic paroxetine treatment which also normalized the enhanced depression-like behavior, strengthening the link between the changes in these proteins and depression-like behavior. Collectively, these findings provide first evidence of low magnesium-induced alteration in brain protein levels and biochemical pathways, contributing to central dysregulation in affective disorders.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Alteration in magnesium (Mg) homeostasis is proposed to be involved in biochemical dysregulations contributing to psychiatric disorders (Murck 2002). In humans, Mg deficiency is common as a significant proportion of people have Mg intakes lower than the recommended daily intake (Galan et al. 1997; Schimatschek and Rempis 2001; for review see Nielsen 2010). In a recent large-scale study, involving approximately 5,700 people, it was found that reduced Mg intake was related to enhanced depression, but not anxiety (Jacka et al. 2009). Furthermore, it was recently reported that decreased intracellular Mg is observed within the anterior cingulate cortex in serotonin-selective re-uptake inhibitor (SSRI) treatment-resistant depressed patients (Iosifescu et al. 2008), which demonstrates for the first time that alteration in brain Mg level is associated with depression. Further, suggesting a role of Mg in mood disorders, it was shown that a reduced serum Mg level is associated with major depressive disorders, suicide attempts as well as mood disorders associated with the menstrual cycle (Banki et al. 1985; Levine et al. 1999; Murck 2002; Rasmussen et al. 1989). Conversely, mood-improving efficacy of Mg supplementation is observed in patients with major depression, traumatic brain injury and postpartum depression (Eby and Eby 2006), and has been shown to result in an anti-depressive phenotype in mice (Poleszak et al. 2004; Poleszak et al. 2007).
It is well established that experimental reduction of Mg levels by Mg-restricted diets elicits enhanced depression-like behavior in rodents (Muroyama et al. 2009; Singewald et al. 2004; Spasov et al. 2008), suggesting a causal relationship between Mg and depression. Evidence suggests that such Mg-restricted diets alter brain Mg levels. Specifically, a 50% reduction in serum Mg concentration has been shown to be associated with a 40% reduction in brain Mg concentration (Altura et al. 1997). However, the underlying neurobiological mechanisms elicited by the Mg-restricted diet leading to enhanced depression are still unknown. Mg is an essential ion and has many diverse functions within the central nervous system including a voltage-dependent block of N-methyl-d-aspartate (NMDA) receptors (Haddad 2005) and being a cofactor in over 300 enzymes including both, tyrosine and tryptophan hydroxylases (Kantak 1988), adenosine triphosphate and enzymes ubiquitous in glycolysis and tricarboxylic acid cycle metabolic pathways (Cowan 2002; Garfinkel and Garfinkel 1985). In energy pathways, energy is released from the ionic species Mg-adenosine triphosphate (Haddad 2005), and interestingly, reduced brain adenosine triphosphate levels have been identified in depressed patients suggesting reduced brain energy metabolism within these patients is associated with depression (Iosifescu et al. 2008). In contrast, it was shown in rodents that chronic dietary Mg-restriction, resulting in a 50% reduction in serum Mg concentration as well as a 40% reduction in brain Mg concentration, does not alter brain adenosine triphosphate concentration (Altura et al. 1997).
In the current experiments we aimed to gain insight into underlying neurobiological changes elicited by a Mg-restricted diet, which has been shown to reduce plasma Mg levels by 45% and to elicit clear depression-like behavior (Singewald et al. 2004). Specifically, we used proteomics to investigate potentially altered expression of soluble proteins in the amygdala/hypothalamus in response to Mg-restriction. Use of proteomics to identify common pathways affected in depression is rare, however, preclinical and clinical proteomic results suggest alteration in signal transduction, energy metabolism and oxidative stress, amongst others, are potentially related to depression (Beasley et al. 2006; Carboni et al. 2006a; Johnston-Wilson et al. 2000; Kromer et al. 2005; Mu et al. 2007). Hence, gel-based proteomics was used to identify novel proteins or signaling pathways altered by a Mg-restricted diet. Quantification of protein expression in the amygdala was chosen (1) because of the importance of this area for depression (Anand and Shekhar 2003; Drevets 2003; Muigg et al. 2007), and (2) based on a preliminary experiment revealing amygdala hyperactivation (assessed by quantifying immediate-early gene expression) in mice fed the Mg-restricted diet compared to normally fed control mice following a forced swim test (Singewald et al. 2010). However, due to methodological sensitivity limitations in the proteomic method, a larger amygdala–hypothalamic area was quantified. To reveal whether our Mg-restricted diet indeed altered Mg levels in this brain area, we quantified Mg tissue content in control fed- and Mg-restricted mice.
Experimental section
Animals
Experiments were carried out in male C57BL/6N mice (Charles River, Germany), 8 weeks old and weighing 19–21 g at the beginning of the experiments. Mice were housed (up to 10 per cage) in a temperature (22 ± 1°C) and humidity (50–60%) controlled vivarium under a 12 h light/dark cycle (lights on at 7 a.m.). All experimental procedures were approved by the Austrian Ethical Committees on Animal Care and Use (Bundesministerium für Wissenschaft und Forschung).
Control and magnesium-restricted diets
Mice assigned to a control diet (n = 18) were allowed to freely take normal mouse chow (ssniff Spezialdiäten, Soest, Germany) that contains 0.2% Mg, which is 4 times more than the minimum Mg requirement of 500 mg/kg of food (Kantak 1988). Mice assigned to the Mg-restricted diet (n = 17) were fed a low Mg-containing diet (50 mg/kg food) which provided about 10% of daily requirements (Kantak 1988) as previously reported (Singewald et al. 2004).
Paroxetine (Sigma-Aldrich, St Louis, MO, USA) was administered to mice fed the Mg-restricted diet via drinking water (n = 18) and the daily dose of 5 mg/kg body weight was calculated based upon drinking water consumption.
Mice were left undisturbed in their home cages for 3 weeks from the commencement of diet and/or drug treatment until the start of experiments. Prior to behavioral testing, mice were allowed to habituate to the test room for at least 24 h.
Behavioral testing
Depression-like behavior
Forced swim test
Mice were subjected to the forced swim test as previously described (Singewald et al. 2004). Mice were individually placed in an open cylinder (diameter 12 cm, height 20 cm) containing 16-cm deep fresh tap water maintained at 23°C. Their activity was videotaped over a period of 6 min. The illumination was set at 100 lux. The total time of immobility was measured during the last 4 min of testing by an observer blinded to the treatments. Mice were considered immobile when floating passively in the water, performing only those movements required for keeping their heads above the water level.
Tail suspension test
Mice were subjected to the tail suspension test as previously described (Whittle et al. 2009). Mice were securely fastened with medical adhesive tape by the tip (c.a. 1.0–1.5 cm) of the tail to a flat metallic surface and suspended for 6 min approximately 30 cm above the surface. The illumination was set at 100 lux. The activity of mice was videotaped over the entire testing period. The total time of immobility was measured during the entire 6 min of testing session by an observer blinded to the treatment. Immobility, defined as when mice hung passively without limb movement, was scored manually.
Locomotor activity
Distance traveled in the open field was quantified as previously described (Tschenett et al. 2003). The open field consisted of a plastic box (41 × 41 × 41 cm) equipped with an automated activity monitoring system (Tru Scan, Coulbourn Instruments, Allentown, USA). Illumination at floor level was 150 lux. Mice were individually placed into the periphery of the open field and their behavior was tracked for 10 min. The overall distance travelled by the mice during the test session was quantified.
Brain dissection
On the fourth day following the forced swim test, mice were killed by carbon dioxide inhalation in a chamber in accordance with established welfare guidelines (Hackbarth et al. 2000). Brains were rapidly removed from the skull and a tissue block containing whole amygdala and hypothalamus (−0.46 mm to −3.40 mm Bregma) was taken on a chilled metal plate and subsequently stored at −80°C; the freezing chain was never interrupted. Amygdala–hypothalamic brain blocks were then subjected to either gel-based proteomic studies (n = 7–8/group) or Western blot analysis (n = 10/group).
Sample preparation for gel-based proteomic studies
Individual tissue blocks of amygdala/hypothalamus were homogenized and suspended in 1.2 mL sample buffer (20 mM Tris, 7 M urea, 2 M thiourea, 4% w/v CHAPS, 10 mM 1,4-dithioerythritol, 1 mM EDTA, 1 mM PMSF, 1 tablet CompleteTM from Roche Diagnostics, and 0.2% v/v phosphatase inhibitor cocktail from Calbiochem). The suspension was sonicated on ice for approximately 30 s and centrifuged at 15,000 × g for 120 min at 12°C. Desalting was carried out with an Ultrafree-4 centrifugal filter unit with a cut-off molecular weight of 10 kDa (Millipore, Bedford, MA, USA) at 3,000 × g at 12°C until the eluted volume was about 4 mL and the remaining volume reached 100–200 μL (Weitzdorfer et al. 2008). The protein content of the supernatant was determined by the Bradford assay.
Two-dimensional gel electrophoresis (2-DE)
Samples of 700 μg protein were subjected to immobilized pH 3–10 nonlinear gradient strips. Focusing started at 200 V and the voltage was gradually increased to 8,000 V at 4 V/min and kept constant for a further 3 h (approximately 150,000 V totally). Prior to the second dimensional run, strips were equilibrated twice for 15 min with gentle shaking in 10 mL of SDS equilibration buffer (50 mM pH 8.8 Tris–HCl, 6 M urea, 30% v/v glycerol, 2% w/v SDS, trace of bromophenol blue). DTT (1%) w/v was added at the first incubation for 15 min and 4% iodoacetamide w/v instead of DTT at the second incubation step for 15 min. The second-dimensional separation was performed on 10–16% gradient SDS-PAGE. After protein fixation for 12 h in 50% methanol and 10% acetic acid, the gels were stained with colloidal Coomassie blue (Novex, San Diego, CA, USA) for 8 h and excess of dye was washed out from the gels with distilled water. Molecular masses were determined by running precision protein standard markers (Bio-Rad Laboratories, Hercules, CA, USA), covering the range of 10–250 kDa. Isoelectric point values were determined as given by the supplier of the immobilized pH gradient strips.
Quantification of protein levels
Protein spots from each gel were outlined (first automatically and then manually) and quantified using the Proteomweaver software (Definiens, Munich, Germany, Burgos et al. 2010). The percentage of the volume of the spots representing a certain protein was determined in comparison with the total proteins present in the 2-DE gel. The software used also revealed that spots evaluated did not contain other proteins. Moreover, only well-separated spots were considered for quantification. Only those proteins (spots) with different levels between groups were identified (Zheng et al. 2009).
Analysis of peptides by nano-LC-ESI-(CID/ETD)-MS/MS (High capacity ion trap, HCT)
Six spots from each group which showed different levels between the three groups were manually excised and placed into 0.5 mL lobind Eppendorf tubes. Gel plugs were washed with 10 mM ammonium bicarbonate and 50% acetonitrile in 10 mM ammonium bicarbonate repeatedly. Addition of 100% acetonitrile resulted in gel shrinking and the shrunk gel plugs were then dried in a Speedvac Concentrator 5301 (Eppendorf, Germany). The dried gel pieces were re-swollen and in-gel digested with 40 ng/μL trypsin (Promega, Madison, WI, USA) in digestion buffer (consisting of 5 mM octyl β-d-glucopyranoside (OGP) and 10 mM ammonium bicarbonate) and incubated over night at 37°C. Chymotrypsin digestion was carried out exactly as published recently (Kang et al. 2009). Peptide extraction was performed with 10 μL of 10 mM ammonium bicarbonate overnight, 15 μL of 1% formic acid (FA) in 5 mM OGP for 30 min, 15 μL of 0.1% FA for 30 min, and subsequently 0.1% FA in 20% acetonitrile (ACN) for 30 min. The extracted peptides were pooled for HCT analysis.
A 40 μL of extracted peptides was analyzed by HCT. The HPLC used was a bio-compatible Ultimate 3000 system (Dionex Corporation, Sunnyvale, CA, USA) equipped with a PepMap100 C-18 trap column (300 μm × 5 mm) and PepMap100 C-18 analytic column (75 μm × 150 mm). The gradient was (A = 0.1% FA in water, B = 0.08% FA in ACN) 4 to 30% B from 0 to 105 min, 80% B from 105 to 110 min, 4% B from 110 to 125 min. The flow rate was 300 nL/min from 0 to 12 min, 75 nL/min from 12 to 105 min, 300 nL/min from 105 to 125 min. A HCT ultra PTM discover system (Bruker Daltonics, Bremen, Germany) was used to record peptide spectra over the mass range of m/z 350–1,500, and MS/MS spectra in information-dependent data acquisition over the mass range of m/z 100–2,800. Repeatedly, MS spectra were recorded followed by three data-dependent CID MS/MS spectra and three ETD MS/MS spectra generated from three highest intensity precursor ions. An active exclusion of 0.4 min after two spectra was used to detect low abundant peptides. The voltage between ion spray tip and spray shield was set to 1,100 V. Drying nitrogen gas was heated to 170°C and the flow rate was 10 L/min. The collision energy was set automatically according to the mass and charge state of the peptides chosen for fragmentation. Multiple charged peptides were chosen for MS/MS experiments due to their good fragmentation characteristics. MS/MS spectra were interpreted and peak lists were generated by DataAnalysis 3.4 (Bruker Daltonics, Bremen, Germany). Searches were done by using the MASCOT 2.2.04 (Matrix Science, London, UK) against latest UniProtKB database for protein identification. Searching parameters were set as follows: enzyme selected as trypsin or chymotrypsin with two maximum missing cleavage sites, species limited to mouse, a mass tolerance of 0.2 Da for peptide tolerance, 0.2 Da for MS/MS tolerance, fixed modification of carbamidomethyl (C) and variable modification of methionine oxidation and phosphorylation (Tyr, Thr, and Ser). Positive protein identifications were based on a significant MOWSE score. After protein identification, an error-tolerant search was done to detect nonspecific cleavage and unassigned modifications. Protein identification and modification information returned from MASCOT were manually inspected and filtered to obtain confirmed protein identification and modification lists of CID MS/MS and ETD MS/MS (Zheng et al. 2009).
Only proteins that were not showing peptides from other proteins were considered.
Western blotting
Aliquots of samples for 2DE were used for Western blotting in order to verify observed DDAH1 levels obtained from 2-DE quantification of amygdala–hypothalamic proteins. Samples (10 μg protein each) were loaded onto 12.5% ExcelGel SDS homogenous gels (Amersham Pharmacia Biotech, Sweden). Electrophoresis was performed with a Multiphor II Electrophoresis System (Amersham Pharmacia Biotech, Sweden). Proteins separated on the gel were transferred onto PVDF membranes (Millipore, MA, USA). Membranes were incubated with a 1:100 diluted primary antibody against DDAH1 (DDAH1, C-19, SC-26068) and a secondary antibody (donkey anti-goat IgG–HRP; diluted 1:2,000, SC-2020, both from Santa Cruz Biotechnology, CA, USA).
Membranes were developed with the Western Lightning™ chemiluminescence reagent (New England Nuclear, MA, USA). Apparent molecular masses were determined by running standard protein markers (Bio-Rad Laboratories Technologies, CA, USA) ranging from 10 to 250 kDa. Arbitrary units of optical density of immunoreactive bands were measured by Image J software program (http://rsb.info.nih.gov/ij/).
Amygdala/hypothalamus Mg determination
Mice, assigned to either a control diet (n = 6) or a Mg-restricted diet (n = 8), were left undisturbed in their home cages for 3 weeks as described above. Mice were then killed by carbon dioxide inhalation in a saturated chamber in accordance with established welfare guidelines (Hackbarth et al. 2000). Brains were rapidly removed from the skull and a tissue block containing whole amygdala and hypothalamus (−0.46 to −3.40 mm Bregma) was taken on a chilled metal plate and subsequently stored at −80°C. Brain tissue was mineralized for 12 h in a 30-fold dilution (wt per vol) of acid solution (3:1 solution of 65% HNO3 and 70% HClO4) at 22 ± 1°C before being further diluted 100-fold with 10% HNO3. Concentration of Mg was analyzed using inductively coupled plasma optical emission spectography (ICP-OES; Philips PU 7000) under standard conditions as previously described (Singewald et al. 2004).
Statistical analysis
All behavioral and protein data were tested for equality of variances using Levene′s test and analyzed using 1-way ANOVA. Where appropriate, post hoc comparison was performed using Bonferroni′s multiple comparison test. Correlations between behavioral output measures and protein expression were performed using the Spearman′s coefficient test. Analysis of Mg tissue levels was performed using student’s t test. A probability level of p < 0.05 was considered as statistically significant.
Results and discussion
This study was undertaken to gain insight into the molecular modifications induced by a Mg-restricted diet. Importantly, no differences in body weight gain (control fed group, 2.7 ± 0.6 g; Mg-restricted diet group, 2.1 ± 0.3 g; Mg-restricted diet group chronically treated with paroxetine, 3.1 ± 0.2 g) or food intake (control fed group, 2.9 ± 0.1 g/mouse per day; Mg-restricted diet group, 2.8 ± 0.1 g/mouse per day; Mg-restricted diet group chronically treated with paroxetine, 2.8 ± 0.2 g/mouse per day) were observed during the course of the experiment. The major finding was that the Mg-restricted diet, which enhanced depression-like behavior, induced altered expression of four proteins associated with nitric oxide signaling, oxidative stress and possibly energy metabolism. The altered expression was normalized after reversal of the enhanced depression-like behavior by chronic paroxetine treatment supporting a relation of the protein changes with depression-like behavior.
Protein identification
Unambiguous identification of proteins is shown in Table 1 providing accession numbers in UniProtKB database, protein name, matched peptides and sequence coverage, peptides used for identification along with information about generation of peptides by trypsin or chymotrypsin, ion score and mass errors in Da. Representative spectra are given in supplementary Fig. 1.
Mg-restricted diet enhances depression-like behavior which is sensitive to chronic paroxetine treatment
Replicating previous reports (Muroyama et al. 2009; Singewald et al. 2004; Spasov et al. 2008), current data demonstrate that a Mg-restricted diet elicits enhanced depression-like behavior. Specifically, increased immobility time was observed in mice fed the Mg-restricted diet compared to control fed mice in both the forced swim and tail suspension tests (Table 2). Importantly, the enhanced depression-like behavior elicited by the Mg-restricted diet was not due to unspecific locomotor effects, indicated by unaltered distance traveled in the open field test (Table 2). The current data further demonstrate the robustness of the Mg-restricted diet model to induce enhanced depression-like behavior in rodents and is congruent with the observation in humans that reduced levels of Mg is associated with pro-depressive behavior (see “Introduction”). Despite this association between results in rodents and humans, it should be kept in mind that the forced swim and tail suspension tests have high predictive validity (Cryan et al. 2002; Petit-Demouliere et al. 2005), but rather limited construct validity (Petit-Demouliere et al. 2005). Hence, additional tests of depression-like behavior including anhedonia, quantified in the sucrose consumption test, should be performed in the future.
We have shown previously that chronic treatment with the tricyclic antidepressant desipramine is effective in rescuing enhanced depression-like behavior elicited by a Mg-restricted diet (Singewald et al. 2004). In the current experiment we have chosen to determine whether chronic treatment with paroxetine, a SSRI (Bourin et al. 2001), is also effective. Chronic paroxetine has been shown to reduce depression-like behavior in mice at concentrations ranging from 1 to 10 mg/kg/day (for example see Elizalde et al. 2008; Gardier et al. 2003; Sillaber et al. 2008). In the current experiment, we have chosen an intermediate dose of 5 mg/kg/day. This dose was sufficient to elicit an antidepressant response in mice fed the Mg-restricted diet as normalization of immobility times in both, the forced swim and tail suspension tests was observed (Table 2). This antidepressive effect was not due to unspecific locomotor effects elicited by chronic paroxetine treatment as locomotor activity, indicated by unaltered distance traveled in the open field test (Table 2). This current result is congruent with the reported antidepressant effect of chronic paroxetine treatment in a rat model of enhanced anxiety and co-morbid depression, which was associated with normalization of central amygdala hyperactivation (Muigg et al. 2007).
Altered protein expression elicited by a Mg-restricted diet
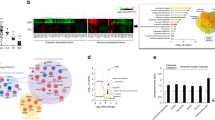
To gain insight into underlying neurobiological changes elicited by a Mg-restricted diet, amygdala/hypothalamus protein expression using gel-based proteomic methods was quantified. This region was chosen on the basis of its importance in depression (see “Introduction”) and the current finding that the Mg-restricted diet reduced Mg tissue concentration in this region (control fed group, 10.9 ± 0.6 μmol/mg tissue; Mg-restricted diet group, 8.8 ± 0.5 μmol/mg tissue; t 12 = 2.60, p < 0.05). To validate the accuracy of the gel-based proteomic experiment, confirmation of a selected protein with altered expression was performed using Western blot. Proteomic quantification of the amygdala/hypothalamus identified 302 individual proteins, represented by 360 protein spots which may reflect the various functional states of the respective gene products (supplementary Table 1). Mice fed a Mg-restricted diet displayed alteration only in four amygdala/hypothalamus proteins compared to fully supplemented control mice; namely N(G),N(G)-dimethylarginine dimethylaminohydrolase 1 (DDAH1; Q9CWS0), manganese superoxide dismutase (MnSOD; SOD2; P09671), voltage-dependent anion channel 1 (VDAC1; Q60932) and glutamate dehydrogenase 1 (GDH1; P26443) (Fig. 1).

An amygdala/hypothalamus protein map of a representative mouse fed the Mg-restricted diet chronically treated with paroxetine on a two-dimensional gel stained with Coomassie blue. Protein spots exhibiting significant alteration in expression either in control, mice fed the Mg-restricted diet and/or mice fed the Mg-restricted diet chronically treated with paroxetine are illustrated. Altered expression was observed in N(G),N(G)-dimethylarginine dimethylaminohydrolase 1 (DDAH1), manganese-superoxide dismutase (MnSOD), voltage-dependent anion channel 1 (VDAC1), glutamate dehydrogenase 1 (GDH1), gamma-enolase (γ-ENO), voltage-dependent anion channel 2 (VDAC2)
Altered nitric oxide signaling
Proteomic quantification revealed that mice fed the Mg-restricted diet displayed reduced expression of DDAH1 compared to control fed mice (Table 3). Western blot quantification validated the proteomic result as it also revealed reduced DDAH1 expression in mice fed the Mg-restricted diet (Table 3; Fig. 2). Importantly, correlation analysis of the whole sample [i.e. control mice, Mg-restricted mice and Mg-restricted mice chronically treated with paroxetine (see below)] revealed a significant negative correlation between measures for depression-like behavior (i.e. immobility time) during both forced swim and tail suspension tests and DDAH1 protein expression in proteomic and Western blot experiments (Fig. 3). As DDAH1 is a signaling protein negatively regulated by nitric oxide (MacAllister et al. 1996), the reduced expression of this protein correlating with enhanced depression-like behavior, may indicate that enhanced central nitric oxide signaling is elicited by dietary induced Mg-restriction. Indeed, a preliminary experiment has revealed that enhanced depression-like behavior elicited by the Mg-restricted diet was antagonized in heterozygous neuronal nitric oxide synthase knock out mice (Singewald et al. 2010) providing behavioral evidence that enhanced nitric oxide signaling elicited by the Mg-restricted diet contributes to the underlying neurobiological mechanisms leading to pro-depression-like behavior. This result is reinforced by findings showing that Mg-restricted diets increase peripheral nitric oxide production in rats (Mak et al. 1996; Rock et al. 1995) which is sensitive to the nitric oxide synthase inhibitor L-NAME (Mak e tal. 1996). Further supporting our collective findings that enhanced nitric oxide signaling contributed to the enhanced depression-like behavior elicited by the Mg-restricted diet, it was recently shown that Flinders rats, a genetic animal model of depression, display increased stress-evoked nitric oxide signaling (Wegener et al. 2010) and that enhanced nitric oxide production, via injection of its precursor l-arginine, elicits enhanced depression-like behavior in rodents (Ergun and Ergun 2007; Inan et al. 2004; Wu 2009; Wu et al. 2001, 2009).

Western blot images of DDAH1 protein expression in the amygdala/hypothalamus in control mice (Ctl), mice fed the Mg-restricted diet (Mg-R) and mice fed the Mg-restricted diet chronically treated with paroxetine (Mg-R PAR)

Correlation analysis between amygdala–hypothalamic protein expression and behavioral measures for depression (i.e. immobility time) in the forced swim and tail suspension tests in control mice (closed circles), mice fed the Mg-restricted diet (closed triangles), and mice fed the Mg-restricted diet chronically treated with paroxetine (open triangles). R correlation coefficient
Within the central nervous system, Mg is present as an inhibitory modulator of NMDA receptor gated channels. Activation of this receptor affects a series of signaling pathways including activation of neuronal nitric oxide synthase and nitric oxide production (Husi and Grant 2001). Thus, enhanced central nitric oxide signaling in Mg-restricted mice may be due to enhanced NMDA receptor activity elicited by the Mg-restricted diet. Supporting evidence for this suggestion comes from ex-vivo brain slice experiments (Libri et al. 1997; Schuchmann et al. 2002).
Altered oxidative stress
In the proteomic experiment, mice fed the Mg-restricted diet displayed increased expression of the anti-oxidant protein MnSOD compared to control fed mice (Table 3). Correlation analysis of the whole sample [i.e. control fed mice, Mg-restricted mice and Mg-restricted mice chronically treated with paroxetine (see below)] revealed a significant positive correlation between measures for depression-like behavior (i.e. immobility time) during both forced swim and tail suspension tests and MnSOD protein expression (Fig. 3). Under physiological conditions there is a balance between oxidative and antioxidative systems, however, when physiological conditions are perturbed towards oxidative systems, oxidative stress, originating from an imbalance between production of reactive oxygen/nitrogen species [including superoxide anion (O −·2 ), hydroxyl radicals (·OH), hydrogen peroxide (H2O2) and peroxynitrite (ONOO−)] and antioxidant capacities of cells results (Hayashi 2009). As MnSOD is an anti-oxidant protein inhibiting accumulation of oxidative stress by reducing superoxide to peroxide (Giulivi et al. 1995) (thus competing with nitric oxide for superoxide to prevent the generation of peroxynitrite, Keller et al. 1998), increased MnSOD expression may be an adaptive change when physiological conditions are perturbed towards oxidative stress (Michel et al. 2004). Indeed, there is evidence that oxidative stress is elicited in peripheral tissue and plasma by Mg-restricted diets as enhanced tissue, erythrocyte and lipoprotein peroxidation (Astier et al. 1996; Bussiere et al. 1995; Freedman et al. 1991, 1992; Gueux et al. 1993; Petrault et al. 2002; Rayssiguier et al. 1993), oxidative modification of proteins (Stafford et al. 1993) and increased plasma nitric oxide production (Mak et al. 1996; Rock et al. 1995) have been reported. Furthermore, a preliminary experiment has revealed that enhanced depression-like behavior elicited by the Mg-restricted diet was antagonized in heterozygous neuronal nitric oxide synthase knock out mice (Singewald et al. 2010) which indicates that enhanced central nitric oxide production, thus oxidative stress, is elicited by the Mg-restricted diet. Our current data showing enhanced MnSOD expression in brain tissue further suggests that enhanced central oxidative stress is elicited in mice fed the Mg-restricted diet as increased MnSOD expression may result as a compensatory mechanism in response to increased oxidative stress. This present result adds to data indicating enhanced central anti-oxidative responses are elicited in rodents fed with a Mg-restricted diet (German-Fattal et al. 2008). Underlining the link to the observed depression-like behavior, evidence for alterations in central oxidative/antioxidative systems is provided in animal models of depression (Carboni et al. 2006, b; Kromer et al. 2005) and in human postmortem tissue (Michel et al. 2007). Taken together, our data provide further evidence for the oxidative stress hypothesis of depression (Michel et al. 2007) which has important implications for understanding the etiology and treatment of depressed patients.
VDAC1, amongst other proteins, is found in the outer mitochondrial membrane and its function is to inhibit cytochrome c release in response to the pro-death Bcl-2 family members Bax and Bid (Baines et al. 2007). We observed that in one out of seven identified VDAC1 gel spots there was enhanced expression in mice fed the Mg-restricted diet (Table 3). The functional relevance of the alteration in expression of this specific VDAC1 protein gel spot remains to be determined.
Altered energy metabolism
In the proteomic experiment, mice fed the Mg-restricted diet displayed reduced protein expression of GDH1 compared to control fed mice (Table 3). Importantly, correlation analysis of the whole sample [i.e. control mice, Mg-restricted mice and Mg-restricted mice chronically treated with paroxetine (see below)] revealed a significant negative correlation between measures for depression-like behavior (i.e. immobility time) both during forced swim and during tail suspension tests and GDH1 protein expression (Fig. 3), suggesting a link between depression-like behavior and GDH1 protein expression. Interestingly, reduced GDH1 protein expression is observed also in the brain (anterior cingulate cortex) of bi-polar patients (Beasley et al. 2006). A relation of GDH levels and energy metabolism has been suggested (Belin et al. 1997) as GDH1 facilitates oxidative transamination of glutamate into α-ketoglutarate, a key intermediate of the tricarboxylic acid cycle (Frigerio et al. 2008). A growing body of evidence suggests that alteration in energy metabolism may contribute to neurobiological mechanisms underlying depression. Studies in animal models of enhanced depression-like behavior show alteration in enzymes associated with the tricarboxylic acid cycle (Carboni et al. 2006a; Mu et al. 2007), which was also found in postmortem samples of depressed patients (Beasley et al. 2006; Johnston-Wilson et al. 2000). Furthermore, non-invasive clinical studies using phosphorus-31 magnetic resonance spectroscopy in patients with mood disorders reveal alteration in markers for energy metabolism. Specifically, reduced baseline levels of β-nucleoside triphosphate and total nucleoside triphosphate, reflecting reduced available adenosine triphosphate, is observed in major depressive patients compared with normal control subjects (Iosifescu and Renshaw 2003). Moreover, a recent clinical study in depressed patients with reduced central Mg concentration revealed reduction in central adenosine triphosphate concentration suggesting reduced brain energy metabolism (Iosifescu et al. 2008). Thus, although this is speculative at the moment, alteration in energy metabolism indicated by reduced protein expression of GDH1 may contribute to the neurobiological mechanisms leading to enhanced depression-like behavior elicited by the Mg-restricted diet.
Effect of chronic paroxetine treatment on altered protein expression elicited by the Mg-restricted diet
In addition to correlating the observed changes in protein expression to changes in depression-like behavior (see above), we wanted to underline the link to depression by experimentally modulating depression-like behavior and investigating the effects on the aberrant expression of the identified proteins. Successful chronic paroxetine treatment which normalized the enhanced depression-like behavior of mice fed the Mg-restricted diet was correlated with alterations in the expression of DDAH1, GDH1 and MnSOD, assessed in the proteomic and Western blot experiments (Table 3; Figs. 2, 3). Specifically, within the proteomic experiment chronic paroxetine treatment increased protein expression in DDAH1 and GDH1 and reduced protein expression in MnSOD to a similar level found in control fed mice (Table 3; Fig. 3). Western blot quantification of DDAH1 protein expression confirmed that chronic paroxetine normalized the expression of this protein (Table 3; Figs. 2, 3).
The exact mechanism(s) by which paroxetine normalized the aberrant protein expression elicited by the Mg-restricted diet is an issue of further investigation. In addition to inhibiting the serotonin re-uptake transporter, SSRIs including paroxetine and fluoxetine have been shown to interact with the nitric oxide system (Crespi 2010; Finkel et al. 1996; Wegener et al. 2003), by reducing brain nitric oxide levels and inhibiting nitric oxide synthase. It is suggested that these effects are important features of the antidepressant activity (Crespi 2010) and that inhibition of nitric oxide formation could be used as a strategy to enhance the clinical efficacy of serotonergic antidepressants (Harkin et al. 2004). Thus, normalization of DDAH1 protein expression may be in part mediated via antagonism of nitric oxide synthase (Finkel et al. 1996) resulting in normalization of nitric oxide signaling. Chronic paroxetine treatment further normalized the expression of MnSOD, potentially indicating restoration in the balance of oxidative and antioxidative systems. This suggestion is supported by a recent preclinical study which has demonstrated reduction in the gene expression of the antioxidant peroxiredoxin-4 following chronic paroxetine treatment (Sillaber et al. 2008), as well as by reports showing that monoamine based antidepressants reduce peripheral markers of oxidative stress (for example, see Kolla et al. 2005; Li et al. 2000; Zafir et al. 2009; Zafir and Banu 2007).
Chronic paroxetine treatment in mice fed the Mg-restricted diet induced expression in two proteins; gamma-enolase (P17183) and voltage-dependent anion channel 2 (Q78MH6) (Table 3; Fig. 3) compared to both control fed mice and mice fed the Mg-restricted diet. It has been suggested that chronic antidepressant treatments may exert their therapeutic effects at the molecular level by stimulating adaptive neuronal plasticity (Duman et al. 1997, 1999; Nestler 1998). Gamma-Enolase is found in mature neurons and neuroendocrine cells (Marangos et al. 1980) and is a multifunctional protein displaying neurotrophic and neuroprotective effects on cultured neurons (Hattori et al. 1995; Takei et al. 1991) in addition to being an enzyme in the tricarboxylic acid cycle (Marangos et al. 1978). Thus, we provide evidence that in an etiologically relevant animal model of enhanced depression-like behavior, an antidepressive response elicited by chronic paroxetine, may enhance neuroplasticity and energy metabolism which may, in part, contribute to mechanisms underlying successful antidepressant treatment. Reasons why increased VDAC2 expression was observed only in the paroxetine treated mice fed the Mg-restricted diet remain unknown as no published report has investigated the interaction of antidepressants and VDAC2. However, increased expression of this protein may be a beneficial action of chronic paroxetine treatment as over-expression of VDAC2 is observed to inhibit BAK-dependent mitochondrial apoptosis (Cheng et al. 2003).
Conclusion
The current study has revealed that enhanced depression-like behavior elicited by feeding mice a Mg-restricted diet was associated with restricted changes in signaling and metabolic protein levels, in addition to reduction in brain Mg tissue levels, in the amygdala/hypothalamus, important in the regulation of depression-like behavior (see “Introduction”). It is intriguing that several proteins with differential expression levels between groups were correlated with key individual parameters for depression-related behavior proposing corresponding tentative functions for these proteins. Specifically, the observed alterations in the expression of proteins indicate increased nitric oxide production, increased anti-oxidant response to increased oxidative stress and potential alteration in energy metabolism. These changes in protein expression elicited by Mg-deficiency were all normalized by chronic paroxetine treatment which also normalized the enhanced depression-like behavior, strengthening the link between changes in these proteins and depression. It must be stated that, at present, we cannot exclude that a combination of behavioral stress and the Mg-restricted diet contributed to the observed alterations in protein expression that was mitigated by chronic paroxetine treatment. This remains subject to further investigations. Furthermore, chronic paroxetine treatment was also associated with increased expression of proteins associated with neuronal plasticity and energy metabolism. Collectively, the present data indicate neurobiological correlates underlying enhanced depression-like behavior elicited by a Mg-restricted diet and reveal putative novel targets for antidepressant therapy.
References
Altura BM, Gebrewold A, Zhang A, Altura BT, Gupta RK (1997) Short-term reduction in dietary intake of magnesium causes deficits in brain intracellular free Mg2+ and [H+]i but not high-energy phosphates as observed by in vivo 31P-NMR. Biochim Biophys Acta 1358:1–5
Anand A, Shekhar A (2003) Brain imaging studies in mood and anxiety disorders: special emphasis on the amygdala. Ann N Y Acad Sci 985:370–388
Astier C, Rock E, Lab C, Gueux E, Mazur A, Rayssiguier Y (1996) Functional alterations in sarcoplasmic reticulum membranes of magnesium-deficient rat skeletal muscle as consequences of free radical-mediated process. Free Radic Biol Med 20:667–674
Baines CP, Kaiser RA, Sheiko T, Craigen WJ, Molkentin JD (2007) Voltage-dependent anion channels are dispensable for mitochondrial-dependent cell death. Nat Cell Biol 9:550–555
Banki CM, Vojnik M, Papp Z et al (1985) Cerebrospinal fluid magnesium and calcium related to amine metabolites, diagnosis, and suicide attempts. Biol Psychiatry 20:163–171
Beasley CL, Pennington K, Behan A, Wait R, Dunn MJ, Cotter D (2006) Proteomic analysis of the anterior cingulate cortex in the major psychiatric disorders: evidence for disease-associated changes. Proteomics 6:3414–3425
Belin MF, Didier-Bazes M, Akaoka H, Hardin-Pouzet H, Bernard A, Giraudon P (1997) Changes in astrocytic glutamate catabolism enzymes following neuronal degeneration or viral infection. Glia 21:154–161
Bourin M, Chue P, Guillon Y (2001) Paroxetine: a review. CNS Drug Rev 7:25–47
Burgos M, Fradejas N, Calvo S, Kang SU, Tranque P, Lubec G (2010) A proteomic analysis of PKCepsilon targets in astrocytes: implications for astrogliosis. Amino Acids (in press)
Bussiere L, Mazur A, Gueux E, Nowacki W, Rayssiguier Y (1995) Triglyceride-rich lipoproteins from magnesium-deficient rats are more susceptible to oxidation by cells and promote proliferation of cultured vascular smooth muscle cells. Magnes Res 8:151–157
Carboni L, Piubelli C, Pozzato C, Astner H, Arban R, Righetti PG, Hamdan M, Domenici E (2006a) Proteomic analysis of rat hippocampus after repeated psychosocial stress. Neuroscience 137:1237–1246
Carboni L, Vighini M, Piubelli C, Castelletti L, Milli A, Domenici E (2006b) Proteomic analysis of rat hippocampus and frontal cortex after chronic treatment with fluoxetine or putative novel antidepressants: CRF1 and NK1 receptor antagonists. Eur Neuropsychopharmacol 16:521–537
Cheng EH, Sheiko TV, Fisher JK, Craigen WJ, Korsmeyer SJ (2003) VDAC2 inhibits BAK activation and mitochondrial apoptosis. Science 301:513–517
Cowan JA (2002) Structural and catalytic chemistry of magnesium-dependent enzymes. Biometals 15:225–235
Crespi F (2010) The selective serotonin reuptake inhibitor fluoxetine reduces striatal in vivo levels of voltammetric nitric oxide (NO): a feature of its antidepressant activity? Neurosci Lett 470:95–99
Cryan JF, Markou A, Lucki I (2002) Assessing antidepressant activity in rodents: recent developments and future needs. Trends Pharmacol Sci 23:238–245
Drevets WC (2003) Neuroimaging abnormalities in the amygdala in mood disorders. Ann N Y Acad Sci 985:420–444
Duman RS, Heninger GR, Nestler EJ (1997) A molecular and cellular theory of depression. Arch Gen Psychiatry 54:597–606
Duman RS, Malberg J, Thome J (1999) Neural plasticity to stress and antidepressant treatment. Biol Psychiatry 46:1181–1191
Eby GA, Eby KL (2006) Rapid recovery from major depression using magnesium treatment. Med Hypotheses 67:362–370
Elizalde N, Gil-Bea FJ, Ramirez MJ, Aisa B, Lasheras B, Del Rio J, Tordera RM (2008) Long-lasting behavioral effects and recognition memory deficit induced by chronic mild stress in mice: effect of antidepressant treatment. Psychopharmacology (Berl) 199:1–14
Ergun Y, Ergun UG (2007) Prevention of pro-depressant effect of L-arginine in the forced swim test by NG-nitro-L-arginine and [1H-[1, 2, 4]Oxadiazole[4, 3-a]quinoxalin-1-one]. Eur J Pharmacol 554:150–154
Finkel MS, Laghrissi-Thode F, Pollock BG, Rong J (1996) Paroxetine is a novel nitric oxide synthase inhibitor. Psychopharmacol Bull 32:653–658
Freedman AM, Cassidy MM, Weglicki WB (1991) Magnesium-deficient myocardium demonstrates an increased susceptibility to an in vivo oxidative stress. Magnes Res 4:185–189
Freedman AM, Mak IT, Stafford RE, Dickens BF, Cassidy MM, Muesing RA, Weglicki WB (1992) Erythrocytes from magnesium-deficient hamsters display an enhanced susceptibility to oxidative stress. Am J Physiol 262:C1371–C1375
Frigerio F, Casimir M, Carobbio S, Maechler P (2008) Tissue specificity of mitochondrial glutamate pathways and the control of metabolic homeostasis. Biochim Biophys Acta 1777:965–972
Galan P, Preziosi P, Durlach V, Valeix P, Ribas L, Bouzid D, Favier A, Hercberg S (1997) Dietary magnesium intake in a French adult population. Magnes Res 10:321–328
Gardier AM, David DJ, Jego G, Przybylski C, Jacquot C, Durier S, Gruwez B, Douvier E, Beauverie P, Poisson N, Hen R, Bourin M (2003) Effects of chronic paroxetine treatment on dialysate serotonin in 5-HT1B receptor knockout mice. J Neurochem 86:13–24
Garfinkel L, Garfinkel D (1985) Magnesium regulation of the glycolytic pathway and the enzymes involved. Magnesium 4:60–72
German-Fattal M, Lecerf F, Sabbagh F, Maurois P, Durlach J, Bac P(2008) Neuroprotective gene profile in the brain of magnesium-deficient mice. Biomed Pharmacother 62:264–272
Giulivi C, Boveris A, Cadenas E (1995) Hydroxyl radical generation during mitochondrial electron transfer and the formation of 8-hydroxydesoxyguanosine in mitochondrial DNA. Arch Biochem Biophys 316:909–916
Gueux E, Cubizolles C, Bussiere L, Mazur A, Rayssiguier Y (1993) Oxidative modification of triglyceride-rich lipoproteins in hypertriglyceridemic rats following magnesium deficiency. Lipids 28:573–575
Hackbarth H, Kuppers N, Bohnet W (2000) Euthanasia of rats with carbon dioxide—animal welfare aspects. Lab Anim 34:91–96
Haddad JJ (2005) N-methyl-D-aspartate (NMDA) and the regulation of mitogen-activated protein kinase (MAPK) signaling pathways: a revolving neurochemical axis for therapeutic intervention? Prog Neurobiol 77:252–282
Harkin A, Connor TJ, Burns MP, Kelly JP (2004) Nitric oxide synthase inhibitors augment the effects of serotonin re-uptake inhibitors in the forced swimming test. Eur Neuropsychopharmacol 14:274–281
Hattori T, Takei N, Mizuno Y, Kato K, Kohsaka S (1995) Neurotrophic and neuroprotective effects of neuron-specific enolase on cultured neurons from embryonic rat brain. Neurosci Res 21:191–198
Hayashi M (2009) Oxidative stress in developmental brain disorders. Neuropathology 29:1–8
Husi H, Grant SG (2001) Proteomics of the nervous system. Trends Neurosci 24:259–266
Inan SY, Yalcin I, Aksu F (2004) Dual effects of nitric oxide in the mouse forced swimming test: possible contribution of nitric oxide-mediated serotonin release and potassium channel modulation. Pharmacol Biochem Behav 77:457–464
Iosifescu DV, Renshaw PE (2003) 31P-magnetic resonance spectroscopy and thyroid hormones in major depressive disorder: toward a bioenergetic mechanism in depression? Harv Rev Psychiatry 11:51–63
Iosifescu DV, Bolo NR, Nierenberg AA, Jensen JE, Fava M, Renshaw PF (2008) Brain bioenergetics and response to triiodothyronine augmentation in major depressive disorder. Biol Psychiatry 63:1127–1134
Jacka FN, Overland S, Stewart R, Tell GS, Bjelland I, Mykletun A (2009) Association between magnesium intake and depression and anxiety in community-dwelling adults: the Hordaland Health Study. Aust N Z J Psychiatry 43:45–52
Johnston-Wilson NL, Sims CD, Hofmann JP, Anderson L, Shore AD, Torrey EF, Yolken RH (2000) Disease-specific alterations in frontal cortex brain proteins in schizophrenia, bipolar disorder, and major depressive disorder. The Stanley Neuropathology Consortium. Mol Psychiatry 5:142–149
Kang SU, Fuchs K, Sieghart W, Pollak A, Csaszar E, Lubec G (2009) Gel-based mass spectrometric analysis of a strongly hydrophobic GABAA-receptor subunit containing four transmembrane domains. Nat Protoc 4:1093–1102
Kantak KM (1988) Magnesium deficiency alters aggressive behavior and catecholamine function. Behav Neurosci 102:304–311
Keller JN, Kindy MS, Holtsberg FW, St Clair DK, Yen HC, Germeyer A, Steiner SM, Bruce-Keller AJ, Hutchins JB, Mattson MP (1998) Mitochondrial manganese superoxide dismutase prevents neural apoptosis and reduces ischemic brain injury: suppression of peroxynitrite production, lipid peroxidation, and mitochondrial dysfunction. J Neurosci 18:687–697
Kolla N, Wei Z, Richardson JS, Li XM (2005) Amitriptyline and fluoxetine protect PC12 cells from cell death induced by hydrogen peroxide. J Psychiatry Neurosci 30:196–201
Kromer SA, Kessler MS, Milfay D, Birg IN, Bunck M, Czibere L, Panhuysen M, Putz B, Deussing JM, Holsboer F, Landgraf R, Turck CW (2005) Identification of glyoxalase-I as a protein marker in a mouse model of extremes in trait anxiety. J Neurosci 25:4375–4384
Levine J, Stein D, Rapoport A, Kurtzman L (1999) High serum and cerebrospinal fluid Ca/Mg ratio in recently hospitalized acutely depressed patients. Neuropsychobiology 39:63–70
Li XM, Chlan-Fourney J, Juorio AV, Bennett VL, Shrikhande S, Bowen RC (2000) Antidepressants upregulate messenger RNA levels of the neuroprotective enzyme superoxide dismutase (SOD1). J Psychiatry Neurosci 25:43–47
Libri V, Santarelli R, Nistico S, Azzena GB (1997) Inhibition of nitric oxide synthase prevents magnesium-free-induced epileptiform activity in guinea-pig piriform cortex neurones in vitro. Naunyn Schmiedebergs Arch Pharmacol 355:452–456
MacAllister RJ, Parry H, Kimoto M, Ogawa T, Russell RJ, Hodson H, Whitley GS, Vallance P (1996) Regulation of nitric oxide synthesis by dimethylarginine dimethylaminohydrolase. Br J Pharmacol 119:1533–1540
Mak IT, Komarov AM, Wagner TL, Stafford RE, Dickens BF, Weglicki WB (1996) Enhanced NO production during Mg deficiency and its role in mediating red blood cell glutathione loss. Am J Physiol 271:C385–C390
Marangos PJ, Zis AP, Clark RL, Goodwin FK (1978) Neuronal, non-neuronal and hybrid forms of enolase in brain: structural, immunological and functional comparisons. Brain Res 150:117–133
Marangos PJ, Schmechel DE, Parma AM, Goodwin FK (1980) Developmental profile of neuron-specific (NSE) and non-neuronal (NNE) enolase. Brain Res 190:185–193
Michel TM, Thome J, Martin D, Nara K, Zwerina S, Tatschner T, Weijers HG, Koutsilieri E (2004) Cu, Zn- and Mn-superoxide dismutase levels in brains of patients with schizophrenic psychosis. J Neural Transm 111:1191–1201
Michel TM, Frangou S, Thiemeyer D, Camara S, Jecel J, Nara K, Brunklaus A, Zoechling R, Riederer P (2007) Evidence for oxidative stress in the frontal cortex in patients with recurrent depressive disorder–a postmortem study. Psychiatry Res 151:145–150
Mu J, Xie P, Yang ZS, Yang DL, Lv FJ, Luo TY, Li Y (2007) Neurogenesis and major depression: implications from proteomic analyses of hippocampal proteins in a rat depression model. Neurosci Lett 416:252–256
Muigg P, Hoelzl U, Palfrader K, Neumann I, Wigger A, Landgraf R, Singewald N (2007) Altered brain activation pattern associated with drug-induced attenuation of enhanced depression-like behavior in rats bred for high anxiety. Biol Psychiatry 61:782–796
Murck H (2002) Magnesium and affective disorders. Nutr Neurosci 5:375–389
Muroyama A, Inaka M, Matsushima H, Sugino H, Marunaka Y, Mitsumoto Y (2009) Enhanced susceptibility to MPTP neurotoxicity in magnesium-deficient C57BL/6N mice. Neurosci Res 63:72–75
Nestler EJ (1998) Antidepressant treatments in the 21st century. Biol Psychiatry 44:526–533
Nielsen FH (2010) Magnesium, inflammation, and obesity in chronic disease. Nutr Rev 68:333–340
Petit-Demouliere B, Chenu F, Bourin M (2005) Forced swimming test in mice: a review of antidepressant activity. Psychopharmacology (Berl) 177:245–255
Petrault I, Zimowska W, Mathieu J, Bayle D, Rock E, Favier A, Rayssiguier Y, Mazur A (2002) Changes in gene expression in rat thymocytes identified by cDNA array support the occurrence of oxidative stress in early magnesium deficiency. Biochim Biophys Acta 1586:92–98
Poleszak E, Szewczyk B, Kedzierska E, Wlaz P, Pilc A, Nowak G (2004) Antidepressant- and anxiolytic-like activity of magnesium in mice. Pharmacol Biochem Behav 78:7–12
Poleszak E, Wlaz P, Kedzierska E, Nieoczym D, Wrobel A, Fidecka S, Pilc A, Nowak G (2007) NMDA/glutamate mechanism of antidepressant-like action of magnesium in forced swim test in mice. Pharmacol Biochem Behav 88:158–164
Rasmussen HH, Mortensen PB, Jensen IW (1989) Depression and magnesium deficiency. Int J Psychiatry Med 19:57–63
Rayssiguier Y, Gueux E, Bussiere L, Durlach J, Mazur A (1993) Dietary magnesium affects susceptibility of lipoproteins and tissues to peroxidation in rats. J Am Coll Nutr 12:133–137
Rock E, Astier C, Lab C, Malpuech C, Nowacki W, Gueux E, Mazur A, Rayssiguier Y (1995) Magnesium deficiency in rats induces a rise in plasma nitric oxide. Magnes Res 8:237–242
Schimatschek HF, Rempis R (2001) Prevalence of hypomagnesemia in an unselected German population of 16, 000 individuals. Magnes Res 14:283–290
Schuchmann S, Albrecht D, Heinemann U, und Halbach O (2002) Nitric oxide modulates low-Mg2+-induced epileptiform activity in rat hippocampal-entorhinal cortex slices. Neurobiol Dis 11:96–105
Sillaber I, Panhuysen M, Henniger MS, Ohl F, Kuhne C, Putz B, Pohl T, Deussing JM, Paez-Pereda M, Holsboer F (2008) Profiling of behavioral changes and hippocampal gene expression in mice chronically treated with the SSRI paroxetine. Psychopharmacology (Berl) 200:557–572
Singewald N, Sinner C, Hetzenauer A, Sartori SB, Murck H (2004) Magnesium-deficient diet alters depression- and anxiety-related behavior in mice—influence of desipramine and Hypericum perforatum extract. Neuropharmacology 47:1189–1197
Singewald N, Sartori SB, Shin JH, Lin L, Lubec G, Whittle N (2010) Magnesium- and zinc-deficiency models for depression: Involvement of NMDA/NO pathways. Biol Psychiatry 67:689
Spasov AA, Iezhitsa IN, Kharitonova MV, Kravchenko MS (2008) Depression-like and anxiety-related behaviour of rats fed with magnesium-deficient diet. Zh Vyssh Nerv Deiat Im I P Pavlova 58:476–485
Stafford RE, Mak IT, Kramer JH, Weglicki WB (1993) Protein oxidation in magnesium deficient rat brains and kidneys. Biochem Biophys Res Commun 196:596–600
Takei N, Kondo J, Nagaike K, Ohsawa K, Kato K, Kohsaka S (1991) Neuronal survival factor from bovine brain is identical to neuron-specific enolase. J Neurochem 57:1178–1184
Tschenett A, Singewald N, Carli M, Balducci C, Salchner P, Vezzani A, Herzog H, Sperk G (2003) Reduced anxiety and improved stress coping ability in mice lacking NPY-Y2 receptors. Eur J Neurosci 18:143–148
Wegener G, Volke V, Harvey BH, Rosenberg R (2003) Local, but not systemic, administration of serotonergic antidepressants decreases hippocampal nitric oxide synthase activity. Brain Res 959:128–134
Wegener G, Harvey BH, Bonefeld B, Müller HK, Volke V, Overstreet DH, Elfving B (2010) Increased stress-evoked nitric oxide signalling in the Flinders sensitive line (FSL) rat: a genetic animal model of depression. Int J Neuropsychopharmacol 13:461–473
Weitzdorfer R, Hoger H, Burda G, Pollak A, Lubec G (2008) Differences in hippocampal protein expression at 3 days, 3 weeks, and 3 months following induction of perinatal asphyxia in the rat. J Proteome Res 7:1945–1952
Whittle N, Lubec G, Singewald N (2009) Zinc deficiency induces enhanced depression-like behaviour and altered limbic activation reversed by antidepressant treatment in mice. Amino Acids 36:147–158
Wu G (2009) Amino acids: metabolism, functions, and nutrition. Amino Acids 37:1–17
Wu G, Haynes TE, Li H, Yan W, Meininger CJ (2001) Glutamine metabolism to glucosamine is necessary for glutamine inhibition of endothelial nitric oxide synthesis. Biochem J 353:245–252
Wu G, Bazer FW, Davis TA, Kim SW, Li P, Marc Rhoads J, Carey Satterfield M, Smith SB, Spencer TE, Yin Y (2009) Arginine metabolism and nutrition in growth, health and disease. Amino Acids 37:153–168
Zafir A, Banu N (2007) Antioxidant potential of fluoxetine in comparison to Curcuma longa in restraint-stressed rats. Eur J Pharmacol 572:23–31
Zafir A, Ara A, Banu N (2009) Invivo antioxidant status: a putative target of antidepressant action. Prog Neuropsychopharmacol Biol Psychiatry 33:220–228
Zheng J, Patil SS, Chen WQ, An W, He J, Hoger H, Lubec G (2009) Hippocampal protein levels related to spatial memory are different in the Barnes maze and in the Multiple T-maze. J Proteome Res 8:4479–4486
Acknowledgments
The authors thank Dr. Richard Tessadri for the determination of Mg levels. This work was funded by the FWF (P22931-B18).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
N. Whittle and L. Li have contributed equally to this article.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Whittle, N., Li, L., Chen, WQ. et al. Changes in brain protein expression are linked to magnesium restriction-induced depression-like behavior. Amino Acids 40, 1231–1248 (2011). https://doi.org/10.1007/s00726-010-0758-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00726-010-0758-1