
Abstract
Viral gastroenteritis is a major public health problem worldwide. In Ethiopia, very limited studies have been done on the epidemiology of enteropathogenic viruses. The aim of this study was to detect and characterize noroviruses (NoVs) and sapoviruses (SaVs) from acute gastroenteritis patients of all ages. Fecal samples were collected from diarrheic patients (n = 213) in five different health centers in Addis Ababa during June-September 2013. The samples were screened for caliciviruses by reverse transcription polymerase chain reaction (RT-PCR) using universal and genogroup-specific primer pairs. Phylogenetic analyses were conducted using the sequences of the PCR products. Of the clinical samples, 25.3 % and 4.2 % were positive for NoV and SaV RNA, respectively. Among the norovirus positives, 22 were sequenced further, and diverse norovirus strains were identified: GI (n = 4), GII (n = 17) and GIV (n = 1). Most strains were GII (n = 17/22: 77.2 %), which were further divided into three different genotypes (GII.4, GII.12/GII.g recombinant-like and GII.17), with GII.17 being the dominant (7/17) strain detected. GI noroviruses, in particular GI.4 (n = 1), GI.5 (n = 2) and GI.8 (n = 1), were also detected and characterized. The GIV strain detected is the first from East Africa. The sapoviruses sequenced were also the first reported from Ethiopia. Collectively, this study showed the high burden and diversity of noroviruses and circulation of sapoviruses in diarrheic patients in Ethiopia. Continued surveillance to assess their association with diarrhea is needed to define their epidemiology, disease burden, and impact on public health.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Acute gastroenteritis (AGE) remains one of the major causes of death in children under the age of five worldwide. It results in 1.8 million fatalities annually in developing countries alone [1] and is associated with about 14 % of deaths in children less than five years of age [2] in Ethiopia. Noroviruses and sapoviruses, members of the family Caliciviridae, are responsible for the majority of gastroenteritis cases. Norovirus, is the leading cause of gastroenteritis in people of all ages and is the second most common cause of diarrhea in children under the age of five, worldwide [3, 4]. In countries where universal rotavirus vaccination has been established, NoVs have become the leading cause of admission to hospital with acute gastroenteritis, also for children <5 years of age [5–8]. Noroviruses and sapoviruses are small, non-enveloped, single-stranded, positive-sense, polyadenylated RNA viruses with a genome of 7.4-7.7 kb in size [9]. Their genome is organized into two to three open reading frames (ORFs), which encode both the structural and the non-structural protein genes, including the RNA-dependent RNA polymerase (RdRp) and the major capsid protein (VP1) [10, 11].
The NoVs are genetically highly diverse, which is attributed to the accumulation of point mutations associated with error-prone RNA replication and recombination. Currently, they are classified into at least five genogroups (GI-V) based on the complete sequence of the capsid protein VP1 [11–13]. Genogroup I (GI), GII, and GIV viruses are primarily human pathogens, although there are three porcine-specific genotypes within the GII genogroup [11, 12]. Comparison of full-length capsid sequences demonstrated that the five different genogroups share >60 % amino acid sequence identity in the VP1 region [12]. Each genogroup is subdivided further into distinct genetic clusters or genotypes that share >80 % identity in the amino acid sequence of VP1 [12]. Due to frequent recombination of NoVs, a dual classification/nomenclature system relating to the diversity of the RdRp and capsid genes has been proposed and appears to be the way forward [14, 15]. The other pathogenic enteric calicivirus, SaV, is classified into five genogroups based on capsid gene (ORF1) sequences, genogroups GI to GV [16]; however, potential new genogroups have been proposed [17–19]. Genogroups GI, GII, GIV, and GV contain the human SaVs. Each genogroup is further divided into genetically diverse genotypes.
The role of NoVs and SaVs as causative agents of gastroenteritis and their diversity in Africa is not well studied, except in some reports from a few African countries [20–36]. There has been only one published study from Ethiopia regarding the presence and genetic diversity of NoVs [37], and none for SaVs to date. Thus, this study focuses on the prevalence and genetic characterization of human NoVs and SaVs in Ethiopia.
Materials and methods
The study design for this work was cross-sectional. The study was approved by the National Health Research Ethics Committee and Institutional Review Board of Aklilu Lemma Institute of Pathobiology. The clinical samples (fecal samples, n = 213) investigated in this study were collected from self-reporting acute gastroenteritis patients attending five different government health care centers in Addis Ababa (Arada, Semen, Lideta, Kolfe and Kirkos Health Centers), between June to September 2013. Patients of all ages with diarrhea and one or more of the following symptoms – nausea, vomiting, abdominal cramps, headache, muscle pain and/or fever – were included. Addis Ababa is the capital and largest city of Ethiopia. According to the 2007 census, 98.6 % of the housing units of Addis Ababa had access to safe chlorinated drinking water, 14.9 % had flush toilets, 70.7 % had pit toilets, and 14.3 % had no toilet facilities [38].
Molecular detection and characterization
Fecal suspension preparation and RNA extraction
Fresh stool samples were collected from human acute gastroenteritis patients, using sterile containers, and stored frozen at -70 °C until processed. Ten-percent (w/v) fecal suspensions were prepared in phosphate-buffered saline (PBS, 0.01 M, pH 7.2) and clarified at 2000 × g (20 min, 4 °C) to eliminate larger debris. Viral RNA was extracted from 250 µl fecal suspensions using an RNeasy Mini Kit (QIAGEN, Valencia, CA) according to the manufacturer’s instruction. It was treated with 5 U of DNase I (Invitrogen, Carlsbad, CA, USA) to remove DNA. The extracted RNA was stored at -70 °C until used.
Reverse transcription
The cDNA was synthesized from the extracted RNA using a MAXIMA First Strand cDNA Synthesis Kit (Thermo Scientific) according to the manufacturer’s instructions. The kit contains highly sensitive primer mixture oligo (dT)18 and random hexamer. The cDNA obtained was used for the subsequent PCR assays [39].
PCR and primers
Human NoVs were detected and characterized by RT-PCR followed by sequencing using a calicivirus universal primer pair and genogroup-specific primer pairs. The universal primer pair (p290/p110) targets a conserved region in the RdRp [40, 41], which correspond to nucleotides (nt) 4568-4590 (p290) and nt 4865 to 4884 (p110) in the prototype NoV genome sequence (Hu/NoV/GI.1/Norwalk, GenBank accession no. M87661). The genogroup-specific primer pairs G1SKF/G1SKR and G2SKF/G2SKR [42], which amplified the 5’ end of the capsid gene, were also used for the detection of human GI and GII NoVs, respectively. The genogroup-specific primer pair PEC66/PEC65, which is often used to detect porcine SaV GIII [43], was also employed in this study to detect potential interspecies transmission. The primer pairs used to detect NoVs and SaVs are listed in Table 1. The PCR conditions were optimized for all primers and the specificity of the primers was evaluated using plasmid DNA of laboratory reference strains of calicivirus obtained from Linda Saif’s laboratory at the Ohio State University, which included Hu/NoV/GII.4/HS194. All RT-PCR reactions were conducted individually with each primer pair using various PCR cycling parameters. Positive controls (NoV and SaV) and negative controls (RNase-free water) were included in each run. The amount of PCR inhibitors in the samples was determined to be negligible based on results obtained by making 10-fold and 100-fold dilutions of each RNA sample and using an internal control as described previously [17]. Conditions for PCR were as follows: initial denaturation at 94 °C for 3 min, followed by 40 cycles of 94 °C for 30 s, 48 °C for 30 s, and 72 °C for 1 min, with a final extension step at 72 °C for 10 min for primer pair p290/p110 and 94 °C for 3 min followed by 40 cycles of 94 °C 30 s, and 55 °C and 72 °C for 1 min for primer pairs G1SKF/G1SKR and G2SKF/G2SKR. The amplicons obtained were analyzed in agarose gels after staining with GelRed dye (Thermo Scientific) using a UV light transilluminator.
The PCR products were separated by electrophoresis in an agarose gel, and samples were purified using a QIAquick Gel Extraction Kit (QIAGEN) before direct sequencing according to manufacturer’s instructions. Sanger sequencing of the RT-PCR products was carried out at the facilities of Segolip Unit of BecA-ILRI Hub, Nairobi, using an ABI 3730 automatic capillary sequencer (Applied Biosystem, Foster City, CA, USA). Complete sequences were obtained by assembling overlapping contigs followed by trimming off primer sequences. The nucleotide sequences obtained from the selected strains were used to search similar sequences in the NCBI genetic database using the BLAST tool (available at http://www.ncbi.nlm.nih.gov/blast/ Blast.cgi). The DNA sequences were aligned using the ClustalW method [44]. Phylogenetic trees were constructed by the neighbor-joining method using DNASTAR and MEGA6 software, with 1,000 bootstrap replicates for each gene [44]. The evolutionary history was inferred using the neighbor-joining method [45]. The tree was drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Tamura-Nei method and are in units of the number of base substitutions per site. Evolutionary analyses were conducted using MEGA6 [44].
Statistical analysis
Data were analyzed using SPSS version 15 software. Proportions were used to summarize the frequency of occurrence of target outcomes in different categories of exposure variables. The precision of the prevalence of enteric viruses across different categories was assessed by determining the 95 % confidence interval around the estimates. The statistical significance of differences of proportion was evaluated using the Chi-square test. A P-value <0.05 was considered an indicator of statistical significance.
Results
Profiles of the study population
During the study period, 213 individuals were enrolled and screened for NoVs and SaVs. All of the participants were residents of the city of Addis Ababa, and 54.9 % were female, resulting in a female-to-male ratio of 1.22:1. The age distribution of the study participants ranged from 4 months to 73 years. The proportion of the participants by age and sex are summarized in Tables 2 and 3.
Prevalence
Norovirus infection
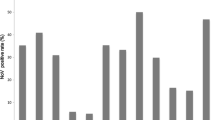
Of the 213 diarrheic patients screened by RT-PCR, 25.3 % (54/213) were positive for NoV RNA (Table 4). Using primer pairs G1SKF/G1SKR and G2SKF/G2SKR, which were designed to detect human NoV GI and GII, respectively, 47 NoV strains were detected (explained in Table 5). For samples that were negative using the NoV genogroup-specific primer pairs, we next employed the calicivirus universal primer pair p290/p110 [40, 41]. Seven additional samples were positive using the primer pair p290/p110, and all were sequenced and identified as human NoVs (Table 5). We also analyzed the prevalence of NoVs in relation to the age of the study participants. The prevalence of NoV differed among age groups, ranging from 9.5 to 42.6 % in subjects >40 years and 1-4 years, respectively (Table 3). Statistical analysis showed that the frequency of human NoV infection was strongly correlated with the age (1-4 years) of the children (p = 0.007).
Sapovirus infection
For SaV detection, we first employed the calicivirus universal primer pair targeting the RdRp region p290/p110, which failed to detect SaV RNA in the present study. We next used the genogroup-specific primer pair PEC66/PEC65, whereby nine of the samples were positive for SaV RNA (4.2 %) (9/213) (Table 3). SaV was detected in four of the five health centers at different percentages, ranging from 1.6 to 8 % (Table 4). The percentage of the samples positive by RT-PCR in the study subjects also differed among age groups: 18.2 % in infants <1 year; 4.3 % in children 1-4 years; 7.9 % in subjects 10-19 years; 3.7 % in adults 30-39 years and 4.76 % in adults >40 years of age.
Genetic analysis of noroviruses
Phylogenetic analysis of a portion of the RNA polymerase region
Seven of the 54 positive samples detected using the primer pair p290/p110 targeting the RNA polymerase region were sequenced (Table 5). After performing a BLAST search, the sequenced strains were identified as human NoVs (ZS-AH21-ETH-2013, ZS-AH74-ETH-2013, ZS-KH05-ETH-2013, ZS-KH21-ETH-2013, ZS-AH12-ETH-2013, ZS-L6-ETH-2013, ZS-AH35-ETH-2013) (Fig. 1). The GenBank accession numbers of the partial NoV RdRp-specific sequences are KM589655, KM589656, KM589657, KM589658, KM589659, KM589660, and KM589661, respectively. A phylogenetic tree was generated using the partial RdRp region of seven NoV sequences that we determined, together with published sequences of NoV reference and representative strains. A neighbor-joining tree based on the partial RdRp gene and BLAST search indicated that the seven sequences we obtained were potential NoV strains of groups GII and GIV. Most of the strains (75 %, 6/7) that were identified were of genogroup GII, with nucleotide identities ranging from 67 to 100 % (Fig. 1). The GII strains were further divided into three potential genotypes (GII.4, GII.g/GII.12, GII.17). The potential recombinant GII.g/GII.12-like genotypes were dominant (4/7), with 95.0 to 100 % nucleotide sequence identity among themselves. They were highly related (95.2 to 96.5 % sequence identity) to human NoV GII.g/GII.12 isolates from Sydney, Australia (Gunnedah/ NSW895P/2010/AU: JQ613569 and Wahroonga/NSW004P/2009/ AU: JQ613568) and China (F106/Beijing/2009: JQ889812 and F106/Beijing/2009: JQ899442) (Fig. 1). Phylogenetic analysis and BLAST search also demonstrated that one strain, identified as L6-ETH-2013 clustered with the GII.17 strain from Japan (Kawasaki/2014/JP: AB983218), with the highest nucleotide sequence similarity (95.4 % identity). The only GII.4 isolate sequenced in the RNA polymerase region, AH12-ETH-2013, shared 98.3 % nucleotide sequence identity with a strain isolated in Italy (113196-4/2014/Italy: KJ598058). The other strain identified, AH35-ETH-2013, clustered with a separate genogroup (GIV), displaying high nucleotide sequence similarity (94.2-95.0 % identity) to several strains from China (CCDCGR1113-59/CHN/2011:KC894731), Australia (LakeMacquarie /NSW268O /2010/AU: JQ613567) and Spain (NV2942/VLC/Spain/2012: KC119502) (Fig. 1).

Neighbor-joining phylogenetic tree based on a partial 300-nucleotide sequence of the RdRP gene of human norovirus. Only bootstrap values greater than 70 % are shown. The prefix ZS and boldface type indicate samples from the current study. GenBank accession numbers are shown in parentheses
Sequence and phylogenetic analysis of a portion of the capsid region
Of the 47 NoV positives amplified using primers designed based on a portion of the capsid region, we sequenced 15 samples (Table 5), and a BLAST search confirmed that the sequenced strains were all human NoV GI and GII strains (ZS-L5-ETH-2013, ZS-5152-ETH-2013, ZS-L38-ETH-2013, ZS-5687-ETH-2013, ZS-KH07-2013-ETH, ZS-L58-ETH-2013, ZS-L80-ETH-2013, ZS-L87-ETH-2013, ZS-AH77-ETH-2013, ZS-KH17-ETH-2013, ZS-50A-ETH-2013, ZS-5847-ETH-2013, ZS-5688-ETH-2013, ZS-5595-ETH-2013, ZS-5379-ETH-2013). We submitted these NoV sequences of the partial capsid region to the GenBank database (KM589642, KM589643, KM589644, KM589645, KM589646, KM589647, KM589648, KM589649, KM589650, KM589651, KM589652, KM589654, KU705716, KU705717, and KU705718, respectively). A neighbor-joining tree indicated that the 15 NoV strains from this study were diverse and belonged to the GI and GII types. The majority of the NoVs identified were GII (11/15:73.3 %), which occurs in the majority of NoV infections worldwide, while four strains (26.7 %) belonged to GI. The GII strains identified were further divided into two different genotypes (GII.4 and GII.17) (Fig. 2). Of the GII isolates analyzed in this study (L5, L38, L80, L58, KH07, 5688, 5152, 5379, 5687, 5847, 5595), the majority (63.6 %, 7/11) belonged to GII.17 (L80, L58, KH07, 5688, 5152, 5687, 5847), and four belonged to GII.4 (L5, L38, 5379, 5595). Six of the seven GII.17 strains (5152, 5687, 5847, L80, L-58, KH-07) identified shared 93-98 % nucleotide sequence similarity among themselves; however, strain 5688-ETH-2013 shared lower nucleotide sequence similarity (66-70 % identity) with the strains identified from this study as well as with previously published strains. The other strains that clustered as GII-17 shared high nucleotide sequence similarity (90-95 % identity) with the prototype strains from the USA (CS-E1/2002/USA: AY502009), French Guiana (C142/GF/1978: JN699043 and C142/1978/GUF: KC597139), Ethiopia (ETH176/2009/Ethiopia: JF909057) and Japan (JP/2002/Saitama/T87: KJ196286) (Fig. 2).

Neighbor-joining phylogenetic tree based on the 300-nucleotide capsid gene of human NoV. Only bootstrap values greater than 70 % are shown. The prefix ZS and boldface type indicate samples from the current study. GenBank accession numbers are shown in parentheses
The phylogenetic analysis and BLAST search also demonstrated that our strains L5-ETH-2013 and L38-ETH-2013 belong to GII.4. The two GII.4 strains from this study clustered with the newly emergent variant Sydney 2012, but with different levels of nucleotide sequence similarity. Strain 5379 shared 72 % sequence identity with strains from Australia (Sydney/NSW0514/2012/AU:JX459908, 35408489/AUS/2012: KF177448, 30949403/ AUS/2012: KF177442), while strain 5595 shared 76 % sequence identity with strains from Sydney (285/2010/BFA: JX416414, Sydney2012/FRA :KF008243, Sydney2012/FRA:KF008242 and NSW369N/2012/AU:KF060041). These two strains may possibly be new genotypes, pending further analysis. Strains L-38 and L-5 shared high nucleotide sequence similarity (98 and 99 % identity), with strains from Australia (Sydney 2012/FRA:KF008240, Sydney 2012/FRA: KF008242, Sydney 2012/FRA: KF008243).
Primer pair G1SKF/G1SKR identified 13 samples, of which four were sequenced (Table 5) and grouped as GI (50A [KM589652], KH17-ETH-2013 [KM589651], L87-ETH-2013 [KM589649], and AH77-ETH-2013 [KM589650]) sharing 77 to 100 % nucleotide sequence identity among themselves. Upon analysis, of these four GI strains, two clustered together, L87-ETH-2013, AH77-ETH-2013, and shared high (99 %) nucleotide sequence identity with one another and 95-97 % identity with GI.5 strains from South Africa (10-07-05-K/2010/ZAF: JN191360), Hungary (Siklos-HUN/2013: KJ402295) and Russia (2003/3168/ Moscow/ RUS: FJ383818). They also shared 91 % nucleotide sequence identity with the prototype strain, MUSGROV-GBR00 (AJ277614). The other strain, KH17-ETH-2013, shared high nucleotide sequence similarity (94.3 % identity) with a strain isolated from Russia (2006/Nizhny Novgorod/RUS: FJ383816) and 80 % and 81 % identity with the prototype GI.7 (Winchester/94/UK: AJ277609) and GI.8 strains (NLV/Boxer/2001/US: AF538679), respectively. Strain 50A-2013-ETH shared very high similarity (99.7 % identity) with GI.4 strains from Taiwan (GQ401129-GI-BFDA-07/2009/TAW and GQ401127-/GI/BFDA-05/2009/TAW) and 94.2 % identity with the prototype AB042808-G1.4-CHIBA-JPN00 from Japan (Fig. 2).
Genetic and phylogenetic analysis of sapoviruses
Sapoviruses are currently divided into five genogroups, although additional potential genogroups have also been proposed [17–19]. In the present study, a total of nine SaVs were detected using the genogroup-specific primer pair PEC66/PEC65, which is often used to detect porcine SaV GIII. Of the nine positive SaV samples detected, one from a 2-year-old child that contained adequate RNA levels was sequenced (ZS-22SA-2013-ETH) (Table 5). A BLAST search confirmed that the sequenced strain was a human SaV GII.2 strain with 91 to 94 % sequence identity to strains isolated in Brazil (VIG-AM-116006/BRA: KF924392 and NSC066/PA/BRA/1993: EU295952) and in Malawi (MW514: AB234230) (Fig. 3). We submitted this partial SaV RdRp sequence to the GenBank database (accession number: KM589661). A phylogenetic tree was generated using SaV sequences from humans and swine, based on the published sequences of SaV reference and representative strains from NCBI (Fig. 3).

Neighbor-joining phylogenetic tree based on the 300-nucleotide capsid gene of human SaV. Only bootstrap values greater than 80 % are shown. The prefix ZS and boldface type indicate samples from the current study. GenBank accession numbers are shown in parentheses
Discussion
Prevalence and genogrouping/genotyping of noroviruses and sapoviruses
The results from this study, based on direct molecular detection and characterization, confirmed the presence of NoVs (25.3 %) and SaVs (4.2 %) in diarrheic patients in all age groups. Recent studies have shown that NoV plays an important etiological role in sporadic diarrhea in infants and young children [46, 47]. In our study also, we observed that the rate of NoV infection was significantly higher (p = 0.007) in children <5 years old than in older children and adults. This is in agreement with other study that reported the high prevalence of this pathogen in children <5 years of age [48]. However, unlike several studies conducted in Africa and elsewhere [49], we found NoV-associated diarrhea occurring mainly among children 1-4 years old, more frequently than among infants <1 year old. This could be related to the small number of samples from infants <1 year old in the present study. However, similar findings were reported from industrialized countries, including England and the Netherlands, with the prevalence of NoV infection peaking in children 1–4 years old, indicating acquisition of NoV at a later age or widespread protection by maternal antibodies in infants <1 year old [50–52].
Genogrouping/genotyping of human enteric viruses
Among the 54 NoV-positive samples, sufficient RNA to enable genotype determination by sequence analysis was obtained from 22 samples (22/54: 40.7 %). Low viral load or unstable RNA in the samples might account for failure in genotyping of some of the viruses. Although NoVs could only be genotyped in 40.7 % of positive samples, the observed genetic diversity was very high. Multiple genogroups (all of the genogroups classified as human NoVs, GI, GII and GIV) were detected from all age groups, with different frequencies. Norovirus GII was the dominant circulating genogroup detected (n = 17: 77.2 %). This finding is consistent with most studies worldwide, including those from Africa, in which NoV GII was reported to be predominantly responsible for gastroenteritis outbreaks and sporadic cases [3, 6, 21, 22, 29, 49, 53, 54]. Nucleotide sequence and phylogenetic analysis of the NoV GII strains revealed that GII.17 NoVs comprise the predominant genotype (36.4 %) detected in this study. This is in contrast to most studies in Africa, as well as in Europe and the USA, where GII.4 is the dominant genotype. However, similar to this study, genotype GII.17 has been identified as the dominant genotype in water in Kenya [55]. It might not be surprising to detect similar newly detected unusual strains dominating in both countries, as the countries share borders and cross-country rivers. Moreover, some previous studies have suggested that non-GII.4 NoVs have been predominant in the past, worldwide [56]. For example, analysis of archived samples from 1974 through 1991 has shown that the frequency of GII.3 was 48 %, compared with 16 % for GII.4 and 14 % for GII.7 strains [57]. Therefore, it is essential to study increases in non-GII.4 strains to determine possible reasons for their increased transmissibility or population susceptibility. GII.17 strains were also reported in patients with acute gastroenteritis in other countries: Brazil, Paraguay, Argentina, South Korea, Thailand, Kenya, the USA and Japan [23, 58–62]. Although not dominant, GII.17 was also detected from clinical specimens in some African countries, including Djibouti [53], Cameroon [49], Morocco [63] and Burkina Faso [64]. Norovirus GII.17 is of clinical relevance, as it has been implicated in nosocomial NoV infection [65] and chronic NoV infection in a kidney transplant patient [66].
Strains homologous to GII.4 NoV strains that are associated with large epidemics of NoV worldwide were also detected in this study. Several NoV pandemics associated with GII.4 have occurred every 2 to 3 years [67–74]. The recent or new pandemic isolate GII.4 2012 Sydney, which was first identified in March 2012 in Australia [75], is currently the dominant GII.4 variant identified in NoV-associated gastroenteritis outbreaks worldwide [75]. The GII.4 Sydney 2012 variant from Australia accounted for most of the GII.4 isolates identified in this study. The sample collection for the current study was conducted during June to September 2013 (Table 4), the time when this new variant was spreading worldwide. This shows the global distribution of this GII.4-Sydney variant. While Australia is geographically isolated in the world, phylogenetic analysis revealed that the same strains are circulating in Asia, Europe, and the Americas. Modern methods of transport and population movement are facilitating the importation of global NoV strains [76]. This study suggests that the majority of the NoV GII outbreak strains isolated in Australia mirror the strains causing outbreaks during similar time frames in Africa too, which demonstrates the ability of these viruses to spread globally very rapidly. Additionally, two of the GII.4 strains identified in this study (5595-ETH, 5379-ETH) were about 75 % identical over 282 nucleotides to GII.4 variant Sydney 2012. According to Zheng et al. [12], to be classified into a genotype, strains are expected to have 80 % nucleotide sequence identity in the capsid (VP1) region. Therefore, this finding indicates that these two viruses, which were characterized as GII.4 NoV using region B primers, may represent novel NoV types, with gene sequences slightly different from the previously recognized human NoVs.
Along with mutations, recombination may also contribute to the genetic variability and evolution of NoVs. The four potential intergenotypic recombinant GII.g/GII.12-like strains from this study were highly related to human NoV recombinants from Sydney. Recombination, which has already been described for NoVs, is an important mechanism in the evolution of RNA viruses, since it can create changes in virus genomes by exchanging sequences and producing new viruses. It commonly occurs at the ORF1-ORF2 junction [77], although other recombination sites, such as the one between the ORF2 and ORF3 regions, have been reported [78, 79]. Recently there has been a significant increase in the number of NoV recombinants that have been detected. Norovirus recombinant GII.g/GII.12, emerged and was first detected in sporadic cases in Australia in 2008 [79, 80] and caused an outbreak in New Zealand in the same year. Ever since, the GII.g/GII.12 recombinant has been reported in several countries, including the United States, where it was responsible for 16 % of gastroenteritis outbreaks in the 2009–2010 winter season [81], and also in Belgium [82], Korea [83], Italy [84], and recently, China [85]. Following the original observation of Yasin et al. (2011) in Ethiopia, ours is the first report of potential GII.g/GII.12-like recombinants in Ethiopia, although, we need to verify more, as recombination may create chimeric strains bearing ORF1 and ORF2 of different parental origin, thus requiring multiple target (ORF1- and ORF2-based) analysis to more accurately characterize the NoV strains [86]. This study also illustrates that recombinants may be more prevalent than was originally thought, which might result in generation of new viruses with unknown pathogenic potential, virulence characteristics and altered species tropism for both animals and humans [87]. In addition, recombination of NoV strains could lead to confusion of the classification system for the virus by affecting phylogenetic grouping, since NoVs are routinely genotyped by analysis of partial capsid or polymerase gene sequences. This will, in turn, affect molecular epidemiological studies, which could ultimately have major implications for vaccine design. Therefore, to monitor the prevalence and emergence of such new strains of public-health importance, a dual nomenclature using both ORF1 and VP1 sequences could be relevant, as proposed by Kroneman et al. [14].
In the present study, GI NoVs, in particular, GI.4, GI.5 and GI.8, were detected and characterized. GI has been described as a genogroup that circulates to a lesser extent than GII [88], but GI viruses have been implicated more often than GII viruses in waterborne outbreaks [89]. This might be because they are more stable in the environment. In Cameroon, GI NoV has been detected in wastewater at a low but stable concentration all year around [90]. Most GI outbreaks worldwide were caused by three genotypes, including GI.4 [89]. In a study in Spain from 2007 to 2009, GI.4 was one of the most abundant genotypes identified in sewage and river water [89]. These findings might reflect better survival of GI.4 in the environment compared to other NoV strains. Furthermore, GI.4 also has been associated with foodborne NoV outbreaks [89].
Although NoVs are a common cause of acute gastroenteritis worldwide, GIV NoV strains related to illness in humans are rarely reported [54, 91–93]. GIV strains have been commonly associated with infections in animals, including dogs, cats and lions [94, 95]. GIV.1 genotypes, classified as Alphatron strain in most typing schemes [11, 12] and as a GII.17 strain in another paper [96], are human strains, while GIV.2 NoV has been designated as a canine NoV. Little is known about the origins of the GIV viruses. However, it has been suggested that GIV strains may have arisen in humans from interspecies transmission [97]. The oldest known GIV.1 strain was detected in archived stool obtained from a child in Entebbe, Uganda, in 1976 (Hu/NoV/E22/Uganda/1976/ GIV.1) [98]. Few partial sequences for the human GIV.1 viruses are available in GenBank [99]. Here, we report a potential human GIV genotype 1 (GIV.1) NoV strain (AH35-2013-ETH: KM589661) identified from a diarrheic child, with high sequence similarity to the recently reported NoV GIV.1 strain from Sydney, China and Spain.
There is scarcity of molecular data on SaVs in Africa. In the present study, SaV was detected and sequenced. A BLAST search confirmed that the sequenced strain was of human SaV GII.2, which was detected using the genogroup-specific primer pair PEC66/PEC65 for GIII SaVs. This is probably due to the low sensitivity of the current primer set PEC66/PEC65, which was designed based on the Po/SaV/GIII/Cowden strain detected in 1980 [100]. The predominant human SaV detected in clinical samples in most countries, including African countries, is GI.1, followed by GI.2. In Africa, GII.2 was detected in Kenya [23], South Africa [15] and Malawi [25]. This is the first detection of human SaVs from Ethiopia.
In conclusion, the present study confirms the significant role of NoV infection in diarrheic patients and provides evidence for the genetic diversity of NoVs in Ethiopia. We detected an undefined NoV genotype, potential recombinant-like strains, newly emerging variants, and rarely detected human NoV strains. This indicated that the emerging and undefined NoV genotypes have importance in Africa, including Ethiopia. Moreover, the detection of multiple genotypes of NoVs other than GII.4, which is the dominant genotype associated with NoV infection worldwide, is also important, since these strains may be circulating silently in the community, but with high clinical significance. The results presented here also suggest that recombination is not a rare phenomenon among NoVs, which thus contributes to their genetic diversity. Additionally, NoV affects more-vulnerable groups, such as the very young, which has important implications in a country like Ethiopia, where the number of children under five is high. Therefore, it is important to protect these vulnerable groups who are at the highest risk and establish a NoV surveillance reporting system that could help in understanding the public-health impact of diarrheal diseases.
References
Boschi-Pinto C, Velebitb L, Shibuyac K (2008) Estimating child mortality due to diarrhoea in developing countries. Bull WHO 86(9):710–719
Liu L, Johnson HL, Cousens S, Perin J, Scott S, Liu L, Johnson HL, Cousens S, Perin J, Scott S, Lawn JE, Rudan I, Campbell H, Cibulskis R, Li M, Mathers C, Black RE (2012) Global, regional, and national causes of child mortality: an updated systematic analysis for 2010 with time trends since 2000. Lancet 379:2151–2161
Patel MM, Widdowson MA, Glass RI, Akazawa K, Vinje J, Parashar UD (2008) Systematic literature review of role of noroviruses in sporadic gastroenteritis. Emerg Infect Dis 14(8):1224–1231
Glass RI, Parashar UD, Estes MK (2009) Norovirus gastroenteritis. N Engl J Med 361(18):1776–1785
Koo HL, Neill FH, Estes MK, Munoz FM, Cameron A, DuPont HL, Atmar RL (2013) Noroviruses: the most common pediatric viral enteric pathogen at a large university hospital after introduction of rotavirus vaccination. J Pediatric Infect Dis Soc 2(1):57–60
Payne DC, Vinjé J, Szilagyi PG, Edwards KM, Staat MA, Weinberg GA, Hall CB, Chappell J, Bernstein DI, Curns AT, Wikswo M, Shirley SH, Hall AJ, Lopman B, Parashar UD (2013) Norovirus and medically attended gastroenteritis in US children. N Engl J Med 368(12):1121–1130
Hemming M, Räsänen S, Huhti L, Paloniemi M, Salminen M, Vesikari T (2013) Major reduction of rotavirus, but not norovirus, gastroenteritis in children seen in hospital after the introduction of RotaTeq vaccine into the National Immunization Programme in Finland. Eur J Pediatr 172(6):739–746
Bucardo F, Reyes Y, Svensson L, Nordgren J (2014) Predominance of norovirus and sapovirus in Nicaragua after implementation of universal rotavirus vaccination. PLoS One 21;9(5):e98201
Clarke IN, Lambden PR (2000) Organization and expression of calicivirus genes. J Infect Dis 181(Suppl 2):S309–S316
Atmar RL, Estes MK (2001) Diagnosis of non-cultivatable gastroenteritis viruses, the human caliciviruses. Clin Microbiol Rev 14(1):15–37
Green K (2013) Caliciviridae: the noroviruses. In: Knipe DM, Howley PM (eds) Fields virology, vol 1, 6th edn. Lippincott Williams & Wilkins, Philadelphia, pp 583–609
Zheng DP, Ando T, Fankhauser RL, Beard RS, Glass RI, Monroe SS (2006) Norovirus classification and proposed strain nomenclature. Virology 346:312–323
Vega E, Barclay L, Gregoricus N, Shirley SH, Lee D, Vinjé J (2014) Genotypic and epidemiologic trends of norovirus outbreaks in the United States, 2009 to 2013. J Clin Microbiol 52(1):147–155
Kroneman A, Vega E, Vennema H, Vinjé J, White PA, Hansman G, Green K, Martella V, Katayama K, Koopmans M (2013) Proposal for a unified norovirus nomenclature and genotyping. Arch Virol 158(10):2059–2068
Mans J, Murray TY, Taylor MB (2014) Novel norovirus recombinants detected in South Africa. Virol J 11:168
Farkas T, Zhong WM, Jing Y, Huang PW, Espinosa SM, Martinez N, Morrow AL, Ruiz-Palacios GM, Pickering LK, Jiang X (2004) Genetic diversity among sapoviruses. Arch Virol 149:1309–1323
Wang QH, Souza M, Funk JA, Zhang W, Saif LJ (2006) Prevalence of noroviruses and sapoviruses in swine of various ages determined by reverse transcription-PCR and microwell hybridization assays. J Clin Microbiol 44(6):2057–2062
Sisay Z, Wang QH, Oka T, Saif LJ (2013) Prevalence and molecular characterization of porcine enteric caliciviruses and first detection of porcine kobuviruses in US swine. Arch Virol 158(7):1583–1588
Scheuer AK, Oka T, Hoet AE, Gebreyes WA, Molla BZ, Saif LJ, Wang QH (2013) Prevalence of porcine noroviruses, molecular characterization of emerging porcine sapoviruses from finisher swine in the US, and unified classification scheme for sapoviruses. J Clin Microbiol 51(7):2344–2353
Sdiri-Loulizi K, Hassine M, Gharbi-Khelifi H, Aouni Z, Chouchane S, Sakly N, Pothier P, Ambert-Balay K, Aouni M (2011) Molecular detection of genogroup I sapovirus in Tunisian children suffering from acute gastroenteritis. Virus Genes 43:6–12
Kamel AH, Ali MA, El-Nady HG, de Rougemont A, Pothier P, Belliot G (2009) Predominance and circulation of enteric viruses in the region of Greater Cairo, Egypt. J Clin Microbiol 47(4):1037–1045
Kiulia NM, Netshikweta R, Page NA, Van Zyl WB, Kiraithe MM, Nyachieo A, Mwenda JM, Taylor MB (2010) The detection of enteric viruses in selected urban and rural river water and sewage in Kenya, with special reference to rotaviruses. J Appl Microbiol 109(3):818–828
Mans J, Murray TY, Kiulia NM, Mwenda JM, Musoke RN, Taylor MB (2014) Human caliciviruses detected in HIV-seropositive children in Kenya. J Med Virol 86:75–81
Mattison K, Sebunya TK, Shukla A, Noliwe LN, Bidawid S (2010) Molecular detection and characterization of noroviruses from children in Botswana. J Med Virol 82(2):321–324
Page N, Esona M, Seheri M (2010) Characterization of Genotype G8 Strains From Malawi, Kenya, and South Africa. J Med Virol 82:2073–2081
Ayukekbong J, Andersson ME, Vansarla G, Tah F, Nkuo-Akenji T, Lindh M, Bergström T (2014) Monitoring of seasonality of norovirus and other enteric viruses in Cameroon by real-time PCR; an exploratory study. Epidemiol Infect 142(7):1392–1402
Kabue JP, Meader E, Hunter PR, Potgieter N (2016) Human Norovirus prevalence in Africa: a review of studies from 1990 to 2013. Trop Med Int Health 21(1):2–17
Munjita SM (2015) Current status of norovirus infections in children in Sub-Saharan Africa. J Trop Med 2015:309648
Mans J, Murray TY, Nadan S, Netshikweta R, Page NA, Taylor MB (2015) Norovirus diversity in children with gastroenteritis in South Africa from 2009 to 2013: GII.4 variants and recombinant strains predominate. Epidemiol Infect 16:1–10 [Epub ahead of print]
Mans J, Murray TY, Taylor MB (2014) Novel norovirus recombinants detected in South Africa. Virol J 17;11:168
Kabayiza JC, Andersson ME, Nilsson S, Bergström T, Muhirwa G, Lindh M (2014) Real-time PCR identification of agents causing diarrhea in Rwandan children less than 5 years of age. Pediatr Infect Dis J 33(10):1037–1042
Elfving K, Andersson M, Msellem MI, Welinder-Olsson C, Petzold M, Björkman A, Trollfors B, Mårtensson A, Lindh M (2014) Real-time PCR threshold cycle cutoffs help to identify agents causing acute childhood diarrhea in Zanzibar. J Clin Microbiol 52(3):916–923
Murray TY, Nadan S, Page NA, Taylor MB (2016) Diverse sapovirus genotypes identified in children hospitalised with gastroenteritis in selected regions of South Africa. J Clin Virol 76:24–29
Murray TY, Taylor MB (2015) Quantification and molecular characterisation of human sapoviruses in water sources impacted by highly polluted discharged wastewater in South Africa. J Water Health 13(4):1055–1059
Matussek A, Dienus O, Djeneba O, Simpore J, Nitiema L, Nordgren J (2015) Molecular characterization and genetic susceptibility of sapovirus in children with diarrhea in Burkina Faso. Infect Genet Evol 32:396–400
Benabbes L, Ollivier J, Schaeffer J, Parnaudeau S, Rhaissi H, Nourlil J, Le Guyader FS (2013) Norovirus and other human enteric viruses in moroccan shellfish. Food Environ Virol 5(1):35–40
Yassin MA, Kirby A, Mengistu AA, Arbide I, Dove W, Beyer M, Cunliffe NA, Cueva LE (2012) Unusual norovirus and rotavirus genotypes in Ethiopia. Paed Int Child Health 32(1):51–55
Central Statistical Agency (2007) Population and Housing Census Report-Addis Ababa Region—2007, 2010–07. http://www.csa.gov.et/index.php/2013-02-20-14-51-51/…/census-2007
Nakamura K, Saga Y, Iwai M, Obara M, Horimoto E, Hasegawa S, Kurata T, Okumura H, Nagoshi M, Takizawa T (2010) Frequent detection of noroviruses and sapoviruses in swine and high genetic diversity of porcine sapovirus in Japan during fiscal year 2008. J Clin Microbiol 48:1215–1222
Jiang X, Huang PW, Zhong WM, Farkas T, Cubitt DW, Matson DO (1999) Design and evaluation of a primer pair that detects both Norwalk- and Sapporo-like caliciviruses by RT-PCR. J Virol Methods 83:145–154
Le Guyader F, Neill FH, Estes MK, Monroe SS, Ando T, Atmar RL (1996) Detection and analysis of a small round-structured virus strain in oysters implicated in an outbreak of acute gastroenteritis. Appl Environ Microbiol 62(11):4268–4272
Kojima S, Kageyama T, Fukushi S, Hoshino FB, Shinohara M, Uchida K, Natori K, Takeda N, Katayama K (2002) Genogroup-specific PCR primers for detection of Norwalk-like viruses. J Virol Methods 100:107–114
Guo M, Hayes J, Cho KO, Parwani AV, Lucas LM, Saif LJ (2001) Comparative pathogenesis of tissue culture-adapted and wild-type Cowden porcine enteric calicivirus (PEC) in gnotobiotic pigs and induction of diarrhea by intravenous inoculation of wild-type PEC. J Virol 75:9239–9251
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729
Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Patel MM, Widdowson MA, Glass RI, Akazawa K, Vinjé J, Parashar UD (2008) Systematic literature review of role of noroviruses in sporadic gastroenteritis. Emerg Infect Dis 14:1224–1231
Ramani S, Kang G (2009) Viruses causing childhood diarrhoea in the developing world. Curr Opin Infect Dis 22:477–482
Martin CCW, Leung TF, Chung TWS, Kwok AK, Nelson EAS, Lee N, Chan PKS (2015) Virus genotype distribution and virus burden in children and adults hospitalized for norovirus gastroenteritis, 2012–2014, Hong Kong. Sci Rep 5:11507
Ayukekbong J, Lindh M, Nenonen N, Tah F, Nkuo-Akenji T, Bergstro MT (2011) Enteric viruses in healthy children in Cameroon: viral load and genotyping of norovirus strains. J Med Virol 83:2135–2142
de Wit MA, Koopmans MP, Kortbeek LM, van Leeuwen NJ, Bartelds A, van Duynhoven YT (2001) Gastroenteritis in sentinel general practices, The Netherlands. Emerg Infect Dis 7:82–91
Iturriza Gomara M, Simpson R, Perault AM, Redpath C, Lorgelly P, Joshi D, Mugford M, Hughes CA, Dalrymple J, Desselberger U (2008) Structured surveillance of infantile gastroenteritis in East Anglia, UK: incidence of infection with common viral gastroenteric pathogens. Epidemiol Infect 136(1):23–33
Gray JJ, Jiang X, Morgan-Capner P, Desselberger U, Estes MK (1993) Prevalence of antibodies to Norwalk virus in England: detection by enzyme-linked immunosorbent assay using baculovirus-expressed Norwalk virus capsid antigen. J Clin Microbiol 31(4):1022–1025
Maslin J, Nicand E, Ambert-Balay K, Fouet C, Kaplon J, Haus R, Pothier P, Kohli E (2008) Detection and characterization of Human caliciviruses associated with sporadic acute diarrhea in adults in Djibouti (horn of Africa). Am J Trop Med Hyg 78(3):522–526
Fankhauser RL, Monroe SS, Noel JS, Ando TA, Glass RI (2002) Epidemiologic and molecular trends of Norwalk-like viruses associated with outbreaks of gastroenteritis in the United States. J Inf Dis 186:1–7
Kiulia N, Mans MJ, Mwenda JM, Taylor MB (2014) Norovirus GII.17 predominates in selected surface water sources in Kenya. Food Environ Virol 6:221–231
Bok K, Abente EJ, Realpe-Quintero M, Mitra T, Sosnovtsev SV, Kapikian AZ, Green KY (2009) Evolutionary dynamics of GII.4 noroviruses over a 34-year period. J Virol 83(22):11890–11901
Rackoff LA, Bok K, Green KY, Kapikian AZ (2013) Epidemiology and evolution of rotaviruses and noroviruses from an archival WHO global study in children (1976–79) with implications for vaccine design. PLoS One 8(3):e59394
Gomes KA, Stupka JA, Diana A, Parra GI (2008) Molecular characterization of calicivirus strains detected in outbreaks of gastroenteritis occurring in Argentina during 2005 and 2006. Revista Argentina de Microbiologia 40:222–228
Ferreira MS, Xavier MdaP, Tinga AC, Rose TL, Fumian TM, Fialho AM (2012) Assessment of gastroenteric viruses frequency in a children’s day care center in Rio De Janeiro, Brazil: a fifteen year study (1994–2008). PLoS One 7(3):e33754
Park SH, Kim EJ, Oh SA, Kim CK, Choi SS, Cho SJ (2011) Viral agents associated with acute gastroenteritis in Seoul, Korea. Clin Lab 57:59–65
Kittigul L, Pombubpa K, Taweekate Y, Diraphat P, Sujirarat D, Khamrin P (2010) Norovirus GII-4 2006b variant circulating in patients with acute gastroenteritis in Thailand during a 2006–2007 study. J Med Virol 82:854–860
Chung JY, Han TH, Park SH, Kim SW, Hwang ES (2010) Detection of GII-4/2006b variant and recombinant noroviruses in children with acute gastroenteritis, South Korea. J Med Virol 82:146–152
El Qazoui M, Oumzil H, Baassi L, El Omari N, Sadki K, Amzazi S, Benhafid M, El Aouad R (2014) Rotavirus and Norovirus infections among acute gastroenteritis children in Morocco. BMC Infect Dis 14:300
Nordgren J, Nitiema LW, Ouermi D, Simpore J, Svensson L (2013) Host genetic factors affect susceptibility to norovirus infections in Burkina Faso. PLoS One 8:e69557
Sukhrie FH, Beersma MF, Wong A, van der Veer B, Vennema H, Bogerman J (2011) Using molecular epidemiology to trace transmission of nosocomial norovirus infection. J Clin Microbiol 49:602–606
Schorn R, Höhne M, Meerbach A, Bossart W, Wüthrich RP, Schreier E, Nicolas JM, Fehr T (2010) Chronic norovirus infection after kidney transplantation: molecular evidence for immune driven viral evolution. Clin Infect Dis 51:307–314
Zheng DP, Widdowson MA, Glass RI, Vinjé J (2010) Molecular epidemiology of genogroup II-genotype 4 noroviruses in the United States between 1994 and 2006. J Clin Microbiol 48:168–177
Lindesmith LC, Donaldson EF, Beltramello M, Pintus S, Corti D, Swanstrom J, Debbink K, Jones TA, Lanzavecchia A, Baric RS (2014) Particle conformation regulates antibody access to a conserved GII.4 norovirus blockade epitope. J Virol 88(16):8826–8842
Debbink K, Lindesmith LC, Ferris MT, Swanstrom J, Beltramello M, Corti D, Lanzavecchia A, Baric RS (2014) Within-host evolution results in antigenically distinct GII.4 noroviruses. J Virol 88(13):7244–7255
Debbink K, Lindesmith LC, Donaldson EF, Costantini V, Beltramello M, Corti D, Swanstrom J, Lanzavecchia A, Vinjé J, Baric RS (2013) Emergence of new pandemic GII.4 Sydney norovirus strain correlates with escape from herd immunity. J Infect Dis 208(11):1877–1887
Lindesmith LC, Costantini V, Swanstrom J, Debbink K, Donaldson EF, Vinjé J, Baric RS (2013) Emergence of a norovirus GII.4 strain correlates with changes in evolving blockade epitopes. J Virol 87(5):2803–2813
Lindesmith LC, Beltramello M, Donaldson EF, Corti D, Swanstrom J, Debbink K, Lanzavecchia A, Baric RS (2012) Immunogenetic mechanisms driving norovirus GII.4 antigenic variation. PLoS Pathog 8(5):e1002705
Debbink K, Donaldson EF, Lindesmith LC, Baric RS (2012) Genetic mapping of a highly variable norovirus GII.4 blockade epitope: potential role in escape from human herd immunity. J Virol 86(2):1214–1226
Lindesmith LC, Donaldson EF, Baric RS (2011) Norovirus GII.4 strain antigenic variation. J Virol 85(1):231–242
Eden JS, Hewitt J, Lim KL, Boni MF, Merif J, Greening G, Ratcliff RM, Holmes EC, Tanaka MM, Rawlinson WD, White PA (2014) The emergence and evolution of the novel epidemic norovirus GII.4 variant Sydney 2012. Virology 450–451:106–113
Widdowson MA, Sulka A, Bulens SN, Beard RS, Chaves SS, Hammond R, Swanson E, Totaro J, Woron R, Mead PS, Bresee JS, Monroe SS, Glass RI (2005) Norovirus and foodborne disease, US, 1991–2000. Emerg Infect Dis 11:95–102
Bull RA, Tanaka M, White PA (2007) Norovirus recombination. J Gen Virol 88:3347–3359
Chabra P, Walimbe AM, Chitambar SD (2010) Complete genome characterization of Genogroup II norovirus strains from India: evidence of recombination in ORF2/3 overlap. Infect Genet Evol 10:1101–1109
Eden JS, Tanaka MM, Boni MF, Rawlinson WD, White PA (2013) Recombination within the pandemic norovirus GII.4 lineage. J Virol 87:6270–6282
Eden JS, Bull RA, Tu E, McIver CJ, Lyon MJ, Marshall JA, Smith DW, Musto J, Rawlinson WD, White PA (2010) Norovirus GII.4 variant 2006b caused epidemics of acute gastroenteritis in Australia during 2007 and 2008. J Clin Virol 49:265–271
Vega E, Vinjé J (2011) Novel GII.12 norovirus strain, US, 2009–2010. Emerg Infect Dis 17:1516–1518
Mathijs E, Denayer S, Palmeira L, Botteldoorn N, Scipioni A, Vanderplasschen A, Thiry E, Dierick K (2011) Novel norovirus recombinants and of GII.4 sub-lineages associated with outbreaks between 2006 and 2010 in Belgium. Virol J 18(8):310
Han TH, Kim CH, Chung JY, Park SH, Hwang ES (2011) Emergence of norovirus GII-4/2008 variant and recombinant strains in Seoul, Korea. Arch Virol 156(2):323–329
Giammanco GM, Rotolo V, Medici MC, Tummolo F, Bonura F, Chezzi C, Martella V, De Grazia S (2012) Recombinant norovirus GII.g/GII.12 gastroenteritis in children. Infect Genet Evol 12:169–174
Sang S, Zhao Z, Suo J, Jia N, Gao Y, Xie L, Du M, Liu B, Ren S, Liu Y (2014) Report of recombinant Norovirus GII.g/GII.12 in Beijing, China. PLoS One 9(2):e88210
Ambert-Balay K, Bon F, Le Guyader F, Pothier P, Kohli E (2005) Characterization of new recombinant noroviruses. J Clin Microbiol 43:5179–5186
Hansman GS, Oka T, Katayama K (2007) Human sapoviruses: genetic diversity, recombination, and classification. Rev Med Virol 17:133–141
Barreira DM, Ferreira MS, Fumian TM, Checon R, Sadovsky AD, Leite JP, Miagostovich MP, Spano LC (2010) Viral load and genotypes of noroviruses in symptomatic and asymptomatic children in southeastern Brazil. J Clin Virol 47:60–64
Gim TAW, Gin Karina YH, Lin L, Oon E, Chen EX, Woo CH (2009) Prevalence and genotypes of human noroviruses in tropical urban surface waters and clinical samples in Singapore. Appl Environ Microbiol 75(15):4984–4992
Nordgren J, Matussek A, Mattsson A, Svensson L, Lindgren PE (2009) Prevalence of norovirus and factors influencing virus concentrations during one year in a full-scale wastewater treatment plant. Water Res 43:1117–1125
Hall AJ, Curns AT, McDonald LC, Parashar UD, Lopman BA (2012) The roles of Clostridium difficile and norovirus among gastroenteritis associated deaths in the US, 1999–2007. Clin Infect Dis 55:216–223
Iritani N, Seto Y, Kubo H, Haruki K, Ayata M, Ogura H (2002) Prevalence of “Norwalk-like virus” infections in outbreaks of acute nonbacterial gastroenteritis observed during the 1999–2000 season in Osaka City, Japan. J Med Virol 66(1):131–138
La Rosa G, Pourshaban M, Iaconelli M, Muscillo M (2008) Detection of genogroup IV noroviruses in environmental and clinical samples and partial sequencing through rapid amplification of cDNA ends. Arch Virol 153:2077–2083
Martella V, Banyai K, Matthijnssens J, Buonavoglia C, Ciarlet M (2010) Zoonotic aspects of rotaviruses. Vet Microbiol 140:246–255
Martella V, Lorusso E, Decaro N, Elia G, Radogna A, D’Abramo M, Desario C, Cavalli A, Corrente M, Camero M, Germinario CA, Bányai K, Di Martino B, Marsilio F, Carmichael LE, Buonavoglia C (2008) Detection and molecular characterization of a canine norovirus. Emerg Infect Dis 14:1306–1308
Kageyama T, Shinohara M, Uchida K, Fukushi S, Hoshino FB, Kojima S, Takai R, Oka T, Takeda N, Katayama K (2004) Coexistence of multiple genotypes, including newly identified genotypes, in outbreaks of gastroenteritis due to Norovirus in Japan. J Clin Microbiol 42(7):2988–2995
Siebenga JJ, Vennema H, Zheng DP, Vinje J, Lee BE, Pang XL, Ho EC, Lim W, Choudekar A, Broor S (2009) Norovirus illness is a global problem: emergence and spread of norovirus GII.4 variants, 2001–2007. J Infect Dis 200(5):802–812
Rackoff Lauren A, Bok Karin, Green Kim Y, Kapikian Albert Z (2013) Epidemiology and evolution of rotaviruses and noroviruses from an archival WHO global study in children (1976–79) with implications for vaccine design. PLoS One 8(3):e59394
Martella V, Decaro N, Lorusso E, Radogna A, Moschidou P, Amorisco F, Lucente MS, Desario C, Mari V, Elia G, Banyai K, Carmichael LE, Buonavoglia C (2009) Genetic heterogeneity and recombination in canine noroviruses. J Virol 83:11391–11396
Saif LJ, Bohl EH, Theil KW, Cross RF, House JA (1980) Rotavirus-like, calicivirus-like, and 23-nm virus-like particles associated with diarrhea in young pigs. J Clin Microbiol 12(1):105–111
Acknowledgments
We thank the staffs of the different health centers, especially the laboratory technologists at Lideta Health Center, who helped us a lot in sample collection and filling out the questionnaire. We are very grateful to the patients who participated in this project. We highly acknowledge the help of Dr. Girmay Medhin in the statistical analysis. We appreciate the support of the Segolip unit at the BecA-ILRI Hub in sequencing the samples from this study. We are also grateful for the cooperation and support of the Department of Molecular, Cellular and Biological Sciences and Aklilu Lemma Institute of Pathobiology, Addis Ababa University, in facilitating the successful accomplishment of the project. We greatly acknowledge the financial support of the African Biosciences Challenge Fund (ABCF)/ Biosciences of east and central Africa (BecA). We also thank the VPH-Biotech East Africa Consortium and the National Institutes of Health-Fogarty (Fogarty Grant D43TW008650, W Gebreyes, PI, The Ohio State University) for their financial support.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Funding
This study was funded by African Biosciences Challenge Fund (ABCF) Program and VPH-Biotech East Africa Consortium and the National Institutes of Health-Fogarty (Fogarty Grant D43TW008650, W Gebreyes, PI, The Ohio State University). The ABCF program is funded by the Australian Department for Foreign Affairs (DFAT) through the BecA-CSIRO partnership; the Syngenta Foundation for Sustainable Agriculture (SFSA); the Bill & Melinda Gates Foundation (BMGF); the UK Department for International Development (DFID) and; the Swedish International Development Cooperation Agency (Sida).
Conflict of interest
None.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Rights and permissions
About this article

Cite this article
Sisay, Z., Djikeng, A., Berhe, N. et al. Prevalence and molecular characterization of human noroviruses and sapoviruses in Ethiopia. Arch Virol 161, 2169–2182 (2016). https://doi.org/10.1007/s00705-016-2887-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-016-2887-7