
Abstract
During a survey in June 2011, severe leaf yellow mosaic disease was observed on about 45 % plants of Jatropha curcas growing in the Katerniaghat wildlife sanctuary in India. An association of a begomovirus with disease was detected in 15 out of 20 samples by PCR using begomovirus genus-specific primers and total DNA isolated from symptomatic leaf samples. For identification of the begomovirus, the complete genome was amplified using a Phi-29 DNA-polymerase-based rolling-circle amplification kit and total DNA from five representative samples and then digested with BamHI. The linearized RCA products were cloned and sequenced. Their GenBank accession numbers are JN698954 (SKRK1) and JN135236 (SKRK2). The sequences of the two begomovirus isolates were 97 % identical to each other and no more than 86 % to those of jatropha mosaic India virus (JMIV, HM230683) and other begomoviruses reported worldwide. In phylogenetic analysis, SKRK1 and SKRK2 clustered together and showed distant relationships to jatropha mosaic India virus, Jatropha curcas mosaic virus, Indian cassava mosaic virus, Sri Lankan cassava mosaic virus and other begomoviruses. Based on 86 % sequence identities and distant phylogenetic relationships to JMIV and other begomoviruses and the begomovirus species demarcation criteria of the ICTV (<89 % sequence identity of complete DNA-A genome), the begomovirus isolates associated with leaf yellow mosaic disease of J. curcas were identified as members of a new begomovirus species and provisionally designated as jatropha leaf yellow mosaic Katerniaghat virus (JLYMKV). Agroinfectious clones of the DNA molecule of the begomovirus isolate were also generated, and the fulfillment of Koch’s postulates was demonstrated in J. curcas plants.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Jatropha curcas (family Euphorbiaceae) is a shrub plant that is grown mainly for biodiesel production in India. It survives in various climatic conditions and has the ability to grow in almost any type of soil, especially on drought soil, and produce 30-40 % of its seed weight in biodiesel [1]. Widespread mosaic disease of J. curcas caused by begomoviruses was first investigated in 2004 in southern India (Karnataka) [2, 3] and then in northern India [4, 5]. Since then, many new begomoviruses have been reported on J. curcas from India and other countries [6–10].
Based on earlier reports of begomoviruses on Jatropha and the widespread cultivation of this plant in India, the Jatropha plots at wild life sanctuary, Katerniyaghat, Uttar Pradesh, India were surveyed during June, 2011 for begomovirus-disease-like symptoms. Severe leaf yellow mosaic disease was noted on J. curcas plants with an incidence of about 45 %. The severely infected plants exhibited yellow mosaic accompanied by blistering and curling on leaves and stunting of plants as compared to non-symptomatic ones (Fig. 1a, b). Leaf and stems samples from 20 diseased plants were collected for detection of begomoviruses, and stem cuttings were planted in pots and maintained in a glass house for propagation of virus cultures.

Naturally infected J. curcas plant showing severe yellow mosaic accompanied by blistering and curling symptoms on leaves (a), compared to a non-symptomatic plant (b) in the field. (c) An agroinfiltrated J. curcas plant with mosaic symptoms at 45 dpi. (d) Agarose gel electrophoresis of PCR products obtained from DNA samples of symptomatic J. curcas using DNA-A specific primers PALIv 722/ PALIc 1960 [12] showing bands of the expected size of ~1.2 kb in 15 of 20 samples (c)
Based on typical begomovirus-disease-like yellow mosaic symptoms and the population of whiteflies in the vicinity, begomovirus infection was suspected. Therefore, whitefly transmission was attempted using a method described earlier [11]. The pathogen was successfully transmitted to healthy J. curcas seedlings, which developed similar yellow mosaic symptoms on leaves of all five plants by 45 days post-inoculation (dpi).
For detection of begomovirus, total DNA from 20 randomly selected symptomatic leaf samples and one asymptomatic one was isolated using a DNeasy Plant Mini Kit (QIAGEN GmbH, Hilden, Germany) and subjected to polymerase chain reaction (PCR) in a thermal cycler (SureCycler 8800, Agilent Technology, Selangor, Malaysia) using begomovirus DNA-A- specific primers (PALIv 722/ PALIc 1960) [12] and DNA-B-specific primers [13]. The PCR with primers PALIv722 and PALIc1960 resulted in a ~1.2-kb amplicon in 15 out of 20 samples (Fig. 1d), which suggested an association of a begomovirus with the symptoms in the plants. PCR using DNA-B-specific primers [13] did not result in any amplicon from any of these samples, indicating the absence of a DNA-B component. Therefore, these isolates are monopartite begomovirus.
For molecular identification of the detected begomovirus, its complete genome was amplified from the DNA of five infected jatropha leaf samples (SKRK1-5) amplified by rolling-circle amplification (RCA) using a Phi-DNA-polymerase-based TempliPhi™ DNA amplification kit (GE Healthcare, Buckinghamshire, UK) as per the manufacturer’s instructions, and the RCA products were linearized with BamHI restriction enzyme (Thermo Fisher Scientific Inc., Pittsburgh, USA). Electrophoresis of the linearized RCA products resulted in a ~2.7-kb band from all five DNA samples which were cloned and sequenced. Analysis of the sequence data showed that the sequences of SKRK1 and SKRK3 were identical, and the SKRK2, SKRK4, and SKRK5 isolates were identical to each other. Therefore, sequence data for two isolates were submitted to the GenBank database under the accession numbers JN135236 (SKRK1) and JN698954 (SKRK2).
The sequences of the isolates SKRK1 and SKRK2 were 2744 nucleotides long and possessed six open reading frames, V1-2 in the virion sense and C1-4 in the complementary sense, which is a typical arrangement for monopartite begomovirus. The untranslated intergenic regions were from nucleotides 2602 to 2740 and 1 to 134 and contained the nonanucleotide sequence (TAATATTAC) required for replication of begomovirus (Fig. 2a).

(a) Genomic organization of jatropha leaf yellow mosaic Katerniaghat virus showing the arrangement of six predicted ORFs along with their nucleotide coordinates and putative proteins (a). Arrows indicate the positions and orientations (virion/complementary sense) of putative ORFs. V2, pre-coat protein gene; V1, coat protein gene in the virion sense (clockwise), C5, C3, replication enhancer protein gene; C2, transcription activator protein gene; C1, replication-associated protein gene; C4, C4 protein gene in the complementary sense (anticlockwise). The intergenic region is represented as filled blocks with a hairpin structure at the origin of replication. (b) Phylogenetic tree showing distant relationships of Jatropha leaf yellow mosaic Katerniaghat virus to isolates of JMIV, JCMV, ICMV, SrLCMV, ELCV, pedilanthus leaf curl virus and tomato leaf curl Kerala virus. The distance tree was generated using the neighbour-joining method in MEGA v6.1 [14] with 1,000 bootstrap replicates. Bootstrap values are shown at the nodes of the tree
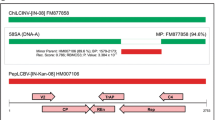
Pairwise comparison of nucleotide sequences showed that the sequences of the begomovirus isolates were 97 % identical to each other and 86 % identical to that of jatropha mosaic India virus isolate JMIV (HM230683) reported in India [8]. SKRK1 and SKRK2 also shared 76-78 % identity with isolates of jatropha curcas mosaic virus (JCMV: JX518289 and GQ924760), 72-77 % identity with isolates of Indian cassava mosaic virus (ICMV: Z24758, AY730035, AJ314739 and AJ575819) and 70-71 % identity with isolates of Sri Lankan cassava mosaic virus (SrLCMV: AJ579307, KC424490, AJ890225 and AJ607394) isolated from J. curcas and Manihot esculenta (Table S1).
For phylogenetic analysis of the begomovirus isolates under study, a tree was constructed using the neighbor-joining method with 1000 bootstrap replicates the MEGA v6.0 [14]. This analysis revealed that SKRK1 and SKRK2 clustered with JMIV isolates and formed a single clade, while JCMV, ICMV, SrLCMV and other begomovirus isolates clustered in a different clade (Fig. 2b). Although the isolates SKRK1 and SKRK2 formed a clade with JMIV, they showed a distant relationship to JCMV, ICMV, SrLCMV and other begomoviruses. The begomovirus isolates under study showed less than 87 % sequence identity and the closest phylogenetic relationship to a JMIV isolate reported from India [8]. The begomovirus species demarcation criterion of the ICTV is 89 % DNA-A sequence identity [15]. The new begomovirus isolates should therefore be considered members of a new species of begomovirus and are provisionally designated as Jatropha leaf yellow mosaic Katerniaghat virus (JLYMKV).
An agroinfectious clone of the DNA molecule of a begomovirus isolate was generated to test the infectivity of the DNA genome. For construction of the agroinfectious clone, the RCA product from the SKRK1 DNA sample was partially digested with BamHI, and a linearized ~5.6-kb band was eluted and ligated into pCAMBIA1300 vector. The resulting dimeric clone was mobilized into Agrobacterium tumefaciens (GV3101), designated as pCAM-JLYMKV-DNA and infiltrated into leaf lamina of J. curcas seedlings at the 4- to 6-leaf stage (3 each). A set of plants were also infiltrated with pCAM-1300 vector alone as a negative control, and all infiltrated plants were kept in an insect-proof chamber and monitored for two months for symptom development. The pCAM-JLYMKV-DNA-infiltrated seedlings of J. curcas (3/3) developed leaf yellow mosaic symptoms at 45 dpi (Fig. 1c) that were similar to those of naturally infected J. curcas plants (from which the virus was cloned), while the plants infiltrated with pCAM-1300 did not develop symptoms. The presence or absence of a DNA genome in the agroinfiltrated plants was tested by PCR using begomovirus-DNA-genome-specific primers [12], and begomovirus DNA was detected in all of the plants infiltrated with pCAM-JLYMKV-DNA. These studies fulfill Koch’s postulates for the DNA genome of the begomovirus isolate from J. curcas plants.
Leaf samples from 57 symptomatic plants were again collected from Katarniaghat for indexing to determine whether they contained JLYMKV. DNA was isolated from all of the samples and dot blotted onto a nylon membrane. The blots were hybridized with a probe prepared from clone of JLYMKV (SKRK1: JN698954) using a method described earlier [16]. Out of 57 tested, 48 showed hybridization with the probe, but no signals were obtained for a healthy J. curcas plant, indicating the presence of JLYMKV in a high percentage, 84.2 % (48/57), of J. curcas plants growing at Katarniyaghat. An uncharacterized disease appeared to be affecting Jatropha simultaneously, as both surveys detected begomovirus in ~85 % of symptomatic plants.
A literature survey revealed many reports of begomoviruses on J. curcas from all over the world, viz. Jatropha mosaic virus [2], Indian cassava mosaic virus [5, 6], African cassava mosaic virus and East African cassava mosaic virus from Kenya [7]; JMIV [8] and JLCrV [10] from India; and jatropha mosaic Nigeria virus from Nigeria [9]. However, a new begomovirus from India, “jatropha leaf yellow mosaic Katerniaghat virus (JLYMKV)” is being reported for the first time. Our indexing study by nucleic acid hybridization test showed the presence of JLYMKV in a high percentage (>84 %) of plants collected from Katernyiaghat. Therefore, JLYMKV may be considered a threat to J. curcas cultivation in India.
References
Singh P, Singh S, Mishra SP, Bhatia SK (2010) Molecular characterization of genetic diversity in Jatropha curcas L. Genes Genomes Genomics 4:1–8
Narayana DSA, Shankarappa KS, Govindrappa MR, Prameela HA, Gururaj Rao MR, Rangaswamy KT (2006) Natural occurrence of Jatropha mosaic virus disease in India. Curr Sci 91:584–586
Narayana DSA, Rangaswamy KT, Shankarappa KS, Maruthi MN, Lakshminarayana Reddy CN, Rekha AR, Keshava Murthy KV (2007) Distinct begomoviruses closely related to Cassava mosaic viruses cause Indian jatropha mosaic disease. Int J Virol 3:1–11
Tewari (2007) Jatropha and biodiesel. Ocean Books Ltd
Raj SK, Snehi SK, Kumar S, Khan MS, Pathre U (2008) First molecular identification of a begomovirus in India that is closely related to Cassava mosaic virus and causes mosaic and stunting of Jatropha curcas L. Australas Plant Dis Notes 3:69–71
Gao S, Qu J, Chua NH, Ye J (2010) A new strain of Indian cassava mosaic virus causes a mosaic disease in the biodiesel crop Jatropha curcas. Arch Virol 155:607–612
Ramkat RC, Alberto C, Fatemeh M, Laimer M (2011) Biotechnological approaches to determine the impact of viruses in the energy crop plant Jatropha curcas. Virol J 3:386
Snehi SK, Srivastava A, Raj SK (2012) Biological characterization and complete genome sequence of a possible strain of Indian cassava mosaic virus from Jatropha curcas in India. J Phytopathol 160:547–553
Kashina BD, Alegbejo MD, Banwo OO, Nielsen SL, Nicolaisen M (2013) Molecular identification of a new begomovirus associated with mosaic disease of Jatropha curcas L in Nigeria. Arch Virol 158(2):511–514
Srivastava A, Jaidi M, Kumar S, Raj SK (2014) Molecular identification of a new begomovirus associated with leaf crumple disease of Jatropha curcas L in India. Arch Virol. doi:10.1007/s00705-014-2288-8
Singh R, Raj SK, Prasad V (2008) Molecular characterization of a strain of Squash leaf curl China virus from North India. J Phytopathol 156(4):222–228
Reddy CRV, Colvin J, Muniyappa V, Seal S (2005) Diversity and distribution of begomoviruses infecting tomato in India. Arch Virol 150:845–867
Rojas MR, Gilbertson RL, Russell DR, Maxwell DP (1993) Use of degenerate primers in the polymerase chain reaction to detect whitefly transmitted geminiviruses. Plant Dis 77:340–347
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol Biol Evol 30:2725–2729
Fauquet CM, Briddon RW, Brown JK, Moriones E, Stanley J, Zerbini M, Zhou X (2008) Geminivirus strain demarcation and nomenclature. Arch Virol 153(4):783–821
Fienberg AP, Vogelstein B (1983) A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Ann Biochem 132:6
Acknowledgments
The authors are thankful to the Director, CSIR-National Botanical Research Institute, Lucknow, India, for facilities, and the Department of Biotechnology (DBT), India, for funding.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Srivastava, A., Kumar, S., Jaidi, M. et al. Molecular characterization of a new begomovirus associated with leaf yellow mosaic disease of Jatropha curcas in India. Arch Virol 160, 1359–1362 (2015). https://doi.org/10.1007/s00705-015-2375-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-015-2375-5