
Abstract
In this study, a model for estimating actual transpiration (T) is developed and tested for three tree species (Olea europaea, Citrus sinensis, Pinus pinea), growing in rainfed conditions in urban environment under Mediterranean semi-arid climate. The model, previously tested on many field crops, is based on the “big leaf” Penman-Monteith (PM) formulation of T in which canopy resistance (rc) is modeled with respect to local climatological conditions in different plant water status. Here rc is expressed as a function of available energy, vapor pressure deficit and aerodynamic resistance, and the trees’ water status was evaluated by means of crop water stress index. On an hourly scale, the comparison with T measured by sap flow thermal dissipation technique shows good performances of the model for all investigated species under contrasting water stress conditions. At daily scale, T modeled is less accurate for all species, because of the lack of stationarity conditions in the PM approach. At seasonal scale, however, the model gives good estimation of T, with an underestimation of − 1% for Olea and Pinus and overestimation of + 8% for Citrus with respect to the measured T values. The proposed model needs species-specific experimental calibration, but it may have good perspective of applicability in supporting water irrigation planning in urban forestry.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Transpiration of urban trees is yet a highly uncertain term of water balance of towns (Pataki et al. 2011a; Litvak et al. 2017). The uncertainty is due to the difficulties in both measurement and modeling of transpiration in urban environments, because of the large heterogeneity of vegetated surfaces. In particular, the quantification of actual transpiration rate of urban vegetation is complicated by different responses to water and energy availabilities by different cohabitant species growing in the same area (Chen et al. 2011, 2012).
Transpiration is part of latent heat flux, a component of the energy balance which can be measured, at city scale, by the eddy covariance (EC) micrometeorological method (Aubinet et al. 2012). By EC approach, latent heat has been linked to the structure of urban environment at different time scales (Loridan and Grimmond 2012). However, in heterogenic vegetated surfaces, any detail can be given by EC about the transpiration of single tree species, by preventing accurate estimation of urban and regional water budgets (e.g., Shields and Tague 2012; Vahmani and Hogue 2014b) and, then, municipality water allocation planning (Pataki et al. 2011a). For these purposes, transpiration needs to be quantified at hourly, daily, and season scales, taking into account the different behavior of the vegetated surfaces involved.
Generally, two methods are currently used to estimate the evapotranspiration (ET, plant transpiration plus surface evaporation) or transpiration (T, when evaporation, E, is neglected) of vegetated urban surfaces.
The first method is a two-step model (e.g., Spano et al. 2009; Vahmani and Hogue 2014a, b; Litvak et al. 2017), where ET is calculated as product of the landscape coefficient (KL) and the reference evapotranspiration (ET0) by using the following equation:
where KL is a reworking of the crop coefficient Kc used for field crops, firstly introduced by Doorenbos and Kassam (1979) and updated by Allen et al. (1998) and expressed as function of (i) species-specific differences in transpiration, (ii) planting density, and (iii) micro-climatic conditions (Costello et al. 2000; Litvak et al. 2017); ET0 is usually calculated on the base of the Penman-Monteith (PM) combination equation (Monteith 1965) applied to a reference surface, usually grass, where canopy resistance (rc) is supposed to be constant in time and in different sites where it is applied (Katerji and Rana 2011). The value of this fixed resistance depends on the time scale considered for the calculation of ET0. The two-step model has the advantage of using the standard weather variables collected by standard weather stations. The accuracy of the ET values determined by Eq. (1) depends on two factors: firstly, on the accuracy of the ET0 determination in different geographical sites, then, on the accuracy of the KL values.
The second approach is the one-step model (e.g., Grimmond and Oke 1991; Mitchell et al. 2008; Chen et al. 2012; Ballinas and Barradas 2015), which directly calculates the ET of a crop without a step through a reference surface. This model applies the PM equation where rc is (i) specific for each species, (ii) function of climatic characteristics of the atmosphere above the urban surface inside the boundary layer, and (iii) depending on the crop water status. The parameterization of rc is based on modeling works (Jarvis 1976; Fanjul and Barradas 1985; Grimmond and Oke 1991; Barradas et al. 2004). The one-step model requires fewer computation steps and, consequently, it is less prone to error sources than the two-step model (Rana et al. 2012).
Typically, ET models have been applied to urban trees: (i) in well-watered conditions (Litvak et al. 2017); (ii) using empirical methods for taking into account the plant water stress (Grimmond and Oke 1991; Barradas et al. 2004; Spano et al. 2009; Ballinas and Barradas 2015; Riikonen et al. 2016); and (iii) in conditions where there were no effects of water shortage on plants (Chen et al. 2012). Indeed, the application of ET models in arid and semi-arid ecosystems, where rainfed crops often experience water stress conditions, is very challenging regarding the canopy resistance estimation. In fact, it is very difficult to estimate rc in different soil, climate, and crop water conditions. In fact, many experiments demonstrated that rc is affected, instantaneously, by solar radiation, vapor pressure deficit, leaf water potential, soil water content (see, for example, the reviews by Rana and Katerji 1998; Katerji and Rana 2011), and hormonal messages (Davies et al. 1986; Schulze 1986). A simple method for modeling this resistance term is a preliminary condition for applying the PM model in urban environments where the optimal water condition is not assured. However, the PM model was established on hourly scale which is not the best time scale choice for applicative purposes, usually performed at daily and seasonal scales. Furthermore, the response of canopy resistance to crop water conditions can be observed on an instantaneous scale, thus leading to important fluctuations of rc during the day (e.g., Rana et al. 1997a; Barradas et al. 2004); hence, the determination of rc throughout the mean daily value becomes arbitrary without a suitable site-specific analysis (Katerji and Rana 2011).
This study proposes a model of transpiration, based on one-step PM approach, applied to three rainfed species (Olea europaea L., Citrus sinensis L., and Pinus pinea L.) growing in an old multi-species garden in the downtown city under semi-arid Mediterranean climate conditions. The objectives are (i) to establish a criterion for identifying the trees’ water stress conditions, (ii) to model, at hourly scale, the canopy resistance by a deterministic approach based on the response of rc to the climate in different trees’ water conditions, (iii) to model at daily scale the actual trees’ transpiration through site-specific coefficients, and (iv) to estimate the trees’ water requirements at seasonal scale.
The key idea is to design a canopy resistance model in the PM equation, already successfully applied to open field crops and trees (Rana et al. 1994, 1997a, b, c, 2001, 2005), to estimate actual transpiration of rainfed urban trees.
2 Materials and methods
2.1 The site
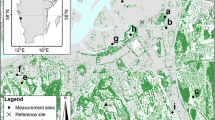
The investigated trees are part of a multi-species garden (surface 12,780 m2) located within the city of Bari in southern Italy on the Adriatic Sea (Fig. 1). The garden is owned by CREA-AA, and it is inside an area that comprises public and home buildings, public offices, schools, and the University Campus. The CREA-AA building hosts a research institute since more than one century, being established in 1881; the majority of trees (about 80%) was planted between the last decades of the nineteenth century and the first decades of the twentieth century.

The studied garden with the monitored trees (indicated with letters and numbers; O Olea, C Citrus, P Pinus)
The site is in the Mediterranean region (EEA 2016), submitted to “warm temperate with hot and dry summer” climate, following the Köppen-Geiger climate classification (Kottek et al. 2006). The mean annual temperature is 16.1 °C with a mean annual precipitation of 567 mm, calculated over the period 1995–2015. Moreover, using the aridity index defined by Holdridge et al. (1971), the climate of the region can be currently defined as dry (Ferrara et al. 2017).
The soil is classified as “Lithic Rhodoxeralf” and is characterized by clay texture, stable structure, cracked limestone subsoil, and fast drainage. The experiment was carried out in 2015 during the hottest period in the region, i.e., from 14 May to 2 August.
About 40% of the total courtyard area is occupied by trees’ projected crown; one half of this surface is occupied by Olea, Citrus, and Pinus trees (10.9%, 1.6%, 7.5% of surface, respectively, with 38 Olea, 42 Citrus, and 12 Pinus trees) as estimated by the Google® Planimeter tool; all other surface types in the courtyard are indicated in Table 1. To estimate the projected crown area, the crown radii of trees were measured in the four cardinal directions from the trunk for each tree (e.g., Karlik and McKay 2002). The management of the garden foresees the pruning of Citrus trees every 5 years (last pruning on 2013), while Olea trees are pruned every 6–8 years (last pruning on 2011); the other trees are never pruned except for safety issues.
2.2 The model of transpiration at hourly scale
The investigated area is covered by vegetation and impervious plus rooftops for about 47% and 46%, respectively (see Fig. 1, Table 1); consequently, plants’ transpiration consumes much more water than evaporation from soil (Amoroso et al. 2010; Sutanto et al. 2012). Moreover, the vegetated surface is well compacted due to no tillage and no irrigation in the garden, resulting in very low evaporation rates (Nassar and Horton 1999), only limited to short periods after rain (Symes and Connellan 2013). Furthermore, since the garden is crossed by paths and asphalted roads, after rain, the water rapidly runoff along these preferential ways, by preventing effective infiltration into the soil (Fini et al. 2017) and limiting surficial soil evaporation.
Actual crop transpiration was estimated by PM type model, which is considered appropriate also for urban inhomogeneous vegetated surfaces (Grimmond and Oke 1991; Ballinas and Barradas 2015, among others). In this model, which is theoretically applicable when the thermodynamic conditions are stationary, i.e., on hourly time scale, the latent heat flux due to transpiration, λTc (W m−2), of each studied species is written as follows:
Δ is the slope of the saturation pressure deficit versus temperature function in kPa °C−1; Q is the available energy in W m−2; ρ is the air density in kg m−3; cp is the specific heat of moist air in J kg−1 °C−1; D is the vapor pressure deficit in kPa; γ is the psychrometric constant in kPa °C−1; ra is the aerodynamic resistance in s m−1; rc is the bulk canopy resistance in s m−1; and λ is the latent heat of evaporation in J kg−1.
ra was calculated between the top of the trees and a reference point (z) where the measurements of weather variables were carried out, according to Perrier (1975) as follows:
where d (m) is the zero-plane displacement and is estimated by d = 0.67ht; ht (m) is the mean height of the trees of the same species; k is the von Kármán constant (0.40); and u* is the friction velocity (m s−1). The wind speed (m s−1) measured at the reference point z above the canopy was expressed as follows (e.g., Arya 2001):
where z0 (m) is the roughness length estimated by z0 = 0.1ht. z0 is a measure of the aerodynamic surface heterogeneity, in this case, vegetation (Ballinas and Barradas 2015). Therefore, the final formula for ra was written as follows:
The use of this relationship for the aerodynamic resistance introduces two simplifying assumptions on the homogeneity of the transpirative layer which needs to be evaluated: (i) ra was assumed to be the same for the whole tree canopy; (ii) ra was assumed to be the same for all trees of same species. The first simplification implies that the source/sink of boundary layer fluxes was at the same height, with no difference among leaves at different positions in the tree canopy. This assumption is supported by the study of Daudet et al. (1999), who found that wind velocity profile and direction weakly influenced the transpiration rate along the tree canopy; thus, the error can be considered negligible. The second assumption on ra implies that the calculation of d and z0 in Eq. (5) was made at the same height ht of all trees of the same species. A sensitivity analysis on ra (data not shown) with respect to these parameters showed that a variation of 50% of their values determined an error in the range 10–15% when the wind speed varied between 1 and 5 m s−1.
2.2.1 The canopy resistance parametrization
The PM formula is the combination of two terms: (i) the radiative term and (ii) the aerodynamic term. To clearly separate these two terms, Eq. (2) can be written under the following form:
From the above relation, it can be argued that there are only two cases in which λTc is equal to the radiation term (i.e., λTc independent of the aerodynamic term): (i) when ra > > 0, i.e., the surface is very smooth which is unusual in urban situation (Grimmond and Oke 1999) and/or the wind speed is very low; (ii) when rc takes the particular value:
This last variable is linked to the “isothermal resistance” introduced for the first time by Monteith (1965). It has also been called “critical resistance” (Daudet and Perrier 1968) because it represents a threshold between two conditions: (i) rc < r* and Tc increases with wind speed; (ii) rc > r* and Tc decreases with wind speed (Rana et al. 1994).
If we substitute Eq. (7) into Eq. (6), the PM formula can be written in a more symmetric form where the mentioned resistances r*, ra, and rc clearly appear in the aerodynamic term as:
The first term is the equilibrium transpiration (Jarvis 1976; Jarvis and McNaughton 1985), and the second fraction is a dimensionless quantity which provides the aerodynamic weighting on the equilibrium transpiration (McNaughton 1976) and can be seen as a crop-climatological coefficient: it modulates the part of the available energy transformed by the soil-canopy-atmosphere system during transpiration. It was experimentally demonstrated (e.g., Perrier et al. 1980; Rana et al. 1997b; Shi et al. 2008; Rana et al. 2012) that rc/ra was a function of r*/ra and this relationship depended on phenological stage and crop water status. In fact, following the fluid mechanics data analysis approach, a relationship between rc/ra and r*/ra can be argued from a dimensional analysis based on the Buckingham theorem (e.g., Kreith 1973) as:
Katerji and Perrier (1983) firstly proposed a linear relation for f in Eq. (9); in the next, many authors experimentally demonstrated the validity of such relationship and the accuracy in the estimation of actual evapotranspiration of different crops (see the synthesis in Katerji and Rana 2013).
The evaluation of the presented model were carried out through two steps: (i) the calibration, i.e., the search for the experimental function “f” linking the ratios rc/ra and r*/ra (see Eq. 9), and (ii) the validation with the comparison between calculated and measured actual transpiration values. In the present case (see Section 3.2), the best fitting function was found to be logarithmic for all species as:
2.2.2 The available energy Q and the measurement of weather variables
The energy available to the trees (Q) can be expressed, following a common nomenclature for urban energy balances (Grimmond and Oke 1991; Christen and Vogt 2004), as:
where Q* is the net all-wave radiation (W m−2), QF is the anthropogenic heat release (W m−2), QA is the net heat advection (W m−2), and QS is the sensible heat storage (W m−2). In this study, (i) Q* was measured by net radiometers; (ii) QF, including all additional energy inputs produced by human activities, such as the energy released by combustion of fuels, electric heating, air conditioning, and traffic, can be considered negligible since the garden is in a residential area with many public buildings and the traffic is strongly seasonal and limited to their opening/closing time (Loridan and Grimmond 2012); (iii) QA can be neglected because the experimental site is on the coast of Adriatic Sea, beneficiating of the sea breeze regime (Christen and Vogt 2004; Lemonsu et al. 2004; Loridan and Grimmond 2012); and (iv) QS was estimated according to the “objective hysteresis model” of Grimmond et al. (1991) as upgraded by Roberts et al. (2006):
where fk indicates the fractions of space occupied by each surface type. Namely, 6 types of surface were identified: grass, bare soil, mixed forest and shrubs, rooftops, impervious concrete, and asphalt. The coefficients a1, a2, and a3 were derived from independent studies on urban surface types and their values are reported in Table 1. t is the time interval (60 min, following the time interval of measurements).
A meteorological station (Weather transmitter WXT520, Vaisala, Helsinki, Finland) was set up on the CREA-AA building; it measured precipitation, temperature, and relative humidity of air, wind, speed, and direction. Global radiation was measured by a pyranometer (CMP3, Kipp & Zonen, Delft, The Netherlands). Net radiation (Q*, W m−2) was measured by two net radiometers (Q*6, REBS, USA). All measurements were taken by data loggers (CR800, CR10X, Campbell Scientific, Utah, USA) every 10 s, and the average values recorded every 60 min. The sensors of the meteorological station were installed on a 1.5-m-high mast mounted on a little tower (2 m height) on the roof of the building at 12 m from ground level. Thus, the reference level of all meteorological variables was set at z = 15.5 m from the ground.
2.3 The model of transpiration at daily and seasonal scales
To calculate daily Tc, two methods could be used.
-
(1)
Tc is estimated as the sum of hourly data so that:
with Tc,h is the hourly values expressed by Eq. (2). This method, theoretically the most correct, is not practical since the parameters measured on an hourly scale are needed.
-
(2)
The second method is based on a simplification of the relationship (8), so it can be written as:
where
The subscript d indicates the daytime average.
It was experimentally demonstrated (Katerji and Perrier 1983; Katerji et al. 1990; Rana et al. 1997a) that, for field crops in good water conditions, the Kd coefficient varies little with the r*/ra ratio and, in a first approximation, could be considered constant, while Kd varies considerably with the crop water status (Itier et al. 1992) and it seems that this last variation is a plant species characteristic (Rana et al. 1997b, c).
In the present study, in analogy to the hourly time scale, the coefficient Kd was calculated as function of the ratio (r*/ra)d, i.e., the daily average of r*/ra; the best fitting function was found to be logarithmic for all species as (see Section 3.2):
Transpiration at season scale was calculated by cumulating daily values.
2.4 The trees’ water conditions
The leaf water potential (LWP in MPa) measured with a pressure chamber has been demonstrated to be the most accurate method for determining the physiological water stress of plants (for trees, see, for example, the review by Wullschleger et al. 1998). For field crops, Rana et al. (1997a, b) demonstrated that the coefficient Kd can be modeled in function of predawn leaf water potential (PLWP), a good indicator of crop water stress (Katerji and Hallaire 1984; Tardieu et al. 1990). By common agreement, soil water shortage has not a significant influence on gas exchanges and, consequently, on crop growth if PLWP values are above a critical species-specific threshold. For applicative purpose, the measurement of PLWP cannot be routinely carried out, while a good alternative can be to consider the available soil water (AW), defined as the amount of water in the soil, normalized by the total water availability (between field capacity and wilting point). However, the accurate monitoring of soil moisture in urban gardens and parks is quite complicated due to the inhomogeneity of the areas since, as abovementioned, besides vegetation, different types of surfaces (bare soil, concrete, asphalt) are present (Nielsen et al. 2007).
Therefore, methods to detect water stress based on the surface temperature of the crop, like the Crop Water Stress Index (CWSI), can be a good compromise between accuracy and feasibility (Stanghellini and De Lorenzi 1994). In this study, CWSI is calculated in function of actual (Tact) and potential (Tp) transpiration (e.g., Jackson et al. 1981; Stanghellini and De Lorenzi 1994):
where Tp is calculated by the PM Eq. (2) with canopy resistance assumed equal to the minimum resistance (rmin). In this case, we used the following values of rmin: 200 s m−1 for Olea europaea, 160 s m−1 for Citrus sinensis (Körner et al. 1979), and 170 s m−1 for Pinus pinea (Kelliher et al. 1995).
The daily CWSI has been calculated adding up hourly transpiration values, considering only daytime ones.
2.4.1 The measured transpiration
The actual tree transpiration is needed to determine the CWSI values and to validate the proposed applicative transpiration model.
Tact was determined by sap flow heat dissipation method (HDM; Granier 1985, 1987). By HDM at first, the sap flow density Js0 (gH2O m−2 s−1) is determined in the outermost sapwood area, then Js0 must be extrapolated to all active water-conducting xylems of the sapwood area (SWA in m2). SWA was determined by measuring sapwood depth on a core collected with a 5-mm-diameter increment borer at breast height in the north side of monitored trees.
The monitored trees were chosen by combining two procedures: (1) the quantile upscaling method (Čermák et al. 2004) and (2) the analysis of the frequency distribution of stem diameters (Rana et al. 2005): 4 Olea, 4 Citrus, and 3 Pinus trees were monitored as shown in Fig. 1.
To avoid direct solar heating, the sap flow probes (20 mm, SFS2 Type M, UP, Steinfurt, Germany) were installed at breast height in the north side of each tree (Lindén et al. 2016), covered by a reflecting radiation screen, which also protects from rain. Js0 was continuously monitored every 10 s, and 10-min averages were stored by data loggers (three CR10X, Campbell Scientific, UT, USA). Hourly values were obtained by averaging the 10-min data.
To account for radial trends in sap flux density, according to Pataki et al. (2011b) and Litvak et al. (2012), the sapwood depth was divided in a set of 2-cm increments and generalized Gaussian functions were applied to estimate the sap flux density in each increment. We used two different functions for angiosperms and gymnosperms (Pataki et al. 2011b; Litvak et al. 2012) to calculate Jsi, i.e., the sap flux density in each increment i:
where x is the normalized depth of each sapwood increment (0 ≤ x < 1) and Js0 is calculated using the standard relationship in HDM:
where ΔT (*C) is the temperature difference between the heated upper probe and the lower reference one and ΔTmax (°C) is the maximum difference in temperature during night.
The whole tree transpiration of each sample was determined as:
where Tact is in g s−1, m is the number of 2-cm increments in sapwood depth, Jsi is the sap flux density (g m−2 s−1), and SWAi is the sap wood area at depth i. The tree’s transpiration per day (kg day−1) was calculated as the sum of hourly values during the daytime (global radiation, Rg > 20 W m−2). For each species, the considered transpiration is the mean of Tact of all sampled trees. The transpiration per unit canopy area was calculated in kg m−2 (mm of transpired water) as the value of Tact of the species divided by the mean projected canopy area.
2.5 Statistical analysis
Following Legates and McCabe (1999), the analysis of performances (Microsoft Excel® 32 bit) of the described model at hourly and daily scale included linear regression analysis, root mean square error (RMSE), mean absolute error (MAE), model efficiency (EF, range − ∞ to + 1, optimum value 1), and index of agreement (d, range 0 to + 1, optimum value 1).
2.6 Sensitivity analysis of the model
A sensitivity analysis of the model with respect to the calibration parameters, pi (i.e., the coefficients “a” and “b” in the logarithmic functions of Eq. 10), is carried out to investigate on how an error on these parameters is propagated to the estimation of tree transpiration.
Following Rana and Katerji (1998), the sensitivity analysis was carried out by calculating the non-dimensional relative sensitivity coefficients:
which represent that fraction of the change in pi that is transmitted to change λTc. Thus, at hourly scale, the sensitivity coefficients for a and b are expressed by the following expressions:
where
Negative coefficients indicate that a reduction in λTc results from an increasing of pi. By these coefficients, it is possible to evaluate the effect, on λTc, due to error on the determination of the input parameters. In fact, following the definition of relative error in a function due to relative errors in the input data, it is possible to write the relative error expression for transpiration as:
with ελTc, εa, and εb relative error on λTc, a, and b, respectively.
3 Results and discussion
3.1 The weather and the trees’ water conditions by CWSI
The daily patterns of Rg (between 7.8 and 31.0 MJ m−2 day−1), Ta (between 10.4 and 31.2 °C), and D (between 0.6 and 3.1 kPa) were typical of the region in the investigated season (data not shown). The precipitation amounts during the study period were high in the second half of May and, mostly, in mid-June (79 mm from 15 to 20 June, Fig. 2), which is usually dry in this area.

CWSI patterns at daily scale for all investigated trees species; precipitations are also shown
The CWSI showed, at hourly scale, very variable and spread values (data not shown) for all species, since it is function of input variables rapidly and instantaneously changing (Stanghellini and De Lorenzi 1994; Rana and Katerji 1998). However, at daily scale, the CWSI trends (Fig. 2) appeared clearer and able to suitably detect the water status of the investigated trees.
Berni et al. (2009) demonstrated that for Olea growing under Mediterranean climate, significant relationships can be found among the leaf stem potential, the canopy conductance, and the CWSI. Ben-Gal et al. (2009) showed that when an olive orchard in arid environment was irrigated with an amount of water between 100 and 50% of potential ET, the daily CWSI values were between 0.2 and 0.4, while when the water supplied was only 25% of potential ET, then the CWSI was around 0.7. Agam et al. (2013) showed that the maximum value of CWSI (calculated by the standard approach as in the present case) achieved by irrigated olive trees was again around 0.7. Therefore, in this study, we fixed at 0.7 the value of CWSI above that the Olea trees were in water stress (see Fig. 2).
Gonzalez-Dugo et al. (2013) in Citrus orchard found that any clear relationship can be envisaged between LWP and CWSI and that the CWSI values are always below 0.6–0.7 even if the irrigation was stopped for inducing water stress. Similar results were found by Waldo and Schumann (2009) and Ballester et al. (2013), and these last authors showed a maximum value of CWSI around 0.5 also when the Citrus trees were induced to water stress. Gonzalez-Dugo et al. (2014) found that when Citrus CWSI values attained 0.85, the crop was under severe water stress and when CWSI was close to 0, it was in optimal water conditions. In this case, Citrus trees had CWSI values almost always between 0.6 and 0.8 (see Fig. 2) without any significant trend. Furthermore, all studied Citrus trees were affected by a typical disease of this species, Dialeurodes citri (Homoptera: Aleyrodidae), which causes damage to crop directly through suck the sap and reducing plant vitality as well as the excretion of honeydew colonized by sooty mold fungi that reduce gas exchanges, which means photosynthesis, respiration, and transpiration (Rapisarda and Cocuzza 2017; Argov et al. 1999). Therefore, it was supposed that Citrus were under water stress conditions all time during this study.
For Pinus trees and in general for conifers, the measurement of water conditions through the CWSI was very rare. Seidel et al. (2016) found that the maximum CWSI value achieved by irrigated scots pine trees was 0.8, which was considered as the threshold value between stress and no-stress condition in this case study (see Fig. 2).
Hourly values of rc during daytime (i.e., between 7:00 and 18:00), obtained by inverting Eq. (2) with Tc and all inputs measured, are shown in Fig. 3 for three species under investigation. Two days is reported as example: 19 June (wet), after 2 days of huge rain (76 mm in total), and 29 July, 34 days after this huge precipitation (dry). On 19 June, the daily values of CWSI were 0.46, 0.74, and 0.57 for Olea, Citrus, and Pinus, respectively, hence below the thresholds previously established for the water stress conditions for Olea and Pinus, while above water stress threshold for Citrus: Citrus trees’ seems did not react to the huge rain in terms of transpiration due to the effect of their pest. On 29 July, the values of CWSI were 0.88, 0.70, and 0.92 for Olea, Citrus, and Pinus, respectively, hence above the water stress threshold values for all species. Under well-watered conditions during wet day, daytime rc were in the range 274–706 and 299–503 s m−1, with means of 359, and 347 s m−1 for Olea and Pinus respectively; these values are greater than those published by other authors for Olea (Berni et al. 2009) under good water conditions, while the values of rc found for Pinus are in line with those found in similar environments (Hoshika et al. 2017).

Canopy resistance (rc) patterns at hourly scale during two sample days
During dry days, under water stress rc was in the range 904–2995 and 1480–3286 s m−1 with means of 1382, and 2118 s m−1 for Olea and Pinus respectively, in this case, a comparison with other studies is very difficult because of the large variability in the relationships between drought conditions and trees’ water status.
For Citrus trees, rc ranged between 460 and 1485 s m−1 on 19 June, and between 302 and 2018 s m−1 on 29 July, and the mean values during wet and dry days were very similar (786 and 893 s m−1, respectively). These high values confirm that these trees were under water stress conditions, with a typical pattern of rc for stressed tree species (see Olea and Pinus behavior) with closure of stoma during hours of the day with decreasing values of solar radiation (Ballester et al. 2013; Gonzalez-Dugo et al. 2014).
Based on these considerations, it was supposed that Olea and Pinus trees were in well-watered conditions from the beginning of the experiments until 8 July, then they were under water stress; Citrus trees were under water stress conditions during the whole experimental period.
3.2 The model at hourly scale
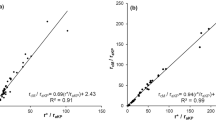
In Fig. 4, the experimental relationships between rc/ra and r*/ra were shown for the three species under investigation in the different water conditions. Each point represented the mean of all available hourly values during daytime (12 values from 7:00 to 18:00) in the 4 days 14–17 May (at the beginning of the experimental period in wet soil conditions) and in the 4 days 9–12 July (at the beginning of the period when also Olea and Pinus trees were under water stress conditions). The number of days for the calibration was suggested by Rana et al. (1997a, b). The logarithmic best fitting relationship between rc/ra and r*/ra are shown in Table 2: the dependence of rc/ra ratio on r*/ra was very clear for any water stress condition and for each species and quite robust (high values of determination coefficient r2). The values of slopes of the logarithmic relationships were higher when the trees were under water stress, having higher values of rc than the well-watered conditions for the same value of ra at given weather conditions as expressed by r*.

Calibration of the presented model at hourly scale (see text for details)
The comparison between calculated (λTc) and measured (λTact) hourly transpiration is shown in Fig. 5 for all species under investigation, excluding calibration days. Water stress and no-stress conditions are also indicated, but the regression has been made without considering water conditions, obtaining good correlation for all species. In the statistical comparison between measured and calculated values, the t test indicated that, in all cases, the intercept was significantly not different from zero at the 95% significance level; therefore, the analysis was done forcing the linear regression model through zero. Considering the whole experimental period, the cumulated values of estimated transpiration by the presented model indicated overestimation of 8% and 4% for Olea and Pinus, respectively, and underestimation of − 4% for Citrus with respect to the measured cumulated values. Other statistical indicators are shown in Table 3 indicating that the model had pretty good performances at hourly scale, with low values of the errors (RMSE and MAE), acceptable values of EF and high values of d.

Comparison between calculated (λTc) and measured transpiration (λTact) at hourly scale; trees’ water conditions are also indicated for the species under investigation
3.3 The model at daily and seasonal scales
To calibrate the relationship between Kd and (r*/ra)d (Fig. 6), we used the daily values of the same days used to calibrate the hourly model (i.e., 14–17 May and 9–12 July). Like in Fig. 5, water stress and no-stress conditions are also indicated using different symbols, but the logarithmic relationships are obtained considering all data together: the parametrization is quite robust with high values of determination coefficients also at daily scale. For Olea and Pinus, the values of Kd were generally lower in water stress conditions than in well-watered conditions. The coefficient Kd calculated by these logarithmic functions was used to calculate the actual transpiration (Eq. 14) at daily scale.

Calibration of the model at daily scale (see text for details)
A comparison between daily measured and calculated Tc is shown in Fig. 7 for the three species under investigation. For Olea and Pinus, Tc ranged between low values before precipitations and high values afterwards, while Citrus seems to increase its transpiration during dry periods (after 8 July). During rainy days, transpiration decreased and the model reproduced these events.

Patterns of calculated and measured transpiration (Tc) at daily scale for the species under investigation during the experimental period
The linear regression statistical analysis between the modeled and measured Tc is shown in Fig. 8 and both well-watered and stress conditions are also indicated, but the regression is done considering all together and excluding the calibration days. In this case, the comparison showed that the daily model is much less accurate (low R2) than the model at hourly scale. The other statistical indicators (see Table 3) indicated that at daily scale, the model presented worst performances than at the hourly scale, with pretty high values of RMSE and MAE and low values of EF, even if the index of agreement d had pretty high values. The loss of accuracy of the daily model with respect to the hourly one was due to the lack of stationarity conditions. However, when the model is evaluated at seasonal scale, by comparing cumulated Tc (as a sum of the daily values in mm), it shows good performances; in fact, it underestimates of − 1% Olea and Pinus transpiration and overestimates of 8% Citrus transpiration.

Comparison between calculated (Tcd) and measured transpiration (Tact) at daily scale; trees’ water conditions are also indicated for the species under investigation
3.4 Error analysis
The relative error on λTc was calculated by Eq. (26) where the terms Sa and Sb are the mean of all available values in the experimental period. Error on λTc due to error in the input parameter a is negligible (data not shown), being between 0 and 1% when the input parameter a varies between 0 and 100% of the experimentally calculated value. The relative error on λTc due to error in the input parameter b is illustrated in Fig. 9 for both stress and no-stress conditions. For Pinus, the error on λTc is always low, less than 10%; for Olea, the error is > 10% when the error on the coefficient b is greater than 50%. For Citrus, the error on λTc is great than 20% when the error on b is greater than 35%.

The relative error on calculated transpiration due to error in the input parameter b (see Eq. (10)) for both stress and no-stress conditions
4 Conclusions
In this study, a transpiration model is presented for three species of urban trees at hourly and daily scales, neglecting the evaporation. The model is based on the PM formula, with the introduction of a semi-empirical parametrization for the canopy resistance. Either at hourly or at daily scale, the canopy resistance is estimated from (i) climatological data and (ii) parameters depending on the species following the plant water status as expressed by the crop water stress index. The species-specific model parameters need experimental calibration.
In this model, canopy resistance is a variable parameter, assuming a specific value at each moment and, consequently, for each day.
The model has been tested in a site of Mediterranean region, under semi-arid climate. On an hourly scale, it seems to work well in contrasting water conditions: when the tree is well watered and under water stress. The performance of the model could be improved by the use of more than two water stress classes; in this case, each interval would be narrow, but the model loses practical applicability.
On a daily scale, the model was not as accurate as at hourly scale giving, however, acceptable estimation of transpiration. It works better at seasonal scale and should be used to determine the trees’ water requirements or as output term of urban water balance.
The difficulty in the application of the present model is in the determination of the trees’ water status, that here is determined in function of the measured transpiration, which is the unknown variable. Therefore, further developments are necessary to improve its application especially at daily scale, in particular using estimates of CWSI based on surface temperature (Jackson et al. 1981; Stanghellini and De Lorenzi 1994) instead of measurements of transpiration. In fact, an alternative applicative definition of CWSI is:
with Ts surface temperature measurable for example by infrared thermometry, Tm and TM minimum and maximum tree temperature, respectively, easily calculable by climatological variables (e.g., Stanghellini and De Lorenzi 1994).
Furthermore, Rana et al. (1997b, 2001) for field crops demonstrated that the presented approach does not need a local calibration, like other Penman and PM-based models, but only a “per crop” calibration. If the coefficients “a,” “b,” “A,” and “B” of the presented model are known for different crop water conditions, the model would be valid at another site. This hypothesis clearly needs to be verified for trees species in urban environments with further research.
References
Agam N, Cohen Y, Bernic JAJ, Alchanatis V, Kool D, Dag A, Yermiyahu U, Ben-Gal A (2013) An insight to the performance of crop water stress index for olive trees. Agric Water Manag 113:79–86
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration. Guidelines for computing crop water requirements. Irrigation and Drainage paper No. 56. FAO, Rome, p 300
Amoroso G, Frangi P, Piatti R, Fini A, Ferrini F (2010) Effect of mulching on plant and weed growth, substrate water content, and temperature in container-grown giant arborvitae. HortTechnology 20:957–962
Argov Y, Rössler Y, Voet H, Rosen D (1999) The biology and phenology of the citrus whitefly, Dialeurodes citri, on citrus in the Coastal Plain of Israel. Entomologia Experimentalis et Applicata 93:21–27
Arya SP (2001) Introduction to micrometeorology. Academic Press, London, 420 pp
Asaeda T, Ca VT (1993) The subsurface transport of heat and moisture and its effect on the environment: a numerical model. Bound.-Layer Meteor. 65:159–179
Aubinet M, Vesala T, Papale D (2012) Eddy covariance: a practical guide to measurement and data analysis. Springer, Dordrecht, Heidelberg, London, New York 438 pp
Ballester C, Jiménez-Bello MA, Castel JR, Intrigliolo DS (2013) Usefulness of thermography for plant water stress detection in citrus and persimmon trees. Agric For Meteorol 168:120–129
Ballinas M, Barradas VL (2015) The urban tree as a tool to mitigate the urban heat island in Mexico city: a simple phenomenological model. J. Environ. Qual. https://doi.org/10.2134/jeq2015.01.0056
Barradas VL, Ramos-Vázquez A, Orozco-Segovia A (2004) Stomatal conductance in a tropical xerophilous shrubland at a lava substratum. Int J Biometeorol 48:119–127. https://doi.org/10.1007/s00484-003-0195-x
Ben-Gal A, Agam N, Alchanatis V, Cohen Y, Yermiyahu U, Zipori I, Presnov E, Sprintsin M, Dag A (2009) Evaluating water stress in irrigated olives: correlation of soil water status, tree water status, and thermal imagery. Irrig Sci 27:367–376
Berni JAJ, Zarco-Tejada PJ, Sepulcre-Cantó G, Fereres E, Villalobos F (2009) Mapping canopy conductance and CWSI in olive orchards using high resolution thermal remote sensing imagery. Remote Sens Environ 113:2380–2388
Brutsaert WH (1982) Evaporation into atmosphere: theory, history and application. D. Reidel, Dordrecth, p 299
Čermák J, Kučera J, Nadezhdina N (2004) Sap flow measurements with some thermodynamic methods, flow integration within trees and scaling up from sample trees to entire forest stands. Trees 18:529–546. https://doi.org/10.1007/s00468-004-0339-6
Chen L, Zhang Z, Li Z, Tang J, Caldwell P, Zhang W (2011) Biophysical control of whole tree transpiration under an urban environment in northern China. J Hydrol 402:388–400
Chen L, Zhang Z, Ewers BE (2012) Urban tree species show the same hydraulic response to vapor pressure deficit across varying tree size and environmental conditions. PLoS One 7(10):47882. https://doi.org/10.1371/journal.pone.0047882
Costello LR, Matheny N, Clark J, Jones K (2000) A guide to estimating irrigation water needs of landscape plantings in California. In: The landscape coefficient method and WUCOLS III. University of California Cooperative Extension California Department of Water Resources, Sacramento
Christen A, Vogt R (2004) Energy and radiation balance of a central European city. Int J Climatol 24:1395–1421
Daudet FA, Perrier A (1968) Etude de l’evaporation ou de la condensation a la surface d’un corp a partir du bilan energetique. Rev Gen Therm 76:353–364
Daudet FA, Le Roux X, Sinoquet H, Adam B (1999) Wind speed and leaf boundary layer conductance variation within tree crown consequences on leaf-to-atmosphere coupling and tree functions. Agric. For. Meteorol. 97:171–185
Davies WJ, Metcafle J, Lodge TA, da Costa AR (1986) Plant growth substances and the regulation of growth under drought. Aust J Plant Physiol 13:105–125
Doll D, Ching JKS, Kaneshiro J (1985) Parameterisation of subsurface heating for soil and concrete using net radiation data. Bound.-Layer Meteor. 32:351–372
Doorenbos, J., Kassam, A.H., 1979. Yield response to water. U.N. Food and Agriculture Organization Irrigation and Drainage Paper No. 33, Rome
EEA (European Environment Agency) (2016) Mapping and assessing the condition of Europe’s ecosystems: progress and challenges. Report No 3/2016, 148 pp.
Fanjul L, Barradas VL (1985) Stomatal behaviour of two heliophile understorey species of a tropical deciduous forest in Mexico. J Appl Ecol 22:943–954. https://doi.org/10.2307/2403242
Ferrara RM, Mazza G, Muschitiello C, Castellini M, Stellacci AM, Navarro A, Lagomarsino A, Vitti C, Rossi R, Rana G (2017) Short-term effects of conversion to no-tillage on respiration and chemical - physical properties of the soil: a case study in a wheat cropping system in semi-dry environment. Italian Journal of Agrometeorology 1:47–58. https://doi.org/10.19199/2017.1.2038-5625.047
Fini A, Frangi P, Moria J, Donzelli D, Ferrini F (2017) Nature based solutions to mitigate soil sealing in urban areas: results from a 4-year study comparing permeable, porous, and impermeable pavements. Environ Res 156:443–454
Gonzalez-Dugo V, Zarco-Tejada P, Nicolás E, Nortes PA, Alarcón JJ, Intrigliolo DS, Fereres E (2013) Using high resolution UAV thermal imagery to assess the variability in the water status of five fruit tree species within a commercial orchard. Precision Agric 14:660–678
Gonzalez-Dugo V, Zarco-Tejada PJ, Fereres E (2014) Applicability and limitations of using the crop water stress index as an indicator of water deficits in citrus orchards. Agric For Meteorol 198–199:94–104
Granier A (1985) Une nouvelle méthode pour la mesure du flux de sève brute dans le tronc des arbres. Ann Sci For 42:81–88
Granier A (1987) Mesure du flux de sève brute dans le tronc du Douglas par une nouvelle méthode thermique. Ann Sci For 44:1–14
Grimmond CSB, Oke TR (1991) An evapotranspiration-interception model for urban areas. Water Resour Res 27:1739–1755
Grimmond CSB, Cleugh HA, Oke TR (1991) An objective urban heat storage model and its comparison with other schemes. Atmospheric Environment B 25:311–326
Grimmond CSB, Oke TR (1999) Aerodynamic properties of urban areas derived from analysis of surface form. J Appl Meteorol 38:1262–1292
Holdridge LR, Grenke WC, Hatheway WH, Liang T, Tosi JA (1971) Forest Environments in Tropical Life Zones: a pilot study. Pergamon Press, Oxford.
Hoshika Y, Fares S, Savi F, Gruening C, Goded I, De Marco A, Sicard P, Paoletti E (2017) Stomatal conductance models for ozone risk assessment at canopy level in two Mediterranean evergreen forests. Agric For Meteorol 234–235:212–221
Itier B, Flura D, Belabbes K, Kosuth P, Rana G, Figueiredo L (1992) Relations between relative evapotranspiration and predawn leaf water potential in soybean grown in several locations. Irrig Sci 13:109–114
Jackson RD, Idso SB, Reginato RL, Pinter PJ Jr (1981) Canopy temperature as a crop water stress indicator. Water Resour Res 17:1133–1141
Jarvis PG (1976) The interpretation of the variation in leaf water potential and stomatal conductance found in canopies. Phil Trans R Soc Lond Ser B 273:593–610
Jarvis PG, McNaughton KG (1985) Stomatal control of transpiration: scaling up from leaf to region. Adv Ecol Res 15:1–49
Karlik, J.F., McKay, A.H., (2002). Leaf area index, leaf mass density, and allometric relationships derived from harvest of blue oaks in a California oak savanna. USDA Forest Service Gen. Tech. Rep. PSW-GTR-184
Katerji N, Ferreira I, Mastrorilli N, Losavio N (1990) A simple equation to calculate crop evapotranspiration: results of several years of experimentation. Acta Hortic 278:477–489
Katerji N, Hallaire M (1984) Les grandeurs de reference utilisables dam l’etude de l’alimentation en eau des cultures. Agronomie 4(10):999–1008
Katerji N, Perrier A (1983) Modelisation de l’evapotranspiration reelle d’une parcelle de luzerne: role dune coefficient culturale. Rev Agron 3(6):513–521
Katerji N, Rana G (2011) Crop reference evapotranspiration: a discussion of the concept, analysis of the process and validation. Water Resour Manag 25:1581–1600
Katerji N, Rana G (2013) FAO-56 methodology for determining water requirement of irrigated crops: critical examination of the concepts, alternative proposals and validation in Mediterranean region. Theor Appl Climatol 116(3):515–536
Kelliher FM, Leuning R, Raupach MR, Schulze E-D (1995) Maximum conductances for evaporation from global vegetation types. Agri For Metereol 73:1–16
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World map of the Köppen-Geiger climate classification updated. Meteorol Z 15(3):259–263
Körner C, Scheel JA, Bauer H (1979) Maximum leaf diffusive conductance in vascular plants. Photosynthetica 13(1):45–82
Kreith F (1973) Principle of heat transfer. Dun Donnelley, New York, 651 pp
Legates DR, McCabe GJ (1999) Evaluating the use of ‘goodness-of fit’ measures in hydrologic and hydroclimatic model validation. Wat Resou Res 35:233–241
Lemonsu A, Grimmond CSB, Masson V (2004) Modeling the surface energy balance of the core of an old Mediterranean city: Marseille. J Appl Met 73:312–327
Lindén J, Fonti P, Espera J (2016) Temporal variations in microclimate cooling induced by urban trees in Mainz. Germany Urb For Urb Gr 20:198–209
Litvak E, McCarthy HR, Pataki DE (2012) Transpiration sensitivity of urban trees in a semi-arid climate is constrained by xylem vulnerability to cavitation. Tree Physiol 32:373–388
Litvak E, McCarthy HR, Pataki DE (2017) A method for estimating transpiration of irrigated urban trees in California. Landsc Urban Plan 158:48–61
Loridan T, Grimmond CSB (2012) Characterization of energy flux partitioning in urban environments: links with surface seasonal properties. Am Meteorol Soc 51:219–241. https://doi.org/10.1175/JAMC-D-11-038.1
McCaughey J (1985) Energy balance storage terms in a mature mixed forest at Petawawa Ontario—a case study. Bound-Layer Meteor 31:89–101
McNaughton KG (1976) Evaporation and advection. I. Evaporation from extensive homogeneous surfaces. Q J R Meteorol Sot 102:181–191
Mitchell VG, Cleugh HA, Grimmond CSB, Xu J (2008) Linking urban water balance and energy balance models to analyse urban design options. Hydrol Process 22:2891–2900
Monteith JL (1965) Evaporation and atmosphere. The state and movement of water in living organisms. Symposia of the Society for Experimental Biology 19:205–234
Nassar IN, Horton R (1999) Salinity and compaction effects on soil water evaporation and water and solute. Soil Sci Soc Am J 63:752–758. https://doi.org/10.2136/sssaj1999.634752x
Nielsen CN, Bühler O, Kristoffersen P (2007) Soil water dynamics and growth of street and park trees. Arboriculture Urban Forest 33:231–245
Novak, M. D. (1981) The moisture and thermal regimes of a bare soil in the Lower Fraser Valley during spring. Ph.D. thesis, The University of British Columbia, Vancouver, BC, Canada, 153 pp.
Offerle B, Grimmond CSB, Fortuniak K (2005) Heat storage and anthropogenic heat flux in relation to the energy balance of a central European city centre. Int J Climatol 25:1405–1419
Pataki DE, Boone CG, Hogue TS (2011a) Socio-ecohydrology and the urban water challenge. Ecohydrology 347:341–347. https://doi.org/10.1002/eco
Pataki DE, McCarthy HR, Litvak E, Pincetl S (2011b) Transpiration of urban forests in the Los Angeles metropolitan area. Ecol Appl 21(3):661–677
Perrier A (1975) Etude physique de l’évapotranspiration dans les conditions naturelles. I. evaporation et bilan d’énergie des surface naturelles. Annals of Agronomy, 26, 1–18. II. Expression et paramètres donnant l’évapotranspiration réelle d’une surface mince. Annals of Agronomy, 26, 105–123. III. Evapotranspiration réelle et potentielle des couverts végétaux. Annals of Agronomy 26:229–243 (in French)
Perrier A, Katerji N, Iier B (1980) Etude ‘in situ’ de l’evapotranspiration reele dune culture de ble. Agric Meteorol 21:295–311
Rana G, Katerji N, Mastrorilli M, El Moujabber M (1994) Evapotranspiration and canopy resistance of grass in a Mediterranean region. Theor Appl Climatol 50(1–2):61–71
Rana G, Katerji N (1998) A measurement based sensitivity analysis of the Penman-Monteith actual evapotranspiration model for crops of different height and in contrasting water status. Theor Appl Climatol 60:141–149
Rana G, Katerji N, Mastrorilli M, El Moujabber M (1997a) A model for predicting actual evapotranspiration under soil water stress in a Mediterranean region. Theor Appl Climatol 56(1–2):45–55
Rana G, Katerji N, Mastrorilli M, El Moujabber M, Brisson N (1997b) Validation of a model of actual evapotranspiration for water stressed soybeans. Agric For Meteorol 86:215–224
Rana G, Katerji N, Mastrorilli M (1997c) Environmental and soil-plant parameters for modelling actual crop evapotranspiration under water stress conditions. Ecol Model 101:363–371
Rana G, Katerji N, Perniola M (2001) Evapotranspiration of sweet sorghum: a general model and multilocal validity in semi arid environmental conditions. Water Resour Res 37(12):3237–3246
Rana G, Katerji N, De Lorenzi F (2005) Measurement and modelling of evapotranspiration of irrigated citrus orchard under Mediterranean conditions. Agric For Meteorol 128(3–4):199–209
Rana G, Katerji N, Lazzara P, Ferrara RM (2012) Operational determination of daily actual evapotranspiration of irrigated tomato crops under Mediterranean conditions by one-step and two-step models: multiannual and local evaluations. Agric Water Manag 115:285–296
Rapisarda, C., Cocuzza, G.E.M. (Eds.), 2017. Integrated pest management in tropical regions. CABI, UK, 359 pp, ISBN 1780648006, 9781780648002
Riikonen A, Järvi L, Nikinmaa E (2016) Environmental and crown related factors affecting street, tree transpiration in Helsinki, Finland. Urban Ecosyst 19:1693–1715. https://doi.org/10.1007/s11252-016-0561-1
Roberts SM, Oke TR, Grimmond CSB, Voogt JA (2006) Comparison of four methods to estimate urban heat storage. J Appl Met Climat 45:1766–1780
Shi T, Guan D, Wang A, Wu J, Jin C, Han S (2008) Comparison of three models to estimate evapotranspiration for a temperate mixed forest. Hydrol Process 22(17):3431–3443
Shields CA, Tague CL (2012) Assessing the role of parameter and input uncertainty in ecohydrologic modeling: implications for a semi-arid and urbanizing coastal California catchment. Ecosystems 15:775–791. https://doi.org/10.1007/s10021-012-9545-z
Schulze ED (1986) Carbon dioxide and water vapour exchange in response to drought in the atmosphere and in the soil. Ann Rev Plant Physiol 37:247–270
Seidel H, Schunk C, Matiu M, Menzel A (2016) Diverging drought resistance of scots pine provenances revealed by infrared thermography. Frontiers in Plant Science 7:Article 1247. https://doi.org/10.3389/fpls.2016.01247
Spano D, Snyder RL, Sirca C, Duce P (2009) ECOWAT-A model for ecosystem evapotranspiration estimation. Agric For Meteorol 149(10):1584–1596. https://doi.org/10.1016/j.agrformet.2009.04.011
Stanghellini C, De Lorenzi F (1994) A comparison of soil- and canopy temperature-based methods for the early detection of water stress in a simulated patch of pasture. Irrig Sci 14:141–146
Symes P, Connellan G (2013) Water management strategies for urban trees in dry environments: lessons for the future. Arboricult Urban For 39:116–124
Sutanto SJ, Wenninger J, Coenders-Gerrits AMJ, Uhlenbrook S (2012) Partitioning of evaporation into transpiration, soil evaporation and interception: a comparison between isotope measurements and a HYDRUS-1D model. Hydrol Earth Syst Sci 16:2605–2616
Tardieu F, Katrerji N, Bethenod O (1990) Relation entre l’état hydrique du sol, le potentiel de base et d’autres indicateurs del a contrainte hydrique chez le mais. Agronomie 8:617–626
Vahmani P, Hogue TS (2014a) High-resolution land surface modeling utilizing remote sensing parameters and the Noah UCM: a case study in the Los Angeles Basin. Hydrol Earth Syst Sci 18(12):4791–4806. https://doi.org/10.5194/hess-18-4791-2014
Vahmani P, Hogue TS (2014b) Incorporating an urban irrigation module into the Noah land surface model coupled with an urban canopy model. Journal of Hydrometeorology:140421133412009. https://doi.org/10.1175/JHM-D-13-0121.1
Waldo LI, Schumann AW (2009) Alternative methods for determining crop water status for irrigation of citrus groves. Proc Fla State Hort Soc 122:63–71
Wullschleger SD, Meinzer FC, Vertessy RA (1998) A review of whole-plant water use studies in trees. Tree Physiol 18:499–512
Yoshida A, Tominaga K, Watatani S (1990–91) Field measurements on energy balance of an urban canyon in the summer season. Energy Build. 15–16:417–423
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Rana, G., Ferrara, R.M. & Mazza, G. A model for estimating transpiration of rainfed urban trees in Mediterranean environment. Theor Appl Climatol 138, 683–699 (2019). https://doi.org/10.1007/s00704-019-02854-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00704-019-02854-4