
Abstract
The eddy covariance technique was used to measure the CO2 flux over four differently grazed Leymus chinensis steppe ecosystems (ungrazed since 1979 (UG79), winter grazed (WG), continuously grazed (CG), and heavily grazed (HG) sites) during four growing seasons (May to September) from 2005 to 2008, to investigate the response of the net ecosystem exchange (NEE) over grassland ecosystems to meteorological factors and grazing intensity. At UG79, the optimal air temperature for the half-hourly NEE occurred between 17 and 20 °C, which was relatively low for semi-arid grasslands. The saturated NEE (NEEsat) and temperature sensitivity coefficient (Q 10) of ecosystem respiration (RE) exhibited clear seasonal and interannual variations, which increased with canopy development and the soil water content (SWC, at 5 cm). The total NEE values for the growing seasons from 2005 to 2008 were −32.0, −41.5, −66.1, and −89.8 g C m−2, respectively. Both the amounts and distribution of precipitation during the growing season affected the NEE. The effects of grazing on the CO2 flux increased with the grazing intensity. During the peak growth stage, heavy grazing and winter grazing decreased NEEsat and gross primary production (45 % for HG and 34 % for WG) due to leaf area removal. Both RE and Q 10 were clearly reduced by heavy grazing. Heavy grazing changed the ecosystem from a CO2 sink into a CO2 source, and winter grazing reduced the total CO2 uptake by 79 %. In the early growing season, there was no difference in the NEE between CG and UG79. In addition to the grazing intensity, the effects of grazing on the CO2 flux also varied with the vegetation growth stages and SWC.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Grasslands cover 32 % of global land surfaces (Parton et al. 1995). Natural grassland ecosystems in semi-arid areas can act as either CO2 sinks or sources, which depends mainly on variations in the amount and timing of precipitation (Gilmanov et al. 2007). Variations in the net ecosystem exchange (NEE) on half-hourly, daily, monthly, and longer time scales have attracted much attention in the past two decades (Aires et al. 2008; Li et al. 2005; Xu and Baldocchi 2004). However, long-term measurements are needed over different grassland ecosystems to understand the control of the CO2 flux under significantly different environmental and vegetation conditions.
Grassland comprises the most widespread ecosystem type in China, where it covers 41.7 % of the total land area (DAHV and CISNR 1996). The main type is temperate grassland in north China, which accounts for 41 % of the total grassland areas (DAHV and CISNR 1996). The typical steppe in the temperate zones represents the most widely distributed grassland ecosystem in the Eurasia typical steppe region, which is the largest contiguous grassland area in the world (Bai et al. 2004), where Leymus chinensis steppe is the main community type. The climate in most of this area is arid and semi-arid, and there was a gradual warming trend during the 1990s. The productivity of steppe ecosystems is expected to be highly sensitive to climate disturbances (e.g., drought and extremely heavy rainfall events) (Hao et al. 2008; Schaffrath et al. 2011).
Grazing is the major type of land management in grasslands. The impact of grazing on NEE is still uncertain although several experiments have been conducted in different areas (Chen et al. 2005, 2014; Lecain et al. 2002; Rogiers et al. 2005; Wang et al. 2011; Wilsey et al. 2002). Experiments have shown that there was no significant difference in NEE between ungrazed and grazed sites over shortgrass steppe in the USA and tropical pasture in Panama (Lecain et al. 2002; Wilsey et al. 2002), whereas heavy grazing significantly decreased the net photosynthetic rate over typical steppes in Inner Mongolia (Chen et al. 2005). Cattle grazing reduced the CO2 uptake by 37 % on a seasonal scale over a typical subalpine grassland in Central Switzerland (Rogiers et al. 2005). In addition, grazing exclusion for 3 years significantly increased the NEE by 47.37 % and ecosystem respiration (RE) by 33.14 % compared with the free-range grazing treatment in a meadow grassland on the Tibetan Plateau (Chen et al. 2014). By contrast, the grazed site always had a higher daily uptake rate or lower release rate than the ungrazed site over L. chinensis steppe in Inner Mongolia (Wang et al. 2011).
In the present study, which was supported by the Sino-German cooperation project MAGIM (Ketzer et al. 2008), the eddy covariance technique was used to measure NEE over four L. chinensis steppes under different grazing conditions to investigate the effects of different grazing intensities on the CO2 flux. We also determined the model parameters for ecosystem photosynthesis and respiration, as well as examining the response of NEE to meteorological variables and the leaf area index (LAI).
2 Site description and methods
2.1 Site description
The study sites are located at the Inner Mongolian Grassland Ecosystem Research Station (latitude: 43.55° N, longitude: 116.67° E, altitude: 1200 m), which experiences a semi-arid continental temperate climate, with wet hot summers and dry cold winters. The annual average temperature and annual precipitation are 2.6 °C and 286.6 mm, respectively (1971–2000, climate data obtained from the meteorology station at Xilinhot). Eighty-eight percent of the precipitation is concentrated in the growing seasons (May to September). The soil is dark chestnut (Mollisol) (100–150 cm depth) without a distinct CaCO3 layer (Wang and Cai 1988), which is composed of 62.0 % sand, 22.2 % silt, and 15.8 % clay. The bulk density at the surface (0–4 cm) is 1.14 ± 0.02 g cm−3 (Zhao et al. 2011).
The sites include four L. chinensis steppes, i.e., ungrazed since 1979 (UG79), winter grazed (WG), continuously grazed (CG), and heavily grazed (HG), which are all dominated by L. chinensis, Stipa grandis, Koeleria cristata, and Agropyron cristatum (Xiao et al. 1997). The UG79 site was lightly grazed by sheep before 1979, but then fenced and ungrazed after 1979. WG was grazed from mid-October until mid-February by 0.5 sheep units/ha. CG was grazed continuously by 1.2 sheep units/ha. HG was grazed by at least 2 sheep units/ha for many years (Ketzer et al. 2008). The maximum grass heights at UG79 in 2005 and 2006 were 26.3 and 27.8 cm, respectively, and UG79 always had taller grass than the grazed site (Table 1). UG79 has a gentle slope of less than 3°, whereas WG, CG, and HG are relatively flat. UG79, WG, and CG are located several hundred meters apart, whereas HG is ca 3 km to the north of UG79.
2.2 Measurements
The eddy covariance system was used to measure the CO2 flux, which is composed of a three-dimensional sonic anemometer (CSAT3, Campbell Scientific, USA) and an Open Path CO2/H2O Gas Analyzer (LI-7500, LI-COR, USA). The instruments were mounted at a height of 2 m above the ground and about 0.1 m apart. Net radiation and photosynthetic active radiation (PAR) were measured at a height of 1.5 m using a net radiometer (CNR1, KIPP&ZONEN, Netherlands) and a PAR quantum sensor (LI190SB, LI-COR, USA), respectively. The soil heat flux was measured with two heat flux plates, which were installed about 2 cm below the ground (HFP01, Hukseflux, Netherlands), and the distance between them was not more than 1.5 m. Air temperature and relative humidity were measured with temperature and relative humidity probes located near the eddy covariance system (HMP45C, Campbell Scientific, USA). Precipitation was measured (52202 Tipping Bucket Rain Gauge, RM Young, USA) at a height of 1 m. A CR5000 data logger (Campbell Scientific, USA) was used to record the data. The soil moisture at a depth of 5 cm depth was measured using horizontally inserted Theta-probes (Type ML2x, Delta-T Devices Ltd., Cambridge, UK), and the data were recorded by an automatic Data Logger (DL2e Data Logger, Delta-T Devices Ltd., Cambridge, UK) (Zhao et al. 2011). LAI was sampled at each site by clipping 5–8 representative 0.08 m2 leaf quadrants. The leaf area was measured using a portable area meter (LI-3000A) and a transparent belt conveyer (LI-3050A, LI-COR, USA) (Fan et al. 2009).
Two flux towers were used in this study. One tower was installed permanently at UG79 from May 15, 2005, until December 2008. Another tower was installed at one of the three grazed sites during 2005 and 2006 (WG, CG, and HG). This tower was moved between the different grazing treatment sites about every around 6 weeks. The detailed measurement scheme is shown in Table 1. Due to the poor data quality of the data obtained during the winter time, only data of the growing seasons were analyzed in this study.
2.3 Date processing
EdiRe software was used to calculate half-hourly CO2 fluxes from 10 Hz raw data (http://www.geos.ed.ac.uk/abs/research/micromet/EdiRe/). Several correction procedures were performed to obtain accurate energy fluxes. After coordinate rotation (double rotation as described by Kaimal and Finnigan 1994), correction of spectral loss (Moore 1986) was necessary. Next, the Webb correction (Webb et al. 1980) was applied to obtain the corrected CO2 flux. Based on the footprint analysis method proposed by Kormann and Meixner (2001), the 90 % flux fetch was below 170 m at WG, CG, and HG. The 90 % flux fetch was about 125 m at UG79 under unstable conditions and within the fenced steppe area.
Data gaps are unavoidable in long-term flux measurements due to sensor malfunctions, precipitation, dew, dust, and other factors. After quality control procedures (including stationary and integral turbulence characteristics tests, described Foken and Wichura 1996), roughly 25 % of the fluxes were missing or discarded. Linear interpolation was used for short data gaps (<2 h), and the lookup table method was used for longer gaps (Falge et al. 2001).
The relationship between the nighttime NEE (RE, μmol CO2 m−2 s−1) and soil temperature at a depth of 5 cm (T s, °C) is described by Van’t Hoff equation (Aires et al. 2008):
where a and b are the regression parameters. The temperature sensitivity coefficient (Q 10) of RE is determined by the following equation.
The partitioning of NEE into gross primary production (GPP) and RE is based on an assumption that the temperature sensitivity of RE is consistent in the nighttime and daytime (Falge et al. 2001). Thus, the nighttime RE value was extrapolated to the daytime to calculate the daytime RE. The daily RE was the sum of the daytime and nighttime RE. The daily GPP was calculated as follows.
The Michaelis-Menten model was used to evaluate the relationship between daytime NEE and PAR (Falge et al. 2001):
where α is the apparent quantum yield (μmol CO2 μmol−1 photons), NEEsat is the NEE at a saturating light level, and RE is the bulk estimated RE.
3 Results and discussions
3.1 Weather conditions and LAI
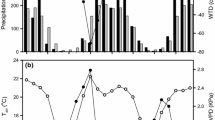
The mean air temperatures in the four growing season from 2005 to 2008 were 15.7, 15.3, 16.7, and 15.0 °C, respectively. The minimum daily mean air temperature was −0.1 °C due to a snowfall event in September 2006 and the maximum value was 28.5 °C in July 2008. The mean vapor pressure deficit (VPD) values in the growing seasons from 2006 to 2008 were 0.93, 1.15, and 0.90 kPa, but the daily mean VPD exhibited a large variation, with a maximum value of 2.56 kPa (Fig. 1a).

The daily mean (a) air temperature (T a ), vapor pressure deficit (VPD), (b) soil water content (SWC), and the daily total precipitation (PPT) at UG79 during the growing seasons (May to September) from 2005 to 2008
The precipitation rates in the four growing seasons from 2005 to 2008 were 146.9, 228.4, 177.5, and 297.1 mm, respectively. The 2008 growing season was a normal year because the precipitation of 297.1 mm was comparable to the mean precipitation during 1982–2004 (data from IMGERS), whereas the 2005, 2006, and 2007 growing seasons were dry, with 48, 19, and 37 % reductions in precipitation, respectively. In the four growing seasons, 24 % of days had precipitation events with more than 1 mm day−1. Among the rainy days, the days with more than 5 mm day−1 precipitation only accounted for 9 %. The soil water content (SWC) at a depth of 5 cm was also low during the growing season due to low precipitation. During the growing seasons, the SWC varied between 0.041 and 0.273 m3 m−3. High SWC values always followed rainy days (Fig. 1b).
The LAI increased from May and peaked during July and August. The exception was the maximum LAI at the UG79 site during 2007, which occurred in September. The difference between UG79 and WG was small in 2005 and 2006, while heavy grazing significantly reduced the LAI. The 2005 growing season was drier than 2006 but the LAI at HG was larger in 2005 than that in 2006, which was possibly due to the higher grazing intensity in 2006 (Fig. 2).

The leaf area index (LAI) at UG79, WG, and HG during the growing seasons (May to September) from 2005 to 2008
3.2 Response of the half-hourly CO2 flux to meteorological variables at UG79
Based on the half-hourly data at UG79, the relationship between the daytime NEE and PAR during the growing season was described by the Michaelis-Menten model. The stomatal regulation of photosynthesis during periods of water stress is well documented (Fu et al. 2006; Zhang et al. 2007). At this observational site, previous study found that soil water stress decreased water evaporation significantly only when SWC was higher than 0.12 m3 m−3 (Wang et al. 2012). Thus, the NEE data were grouped into two classes: SWC ≥ 0.12 m3 m−3 and SWC < 0.12 m3 m−3. Under both soil water conditions, NEE decreased (a more negative NEE represents more CO2 uptake and the minimum NEE represents the largest CO2 uptake, which is the same for NEEsat) with PAR until it reached a critical value (PARcri), after which it declined. The PARcri values were usually 900 and 1000 μmol m−2 s−1 with dry and wet soil conditions, respectively. PARcri increased to 1300 μmol m−2 s−1 in the wet year of 2008. In the present study, PARcri was lower than that determined in other grassland ecosystems (1000 to 1200 μmol m−2 s−1 for a typical steppe; 1600 μmol m−2 s−1 for a desert steppe) (Yang et al. 2011; Zhang et al. 2007).
The Michaelis-Menten model was only used when PAR was less than PARcri. The light-response curves had clear seasonal variations (Fig. 3a). In 2006, the minimum NEEsat on a monthly scale occurred during the peak growing stage (July, −8.94 μmol m−2 s−1) with the maximum LAI, but NEEsat (−3.37 μmol m−2 s−1) was depressed in August 2006 by drought and it was only 39 % of that in July. In 2006, this NEEsat seasonality and the drought effect were similar to the other 3 years.

The relationship between the half-hourly NEE and PAR at UG79 a from May to September in 2006, b for the growing season from 2005 to 2008. The relationship between the half-hourly NEE and air temperature (T a) at UG79 c from May to September in 2006, d for the growing season from 2005 to 2008. The relationship between the RE and soil temperature (T s) at UG79 e from May to September in 2006, f for the growing season in 2005 and 2006 when SWC < 0.12 m3 m−3 and SWC > 0.12 m3 m−3 and for the 2006 growing season. The dependent and independent variables were averaged with a, b PAR bins of 100 μmol m−2 s−1, c, d T a bins of 1 °C, and e, f T s bins of 1 °C
NEEsat ranged from −4.57 to −7.50 μmol m−2 s−1 during the entire growing season from 2005 to 2008, where the maximum value was in the dry year of 2005 and the minimum value was in the wet year of 2008 (Fig. 3b, Table 2). NEEsat reached −6.99 to −13.09 μmol m−2 s−1 with high SWC values, whereas low SWC values increased NEEsat by 47 to 65 % compared with the good water conditions. The effect of SWC was the same for the parameter α (Table 3). This phenomenon was consistent with that in other previously examined semi-arid typical and desert steppes, where the drought conditions depressed the photosynthetic parameters (NEEsat and α) (Li et al. 2005; Yang et al. 2011; Zhang et al. 2007).
In 2006, the optimal air temperature (T a) for the minimum half-hourly NEE at UG79 was highest in July 2006 (22 °C) and lowest in September due to grass senescence (14 °C). The optimal T a decreased to 16 °C in August 2006 due to the dry soil conditions (Fig. 3c). For the whole growing seasons, the value ranged between 17 and 20 °C from 2005 to 2008 (Fig. 3d). NEE started to increase above 20 °C over this L. chinensis steppe. The optimal value for T a was within the range reported previously for a Duolun Stipa kryroii steppe (15 to 25 °C) (Zhang et al. 2007), but significantly lower than that for a Mongolian steppe (24 °C) (Li et al. 2005).
The RE data at UG79 were selected when u* was greater than 0.2 m s−1. It is well documented that RE increases as an exponential function of the soil temperature (Falge et al. 2001). The RE-T s curve had a clear seasonal variation due to plant growth (Fig. 3e). In addition to the soil temperature and canopy development, RE was also strongly limited by soil water stress during August 2006 (mean SWC = 0.08 m3 m−3, Fig. 3e), which was similar to that found in a temperate grassland in the humid area of Germany (Hussain et al. 2011). The correlation between the low SWC and high soil temperature resulted in a decrease in RE when the soil temperature exceeded 17 °C for the entire growing season in 2006 (Fig. 3f). Thus, the RE data for each growing season were grouped into two SWC classes.
The temperature sensitivity of respiration can be expressed by Q 10, which has been used widely to calculate RE in models (Hunt et al. 1996; Potter et al. 1993; Tian et al. 1999). In this study, Q 10 ranged from 2.0 to 3.1 (Table 3), which is within the normal range for grasslands (1.2 to 3.7) (Aires et al. 2008; Falge et al. 2001; Flanagan and Johnson, 2005; Fu et al. 2006; Wang et al. 2008; Xu and Baldocchi, 2004; Zhang et al. 2007). The results were similar to the Q 10 values determined over the Duolun S. kryroii steppe (2.29 and 3.31) (Zhang et al. 2007), but they were much higher than the values calculated over a L. chinensis steppe (1.2 and 0.2) and a desert steppe in Inner Mongolia (1.6) (Hao et al. 2008; Yang et al. 2011). We found that the value of Q 10 increased with SWC at this observational site (Table 3), which was consistent with the results obtained for a Canadian mixed grassland and temperate steppes (Flanagan and Johnson, 2005; Hao et al. 2008; Zhang et al. 2007). However, our results differed from those for a Stipa krylovii steppe and a desert steppe, where Q 10 was highest at medium SWC compared with low or high SWC conditions (Wang et al. 2008; Yang et al. 2011). This discrepancy may be related to the management history of the sites. This study site had been fenced since 1979. Microbial carbon in the surface soil and labile carbon were significantly higher compared with the grazed steppe (Ma et al. 2005), and large amounts of aboveground litter accumulated. Soil respiration, which contributes up to 90 % of RE (Dugas et al. 1999), and the decomposition of accumulated litter both increased with the SWC, as well as their temperature sensitivities.
3.3 Diurnal variation in CO2 fluxes
The data in 2006 at UG79 illustrated the monthly average diurnal variations in the CO2 flux (Fig. 4a). In May, the canopy absorbed CO2 only weakly (less than −0.56 μmol m−2 s−1) during the daytime. As the canopy developed, photosynthesis and respiration were enhanced significantly in June (NEE −3.53 μmol m−2 s−1) and they peaked in July. The uptake of CO2 was depressed by drought in August. During the late growing season, the magnitude of the CO2 flux declined in the senescent stage of canopy growth. The maximum CO2 uptake and release for the monthly average diurnal cycle were −7.4 and 3.4 μmol m−2 s−1 (NEE, where a negative value indicates CO2 uptake), respectively, where both occurred in August 2008. The maximum CO2 uptake was larger than that determined over a S. kryroii steppe (−6.6 μmol m−2 s−1, Zhang et al. 2007) and a desert steppe (−3.1 μmol m−2 s−1, Yang et al. 2011) in Inner Mongolia, as well as over a grazed steppe in central Mongolia (−3.6 μmol m−2 s−1, Li et al. 2005) and a tussock grassland in New Zealand during summer drought (−4.9 μmol m−2 s−1, Hunt et al. 2002), but lower than that over C4 prairie grassland ecosystems in North America (−28.0 μmol m−2 s−1, Dugas et al. 1999; −23.0 μmol m−2 s−1, Ham and Knapp 1998; −32 μmol m−2 s−1, Suyker and Verma 2001).

a The monthly average diurnal variations in NEE at UG79 from May to September in 2006 and for August 2008. b The average diurnal variations in NEE at UG79 for the growing season from 2005 to 2008
The half-hourly NEE generally decreased as PAR increased. PAR was largest at noon, but the half-hourly NEE on the diurnal cycle was lower in the morning than in the afternoon (Fig. 4). This depression of the daytime NEE at noon is a common phenomenon in semi-arid grasslands (Fu et al. 2006; Li et al. 2005; Zhang et al. 2007), which is related to the increase in RE with T s. Another explanation is that CO2 assimilation is limited by the low enzyme activity at high temperatures and stomatal closure at high VPD (Zhang et al. 2007). In general, high T a and high VPD are always coupled together in semi-arid environments. The survival strategy of plants induces stomatal closure under high VPD and in high evaporation demand conditions (Chaves et al. 2002). Stomatal closure is more sensitive to an increase in VPD under water-stressed conditions because the supply of water from the soil to plants is deficient and leaf water potential declines (Jones 1998; Tardieu and Simonneau 1998). Thus, the depression of the daytime NEE at noon was stronger during the dry August in 2006 compared with that in the relatively wet August of 2008 (Fig. 4a).
3.4 Growing season variations in the daily CO2 flux
We detected significant seasonal variations in NEE, GPP, and RE over this steppe (Fig. 5). In the four growing seasons from 2005 to 2008, NEE became negative on May 17, 1, 18, and 9, respectively. The NEE then decreased significantly in June and reached its minimum daily value in June (−3.5 g C m−2 day−1 for 2007), July (−2.1 g C m−2 day−1 for 2006; −3.4 g C m−2 day−1 for 2008), or August (−1.5 g C m−2 day−1 for 2005). The minimum NEE was smaller than that for a Mongolian grazed steppe (−2.3 g C m−2 day−1, Li et al. 2005) and a desert steppe in Inner Mongolia (−1.6 g C m−2 d−1, Yang et al. 2011).

The daily NEE, GPP, and RE during the growing seasons from 2005 to 2008
The LAI peaked in August during each growing season, but the maximum CO2 uptake on a monthly scale occurred during July in 2006 and June in 2007. This indicates that in addition to the growth of vegetation, the seasonal variation in NEE was also regulated by the distribution of precipitation and the SWC. In August 2006, the total precipitation and mean SWC were only 17.0 mm and 0.08 m3 m−3, respectively (30-year average precipitation =70.2 mm). During this drought period, both GPP and RE decreased greatly, and GPP became smaller than RE. Thus, the ecosystem changed from being a carbon sink into a carbon source, before returning to a carbon sink in September. This summer drought caused a weak release of CO2 in August 2006. Similar responses of the CO2 fluxes to summer droughts were also observed in the other 3 years.
The duration of net CO2 assimilation differed slightly (below 10 %) among the four years, i.e., 92, 95, 95, and 101 days from 2005 to 2008, respectively. However, the magnitudes of the daily NEE, GPP, and RE were much smaller in 2005 compared with those in the other 3 years. For comparison of the total NEE for the growing seasons, the CO2 flux from 1 to 15 May 2005 was filled by the data from the same period in 2006. The cumulative NEE differed significantly for the growing seasons from 2005 to 2008, where the maximum value was −32.0 g C m−2 in the driest year of 2005 and the minimum value was −89.8 g C m−2 in the relatively wet year of 2008 (Fig. 6, Table 2). The total precipitation was higher in 2006 than in 2007, but the total NEE was smaller in 2007 (−66.1 g C m−2) than that in 2006 (−41.5 g C m−2). This difference was primarily due to the more frequent drought spells in 2006, especially the droughts in June and August of 2006 (Fig. 1).

The cumulative NEE, GPP, and RE for the four growing seasons from 2005 to 2008
3.5 Effects of grazing on the CO2 flux
The effects of grazing on the NEE depend on how grazing influences the different processes that control the GPP and RE. First, the photosynthesis parameter NEEsat was derived from the half-hourly NEE data between UG79 and the grazed sites. This comparison focused on the peak growth stage (HG in 2005 and WG in 2006). The dependence of the daytime NEE on PAR and T a was clearly affected by HG and WG. With the same PAR or T a, the NEE was lower at UG79 than that at HG and WG (Fig. 7a, b). Consequently, the CO2 uptake rate was higher in the daytime at UG79 compared with that at HG and WG (Fig. 7c). The large difference in the NEE occurred between UG79 and WG, but their difference in the LAI was small due to the non-grazing at WG during the growing season (Fig. 2). This means that the NEE difference was also attributable to other factors, including the significantly increased bulk density and the decreased organic carbon and total nitrogen of topsoils with grazing intensity (Steffens et al. 2008).

The relationship between a the daytime half-hourly NEE and PAR, b the daytime half-hourly NEE and T a, and c the average diurnal variation in the NEE at UG79, HG, and WG in 2005 and 2006. The periods are from July 10 to August 19 in 2005 and from July 1 to August 11 in 2006. The dependent and independent variables were averaged with a PAR bins of 100 μmol m−2 s−1 and b T a bins of 1 °C
In 2006, the NEEsat at WG was 59 % of that at UG79, and the ratio of HG relative to UG79 decreased to 53 % in 2005 (Table 1). However, when drought depressed the NEEsat during the late growing season in 2006, NEEsat was slightly lower at WG than at UG79 (Table 1), which was possibly due to compensatory plant growth at WG (Lecain et al. 2000; Milchunas and Lauenroth 1993). Second, WG and HG significantly reduced Q 10, whereas CG had little effect on Q 10. However, an exception occurred where the Q 10 value at HG was higher than that at UG79 during the rapid growth stage (June) in 2006 (Table 1).
On a daily scale, there was a distinct difference in the NEE between UG79 and HG, whereas the NEE was similar at UG79 and CG (Fig. 8). On a seasonal time scale, the effects of grazing on the CO2 fluxes varied according to the differences in the grazing intensities, soil moisture, and vegetation growth conditions (Table 1). In general, GPP was reduced greatly by HG (35 and 45 %) and WG (34 %) due to the reduction in the LAI. However, there was no significant difference (below 5 %) in the GPP between UG79 and HG in the late growing season of 2006 because GPP was depressed by the dry soil water conditions at UG79. In the peak stage of vegetation growth, HG reduced RE by 28 % because the shorter canopy respired less CO2 (Wilsey et al. 2002) and the increased soil bulk density due to trampling by sheep decreased RE (Binkley et al. 2003), although the RE at HG was slightly higher than that at UG79 (below 10 %) during the early and late growing seasons. A possible explanation is that both the higher soil temperature and respiration from sheep contributed to RE at HG, and this was more important than the difference in the LAI between UG79 and HG. These negative impacts on RE and Q 10 are consistent with the results obtained for a tropical pasture in Panama (Wilsey et al. 2002), but different from studies of a subalpine grassland in the Swiss Alps and salt marshes in Denmark, where no difference in the RE was found (Morris and Jensen 1998; Rogiers et al. 2005).

The daily NEE at UG79, WG, CG, and HG during the growing seasons in 2005 and 2006
The effects of grazing on the NEE increased with the grazing intensity. HG changed the steppe from a CO2 sink into a CO2 source. When the grazing intensity decreased, WG weakened the CO2 uptake rate greatly by 79 % in 2006. In particular, during the early growing season, CG benefited photosynthesis and respiration in the ecosystem, and the NEE was similar at UG79 and CG. The impact of grazing on the NEE varied among different ecosystems. In this study, the negative effects of grazing on the NEE were consistent with the results reported for a meadow grassland on the Tibetan Plateau, a subalpine grassland in the Swiss Alps, and a Mediterranean grassland (Aires et al. 2008; Chen et al. 2014; Rogiers et al. 2005), whereas they differed completely from the effects of grazing on the NEE over the L. chinensis steppe in Inner Mongolia (Wang et al. 2011). This difference may be explained partly by a compensatory effect at the grazed site, which had more leaves with a greater leaf-level photosynthetic capacity and an open canopy architecture with a higher radiation use efficiency (Wang and Wang 1999; Zhou et al. 2007). However, this greater photosynthetic capacity at the ecosystem level was not observed at the grazed site in the present study (Table 1). Moreover, there was no significant difference in the NEE between ungrazed and grazed sites over a shortgrass steppe in the USA and tropical pasture in Panama (Risch and Frank 2006; Wilsey et al. 2002). Risch and Frank (2006) hypothesized that moisture stress reduced the effect of grazing in the Yellowstone Park. This was also the case for the GPP at this study site during the late growing season in 2006.
3.6 Factors controlling the CO2 flux on a daily scale
On a daily scale, both the GPP and RE increased linearly with the SWC. The response of the GPP to SWC was more sensitive than that for RE over this steppe (Fig. 9a), which was consistent with the results over a desert steppe in Inner Mongolia (Yang and Zhou 2013). By contrast, Reichstein et al. (2002) found that the RE was limited by drought more than GPP in two Mediterranean forests because most of the respiration occurred in the litter and upper soil layers, which dried first, whereas the leaves could be recharged with water acquired by the roots in the deeper soil layers to support photosynthesis. In semi-arid grasslands, the roots of grasses are distributed mainly in the soil layers located at less than 30 cm (Ping et al. 2010). Thus, the recharge of grasses with water from deeper soil layers is very limited, and the GPP is more restricted by drought than RE.

a The relationship between the GPP, RE, and SWC in peak growth stage (July and August) from 2005 to 2008 at UG79 (rainy days were excluded). b The relationship between the GPP and RE for the peak growth (July and August) and the early and late growth (June and September) stages from 2005 to 2008 at UG79
In addition to the soil temperature and soil moisture levels, RE also responded to the canopy productivity (Fig. 9b), which produced the available substrate for respiration (Thomey et al. 2011; Yan et al. 2011). The RE had a linear correlation with the GPP over this steppe. The daily RE accounted for more of the daily GPP in the peak growth stage (RE = 0.52GPP + 0.73, R 2 = 0.75) compared with the early and late growth stages (RE = 0.38GPP + 0.82, R 2 = 0.73) (Fig. 9b). This strong relationship between the RE and GPP on daily, seasonal, or annual scales has also been documented in Mediterranean C3/C4 grasslands (Aires et al. 2008; Xu and Baldocchi 2004), an oak/grass savanna (Ma et al. 2007), semi-arid steppes (Li et al. 2005; Yang and Zhou 2013), and a degraded grassland (Du and Liu 2013).
The monthly total NEE varied linearly with the monthly mean LAI (Fig. 10a). The slope of the linear relationship at this site was similar to that for a Mediterranean C3/C4 grassland, although the correlation was weaker (R 2 = 0.77, slope = −1.71) (Aires et al. 2008). The change in the LAI explained 46 % of the variation in the NEE, and the remaining percentage was associated with the changes in the PAR, SWC, and other meteorological variables. For example, with a similar LAI at UG79, the mean daily NEE were 0.59 and −0.67 g C m−2 day−1 in July and September 2008, respectively. This large difference in the NEE is partly attributable to the changes in the SWC, as shown in Fig. 10b.

a The relationship between the NEE and LAI; the NEE was the monthly average value before and after the LAI sampling day; b The relationship between the NEE and SWC on a daily scale during July and September, 2006, at UG79
4 Conclusion
The ecosystem photosynthesis and respiration parameters (NEEsat, α, and Q 10) were investigated in different years and soil water conditions at four differently grazed sites. The daily and total NEE indicated that both the amounts and distribution of precipitation affected the growing season CO2 fluxes. By altering the light-response curve, grazing (WG and HG) significantly reduced the GPP. In the peak growth stage, WG and HG reduced RE and Q 10, where HG even changed the CO2 sink into a CO2 source. However, the NEE was similar at UG79 and CG during the early growing season. The negative effects of grazing on the CO2 fluxes increased with the grazing intensity, but the effects also varied with the growth stages of vegetation. Thus, understanding the effects of grazing on the total CO2 fluxes during the growing season and the whole year still requires more measurements.
References
Aires LM, Pio CA, Pereira JS (2008) Carbon dioxide exchange above a Mediterranean C3/C4 grassland during two climatologically contrasting years. Glob Chang Biol 14:539–555
Bai YF, Han XG, Wu JG, Chen ZZ, Li LH (2004) Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 431:181–184
Binkley D, Singer F, Kaye M, Rochelle R (2003) Influence of elk grazing on soil properties in Rocky Mountain National Park. For Ecol Manag 185:239–247
Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CPP, Osorio ML, Carvalho I, Fraia T, Pinheiro C (2002) How plants cope with water stress in the field? Photosynthesis and growth. Ann Bot 89:907–916
Chen J, Shi WY, Cao JJ (2014) Effects of grazing on ecosystem CO2 exchange in a meadow grassland on the Tibetan Plateau during the growing season. Environ Manag. doi:10.1007/s00267-014-0390-z
Chen SP, Bai YF, Lin GH, Liang Y, Han XG (2005) Effects of grazing on photosynthetic characteristics of major steppe species in the Xilin River basin, Inner Mongolia, China. Photosynthetica 43(4):559–565
DAHV (Department of Animal Husbandry and Veterinary, Institute of Grasslands, Chinese Academy of Agricultural Sciences), CISNR (Commission for Integrated Survey of Natural Resources, Chinese Academy of Sciences) (1996) Rangeland resources of China. China Agricultural Science and Technology, Beijing
Du Q, Liu HZ (2013) Seven years of carbon dioxide exchange over a degraded grassland and a cropland with maize ecosystems in a semiarid area of China. Agric Ecosyst Environ 173:1–12
Dugas WA, Heuer ML, Mayeux HS (1999) Carbon dioxide fluxes over Bermuda-grass, native prairie, and sorghum. Agric For Meteorol 93:121–139
Falge E, Baldocchi D, Olson R (2001) Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric For Meteorol 107:43–69
Fan L, Gao Y, Brück H, Bernhofer C (2009) Investigating the relationship between NDVI and LAI in semi-arid grassland in Inner Mongolia using in-situ measurements. Theor Appl Climatol 95:151–156
Flanagan LB, Johnson BG (2005) Interacting effects of temperature, soil moisture and plant biomass production on ecosystem respiration in a northern temperate grassland. Agric For Meteorol 130:237–253
Foken T, Wichura B (1996) Tools for quality assessment of surface-based flux measurements. Agric For Meteorol 78:83–105
Fu Y, Zheng Z, Yu G, Hu Z, Sun X, Shi P, Wang Y, Zhao X (2006) Depression of net ecosystem CO2 exchange in semi-arid leymus chinensis steppe and alpine shrub. Agric For Meteorol 137:234–244
Gilmanov TG, Soussana JF, Aires L (2007) Partitioning European grassland net ecosystem CO2 exchange into gross primary productivity and ecosystem respiration using light response function analysis. Agric Ecosyst Environ 121:93–120
Ham JM, Knapp AK (1998) Fluxes of CO2, water vapor, and energy from a prairie ecosystem during the seasonal transition from carbon sink to carbon source. Agric For Meteorol 89:1–14
Hao Y, Wang Y, Mei X, Huang X, Cui X, Zhou X, Niu H (2008) CO2, H2O and energy exchange of an Inner Mongolia steppe ecosystem during a dry and wet year. Acta Oecologia 33:133–143
Hunt ER, Piper SC, Nemani R, Keeling CD, Otto RD, Running SW (1996) Global net carbon exchange and intra-annual atmospheric CO2 concentrations predicted by an ecosystem process model and three-dimensional atmospheric transport model. Glob. Biogeochem. Cycle 10:431–456
Hunt JE, Kelliher FM, McSeveny TM, Byers JN (2002) Evaporation and carbon dioxide exchange between the atmosphere and a tussock grassland during a summer drought. Agric For Meteorol 111:65–82
Hussain MZ, Grünwaldb T, Tenhunen JD, Li YL, Mirzae H, Bernhofer C, Otieno D, Dinh NQ, Schmidt M, Wartinger M, Owen K (2011) Summer drought influence on CO2 and water fluxes of extensively managed grassland in Germany. Agric Ecosyst Environ 141:67–76
Jones HG (1998) Stomatal control of photosynthesis and transpiration. J Exp Bot 49:387–398
Kaimal JC, Finnigan JJ (1994) Atmospheric boundary layer flows: Their structure and measurement. p. 289. Oxford University Press, New York
Ketzer B, Liu H, Bernhofer C (2008) Surface characteristics of grasslands in Inner Mongolia as detected by micrometeorological measurements. Int J Biometeorol 52:563–574
Kormann R, Meixner FX (2001) An analytic footprint model for neutral stratification. Boundary Layer Meteorology 99:207–204
LeCain DR, Morgan JA, Schuman GE, Reeder JD, Hart RH (2000) Carbon exchange rates in grazed and ungrazed pastures of Wyoming. J Range Manag 53:199–206
LeCain DR, Morgan JA, Schuman GE, Reeder JD, Hart RH (2002) Carbon exchange and species composition of grazed pastures and exclosures in the shortgrass steppe of Colorado. Agriculture Ecosystems and Environment 93:421–435
Li SG, Asanuma J, Eugster W, Kotani A, Liu JJ, Urano T, Oikawa T, Davaa G, Oyunbaatar D, Sugita M (2005) Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia. Glob Chang Biol 11:1941–1955
Ma S, Baldocchi D, Xu L, Hehn T (2007) Inter-annual variability in carbondioxide exchange of an oak/grass savanna and open grassland in California. Agric For Meteorol 147:157–171
Ma XZ, Wang YF, Wang SP, Li CS (2005) Impacts of grazing on soil carbon functions in the grassland of xilin river basin, Inner Mongolia. Acta Phytoecol. Sin. 29:569–576
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366
Moore CJ (1986) Frequency response corrections for eddy correlation systems. Boundary Layer Meteorology 37:17–35
Morris JT, Jensen A (1998) The carbon balance of grazed and nongrazed Spartina anglica saltmarshes at skallingen, Denmark. J Ecol 86:229–242
Parton WJ, Scurlock JMO, Ojima DS, Schimel DS, Hall DO (1995) Impact of climate change on grassland production and soil carbon worldwide. Glob Chang Biol 1:13–22
Ping X, Zhou G, Zhuang Q, Wang Y, Zuo W, Shi G, Lin X, Wang Y (2010) Effects of sample size and position from monolith and core methods on the estimation of total root biomass in a temperate grassland ecosystem in Inner Mongolia. Geoderma 155:262–268
Potter CS, Randerson JT, Field CB, Matson PA, Vitousek PM, Mooney HA, Klooster SA (1993) Terrestrial ecosystem production: a process model based on global satellite and surface data. Glob. Biogeochem. Cycle 7:811–841
Reichstein M, Tenhunen JD, Roupsard O, Ourcival JM, Rambal S, Miglietta F, Peressotti A, Pecchiari M, Tirone G, Valentini R (2002) Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: revision of current hypotheses? Glob Chang Biol 8:999–1017
Risch AC, Frank DA (2006) Carbon dioxide fluxes in a spatially and temporally heterogeneous temperate grassland. Oecologia 147:291–302
Rogiers N, Eugster W, Furger M, Siegwolf R (2005) Effect of land management on ecosystem carbon fluxes at a subalpine grassland site in the Swiss alps. Theor Appl Climatol 80:187–203
Schaffrath D, Barthold FK, Bernhofer C (2011) Spatiotemporal variability of grassland vegetation cover in a catchment in Inner Mongolia, China, derived from MODIS data products. Plant Soil 340:181–198
Steffens M, Kölbl A, Totsche KU, Kögel-Knabner I (2008) Grazing effects on soil chemical and physical properties in a semiarid steppe of Inner Mongolia (P.R. China). Geoderma 143:63–72
Suyker AE, Verma SB (2001) Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie. Glob Chang Biol 7:279–289
Tardieu F, Simonneau T (1998) Variability among species of stomatal control under fluctuating soil water status and evaporative demand. Modeling Isohydric and Anisohydric Behaviors Journal of Experimental Botany 49:419–432
Tian H, Melillo JM, Kicklighter DW, McGuire AD, Helfrich J (1999) The sensitivity of terrestrial carbon storage to historical climatic variability and atmospheric CO2 in the United States. Tellus 51B:414–452
Thomey ML, Collins SL, Vargas R, Johnson JE, Brown RF, Natvig DO, Friggens MT (2011) Effect of precipitation variability on net primary production and soil respiration in a chihuahuan desert grassland. Glob Chang Biol 17:1505–1515
Wang J, Cai Y (1988) Studies of genesis, types and characteristics of the soils of the xilin river basin. In: Inner Mongolia grassland ecosystem research station (Ed.), research on grassland ecosystem 3. Science Press, Beijing, pp. 23–83
Wang L, Liu HZ, Ketzer B, Horn R, Bernhofer C (2012) Effect of grazing intensity on evapotranspiration in the semiarid grasslands of Inner Mongolia, China. J Arid Environ 83:15–24
Wang YF, Wang SP (1999) Influence of different stocking rates on aboveground present biomass and herbage quality in Inner Mongolia steppe. Acta Prataculturae Sin 8:15–20
Wang Y, Zhou G, Wang YH (2008) Environmental effects on net ecosystem CO2 exchange at half-hour and month scales over Stipa krylovii steppe in northern China. Agric For Meteorol 148:714–722
Wang YF, Cui XY, Hao YB, Mei XR, Yu GR, Huang XZ, Kang XM, Zhou XQ (2011) The fluxes of CO2 from grazed and fenced temperate steppe during two drought years on the Inner Mongolia plateau, China. Sci Total Environ 410:182–190
Webb EK, Pearman GI, Leuning R (1980) Correction of flux measurements for density effects due to heat and water vapour transfer. Q J R Meteorol Soc 106:85–100
Wilsey BJ, Parent G, Roulet NT, Moore TR, Potvin C (2002) Tropical pasture carbon cycling: relationships between C source/sink strength, above-ground biomass and grazing. Ecol Lett 5:367–376
Xiao X, Ojima D, Parton W, Bonham C (1997) Modelling of biomass and soil organic matter of leymus chinensis steppe and Stipa grandis steppe. In: Station IMGER (ed) Research on grassland ecosystem 5. pp 118–129. Science Press
Xu LK, Baldocchi D (2004) Seasonal variation in carbon dioxide exchange over a Mediterranean annual grassland in California. Agric For Meteorol 1232:79–96
Yan L, Chen S, Huang J, Lin G (2011) Water regulated effects of photosynthetic substrate supply on soil respiration in a semiarid steppe. Glob Chang Biol 17:1990–2001
Yang FL, Zhou GS, Hunt JE, Zhang F (2011) Biophysical regulation of net ecosystem carbon dioxide exchange over a temperate desert steppe in Inner Mongolia, China. Agric For Meteorol 142:318–328
Yang FL, Zhou GS (2013) Sensitivity of temperate desert steppe carbon exchange to seasonal droughts and precipitation variations in Inner Mongolia, China. PLoS One 8(2):e55418. doi:10.1371/journal.pone.0055418
Zhang WL, Chen SP, Chen J, Wei L, Han XG, Lin GH (2007) Biophysical regulations of carbon fluxes of a steppe and a cultivated cropland in semiarid Inner Mongolia. Agric For Meteorol 146:216–229
Zhao Y, Peth S, Reszkowska A, Gan L, Krümmelbein J, Peng X, Horn R (2011) Response of soil moisture and temperature to grazing intensity in a leymus chinensis steppe, Inner Mongolia. Plant Soil 340:89–102
Zhou X, Wan S, Luo Y (2007) Source components and interannual variability in soil CO2 efflux under experimental warming and clipping in a grassland ecosystem. Global Chang Biology 13:761–775
Acknowledgments
The work presented is supported by the National Natural Science Foundation of China (grant No. 41275023 and 41461144001), the Strategic Priority Research Program of Chinese Academy of Sciences under Grant XDA05110102, and the German Science Foundation within the research group For 536 MAGIM (P6). Use of data collected by IMGERS (additional meteorological data), by Yingzhi Gao (P2, LAI data) and by Zhao Ying, Gan Lei (P7, SWC data), is greatly appreciated.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Wang, L., Liu, H. & Bernhofer, C. Response of carbon dioxide exchange to grazing intensity over typical steppes in a semi-arid area of Inner Mongolia. Theor Appl Climatol 128, 719–730 (2017). https://doi.org/10.1007/s00704-016-1736-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00704-016-1736-7