
Abstract
The reproductive biology of the sympatric species Gagea lutea and Gagea spathacea was analyzed morphologically and by resource allocation measurements. Both taxa reproduce vegetatively by bulbils. The hexaploid G. lutea regularly forms seeds, but flowering plants cease to produce bulbils. Seed set was never observed in nonaploid G. spathacea which does not stop vegetative reproduction when flowering. In this species, the pollen contains a high proportion of non-viable grains. Already sterile plants invest more resources (per cent of total nitrogen) into bulbils than G. lutea (10.9 vs. 5.9%). For flowering plants, the respective values are 6.1% (flowers) plus 18.4% (bulbils) for G. spathacea versus 14.8% (flowers only) for G. lutea. G. spathacea lost the ability to reproduce sexually and relies solely on vegetative reproduction. This seems to require the breakdown of the switch mechanism between the bulbil and flower formation (as in G. lutea) and a higher net investment in reproduction, hampering individual growth.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The genus Gagea Salisb. (Star of Bethlehem, Golden Star) is one of the most diverse genera of the Liliaceae. More than 670 specific and intraspecific combinations have been published. Uphof (1958–1960) mentioned at least 90 valid species, but Levichev (1999a) listed many more, and his most recent list comprises over 280 species (Levichev 2008), most of them described from Central Asia. Eastern Asia was identified as the probable centre of origin for the Tulipa–Erythronium–Gagea clade (Patterson and Givnish 2002). For Germany, seven species are accepted (Rothmaler et al. 2005: p. 783). In addition, with G. pomeranica Ruthe and G. megapolitana Henker, two taxa of putative hybridogeneous origin were described (see Peterson et al. 2004, 2008a; Henker 2005; Peruzzi 2008b). All German species have a similar life history and are sometimes difficult to distinguish in non-flowering condition, since some traits useful for species differentiation are hidden in the subterraneous organs. A one-leaved parent bulb stores resources nourishing the current-year plant and grows a replacement bulb to support the plant in the next year, plus one to many daughter bulbs (furthermore called bulbils) whose arrangements differ significantly between species (Levichev 1999b). The species of Gagea belong to different ploidy levels, with x = 12 as the basic (haploid) chromosome number for the genus (Peruzzi 2003, 2008a; Henker 2005; Peruzzi and Aquaro 2005; Rothmaler et al. 2005).
As already stated by Irmisch (1850) and observed by us as well, the German species of the genus follow a rather uniform growth pattern: in a given year, the lowermost leaf on the main shoot is the main part of the subterranean bulb and stores all resources to grow the current-year plant. The second basal leaf is the first visible one; usually the main assimilating leaf. The axillary shoot of this leaf initiates another subterranean leaf to form the replacement bulb. If this shoot elongates, the plant forms a visible replacement bulb outside the old one, otherwise the plant continues within the old sheath. Some species develop a second green leaf on the main shoot if the plant becomes stronger. In others, this leaf remains rudimentary or forms a bract around the inflorescence. Regardless, whether a second basal leaf develops or not, it can form a reduced axillary shoot with one to many daughter bulbils. In strong plants, the main shoot grows into an inflorescence with additional leaves that are gradually reduced in size. In some Central Asian species and exceptionally in G. lutea (L.) Ker Gawl., these upper leaves may develop bulbiferous axillary shoots as well. In rare cases, among the German species most often observed in G. villosa (M. Bieb.) Sweet, the inflorescence can develop bulbils instead of flowers; similar to many species of the genus Allium L. (wild onion). The axillary shoot of the second basal leaf will become the flowering shoot of the following year; the plant grows sympodially. After seed set, all epigeous parts of the plants decay, and the replacement bulb regrows a completely new above-ground plant in the following season.
According to the classification proposed by Peterson et al. (2008b), the two species studied herein, G. lutea and G. spathacea Salisb., belong to different sections of the genus and are placed distantly in the phylogenetic tree constructed using chloroplast and nuclear DNA sequences. However, the two species are often found together and show a superficially identical annual growth cycle.
In Germany, G. lutea is the most abundant species of the genus in deciduous woodlands with moderately moist soils which are not too poor in nutrients. It possesses a large European range with a closed distribution stretching from western Central Europe to the Ural Mountains and some additional outposts in northern Spain, Central Norway and western Siberia (Meusel et al. 1965; Hultén and Fries 1986). Peterson et al. (2008b) place it in the Sect. Gagea. The diploid chromosome number of this species is reported as 6x = 72 (Peruzzi 2003, 2008a; Henker 2005). The species flowers readily and can form large stands especially in mown lawns of parks or churchyards where it perceives more light than in closed woodlands. It develops only one basal leaf; the second visible leaf occurs only in flowering plants, being connate with the inflorescence to form a bract beneath the umbel.
The second species, G. spathacea, often occurs together with G. lutea, but prefers slightly moister soils. In most regions of Germany, this species is much rarer than G. lutea; and its weak tendency to flower is well-known among field botanists. Levichev (in Peterson et al. 2008b) lists this species in the monotypic sect. Spathaceae Levichev and molecular analyses confirm this isolated position (Peruzzi et al. 2008). In contrast to G. lutea, older plants develop mostly two basal leaves. Like in G. lutea, the next leaf is connate with the depauperate inflorescence, ensheathing it like a spathe. However, often whole populations consist of sterile plants only, and even large stands with thousands of plants may include only a few flowering plants. The species seems to be common only in northern Germany and southern Scandinavia (Meusel et al. 1965; Hultén and Fries 1986), but scattered occurrences are reported from numerous other European countries (e.g., Tomović and Niketić 2005; Peruzzi et al. 2008). With more than three-fourth of the contiguous world range within Central Europe, this region has a particular importance for the species’ conservation (Schnittler and Günther 1999); and among Central European countries, Germany is most responsible (Korneck et al. 1996). Chromosome counts range between 102 and 108 (Westergård 1936; Peruzzi 2003; Henker 2005), which can be explained best by a relation of 9x = 108. The species would thus be nonaploid.
This study reports investigations on the growth pattern of both species to compare (1) pollen viability, (2) seed set, (3) numbers of bulbils formed, (4) thresholds for the respective reproduction types, and (5) resources invested into generative and vegetative reproduction, respectively. If G. spathacea is indeed completely sterile, as guessed by several authors (e.g., Peterson et al. 2008b) the question arises as how the species can compensate the loss of seed production?
Materials and methods
Sampling
During end of March to beginning of May in the years 2003, 2006 and 2007, we unearthed plants of both Gagea species from several localities in Germany (Western Pomerania, vicinity of Greifswald) and Denmark (Island Moen), the soil was removed carefully and the plants and their current-year bulbils were collected as herbarium specimens. We tried to obtain a representative selection of all life stages for the two species from very-young plants showing a first, filiform leaf to old, richly flowering individuals. Plants were harvested towards the end of their flowering period to ascertain that flowers were still easy to count but newly formed bulbils were fully developed.
We used the traits described by Haeupler (1969) to distinguish young individuals of the two species, which often occur together. Even juvenile leaves of G. lutea with a diameter of less than 0.5 mm have edges (their cross-section being rather polygonal than spherical), whereas those of G. spathacea are always completely round. Both species can be told apart simply by rolling the leaves gently between thumb and index finger: the edges of G. lutea cause the leaf to roll less smoothly than in G. spathacea.
Geographical coordinates were taken with a hand-held GPS at each collecting site. Most localities are in the parks of former estates, where old-grown trees and mown lawns provide a suitable environment for large and dense populations. Table 1 lists all plants sampled for (1) morphological analyses, (2) to control seed set and (3) to estimate pollen viability with harvesting dates from the following localities:
-
1.
Germany, Mecklenburg-Western Pomerania, Greifswald, city limits, old trees around the former city wall, 54°05′38′′N, 13°22′22′′E
-
2.
Greifswald, New Cemetery W of the old city, 54°05′57′′N, 13°21′43′′E
-
3.
Forest Elisenhain 4.8 km ESE Greifswald city centre, old-growth forest in a reserve (NSG “Eldena”), 53°04′54′′N, 13°26′38′′E
-
4.
Forest Elisenhain ca. 6 km SW Greifswald, southern part near village Koitenhagen, woodland fringe, 53°03′52′′N, 13°27′08′′E
-
5.
Wampen ca. 5 km NE Greifswald, abandoned park of the estate, 54°08′02′′N, 13°25′05′′E
-
6.
Griebenow estate ca. 9.6 km WSW Greifswald, park behind the castle, 54°04′48′′N, 13°14′55′′E
-
7.
Wrangelsburg ca. 16.5 km SE Greifswald, old-growth deciduous forest at the eastern shore of a lake, NE of Wrangelsburg castle, 54°01′05′′N, 13°36′13′′E
-
8.
Behrenhoff estate ca. 10 km S Greifswald, deciduous woodland of the former park, 54°00′29′′N 13°23′23′′E
-
9.
Ranzin, ca. 19 km SSE Greifswald, old-growth deciduous forest near the estate, NW of the castle, 53°56′45′′N, 13°32′26′′E
-
10.
Denmark, Island Moen, Liselund castle, park between the castle and a lake, near the chalk cliffs of the northwestern part of the island, 55°00′01′′N, 12°31′20′′E
Morphological analysis
All organs of a plant were counted and measured in fresh condition: flowers, cauline leaves, bulbs and bulbils. Only bulbils of the current year were recorded, they are easy to recognize by their light colour. Even when not growing a green leaf, previous year bulbils seem to develop a new nutrient-storing leaf inside the bulbil, which causes the bulbil to darken in colour when the outer leaf weathers away. Using a ruler and/or a digital caliber (TCM 227579, precision ± 0.02 mm), the following measures were taken: maximum diameter of parent bulb and bulbils, and length and width (broadest part) of the (one or two) basal aboveground leaves. Leaf length was measured from the emergence point from the bulb to the tip, thus including the subterranean part. The upper leaf surface was roughly estimated according to the form of a rhombus calculated from its length and maximum width. In total, 505 plants of G. lutea and 506 of G. spathacea were investigated. For all measures, the standard deviation (SD) is given.
The size of parent bulbs (maximum circumference) was taken as a proxy for age of the plant, which can be determined only within the first years until the oldest sheaths of bulbs are completely rotten. Within the classes of 0.25 mm bulb diameter, the proportions of plants that flowered and/or had daughter bulbils were calculated. To determine a threshold for the diameter of the parent bulb needed to develop bulbils and/or flowers, these graphs were fitted against the equation y = 1/[1 + e−(x−xo)/b] which describes a sigmoid function with xo as the threshold (the turning point of its slope). For G. lutea, only the part of the curve up to the maximum productivity of daughter bulbils was considered for regression.
Resource allocation
To compare between resources used for vegetative and generative reproduction, a subsample of plants from both Gagea species was dried for 3 h at 60°C, and plants were dissected into five parts: parent bulb, bulbils, leaves, inflorescence (including bracts) and the flowers itself. For each species a cohort of 103 plants from a single locality was sampled (G. lutea: Griebenow estate, 29 April 2006; G. spathacea: Ranzin estate, 7 May 2006). Collection was timed in a way that all flowers had opened; with G. spathacea usually reaching its flowering peak nearly 2 weeks later than G. lutea. In addition, mature seeds of G. lutea were collected about 6 weeks later (11 June 2006). For each species, ten randomly chosen aliquots of the five parts were cut off or pooled in a way that their dry mass was between 10 and 15 mg. This weight corresponds with a medium-sized parent bulb, 7–15 daughter bulbils, or 2–5 flowers. Samples were wrapped into tin foil and analyzed for carbon (C) and nitrogen (N) content with an automatic analyzer Vario EL III. Using the means of ten samples, nitrogen contents for parts of the whole investigated plant cohort were calculated and used as a resource estimator (compare Ashman 1994).
For 97 plants of G. lutea (68 of these sterile, 29 flowering) and 64 plants of G. spathacea (46 of these sterile, 18 flowering) all parts of a plant (parent bulb, daughter bulbils, cauline leaves, inflorescence without flowers, and flowers itself) were separated and weighted (precision: 1 μg). For logit-log plots of dry mass of plant parts versus total dry mass as well as nitrogen content of plant parts versus total nitrogen regression analyses were performed. For these plots, the slope of the regression line equals one if the amount of resources allocated to a plant part remains constant with increasing plant size. Slopes below one indicate decreasing investment in the respective plant part, where as slopes above one indicate increasing investment.
Pollen viability and seed set
Four populations of G. lutea (9 plants) and three populations of G. spathacea (16 plants) were examined for pollen viability. Plants were transferred to a greenhouse prior to the analysis, and anthers from flowering plants were removed shortly after anthesis and pollen grains were subjected to a viability analysis as described by Kearns and Inouye (1993): Pollen grains were incubated for 10 min in a 10% saccharose solution containing a few drops of fluorescein–diacetate in acetone (2 μg/ml). Pollen grains counted as viable take the substance up and reduce it via an esterase to fluorescein, causing those cells to fluoresce in yellow-green after excitation at 480 nm. Using an Olympus IX50 reverse microscope with a U-RFL-T fluorescence lamp, for each species at least 1,200 randomly chosen pollen grains (ten fields of a 10× microscope lens) were sorted into viable (fluorescing) and non-viable pollen with analySIS 3.1 LabFlow software (Soft Imaging Systems). For an additional ten fields of a 40× lens (at least 220 grains for each species), length and width were measured; and pollen grain volumes were calculated according to the formula for an ellipsoid and given in nanoliters.
Seed set of both species was recorded in the field for plants that were individually marked at the peak of the flowering period. The number of flowers was recorded, and fruiting inflorescences were harvested before the capsules opened. At this time, the capsules were still green and fleshy. For G. lutea all non-aborted seeds, including the elaiosome, were almost fully developed, and had turned in colour from white to brown. Seed set was assessed for five populations with 217 plants (695 flowers) in G. lutea and four populations with 177 plants (268 flowers) in G. spathacea.
Results
Morphological analyses
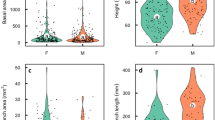
Of the two analyzed species, G. lutea develops much larger individuals. All except for three neighbouring plants developed only a single, flat basal leaf (up to 57 cm long). In young individuals, the leaf is polygonal in cross-section but flattens when plants grow; old flowering plants develop leaves up to 12 mm wide. The diameter of the parent bulb depends on leaf area (Fig. 1). The diameter of parent bulbs for the 505 plants investigated ranged from 0.76 to 13.17 mm, with a mean diameter of 6.71 ± 2.68 mm (Table 2). These values come close to a normal distribution (Kolmogorov–Smirnov-distance: 0.055). Larger sterile plants of G. lutea form 1–23 daughter bulbils (average 8.7 ± 5.3). Without any exception, flowering plants do not develop daughter bulbils any more; flowers number between 1 and 9 (mean 4.0 ± 1.8).

Relationship between leaf area (estimated as 0.5 × length × width) and diameter of the parent bulb in Gagea lutea and G. spathacea. A logistic regression according to the function log y = a log x + b with y as bulb diameter and x as leaf area resulted in a = 0.415 and b = −0.331 with R 2 = 0.744 for G. lutea and a = 0.295, b = −0.021 with R 2 = 0.578 for G. spathacea, respectively. Shown are the regression line (solid) and the 95% confidence limits (dashed line)
In contrast to G. lutea, larger vegetative plants of G. spathacea often develop a second basal aboveground leaf (129 vegetative plants with two, 166 with one leaf). Both leaves are much smaller than in G. lutea and always tubular (up to 1.9 mm in diameter, maximum length 16.0 and 17.2 cm, respectively). Almost all flowering plants (88 of 91) had two cauline leaves as well. The correlation between leaf area (here taken as the sum of the area for both leaves) and diameter of parent bulb is less strong, the regression line shows a lower slope (Fig. 1). On an average, plants form significantly smaller bulbs (range 0.79–8.30 mm, mean 3.67 ± 1.34 mm). Values for the 506 plants investigated deviate somewhat from a normal distribution (K.–S.-distance: 0.084). The main difference to G. lutea lies in reproductive behaviour: all flowering plants develop daughter bulbils (Table 2). Vegetatively reproducing plants possess 1–54 (mean 9.3 ± 9.2) daughter bulbils. Of these, sterile plants have an average of 5.6 ± 4.9 (1–31), but flowering plants 21.0 ± 10.1 (1–54) bulbils. Flowers number between one and three (mean 1.5 ± 0.6).
In both species of Gagea, the minimum diameter of the parent bulb needed to store enough resources to initiate daughter bulbils is considerably lower than that to form flowers (Fig. 2). For G. lutea, the threshold (50% of the plants developing the respective structure) is 2.63 ± 0.21 mm (R 2 = 0.902 for a regression of the increasing part of the graph with a sigmoid function, Fig. 2c). The smallest plants with a single bulbil had a bulb diameter <2 mm. However, older plants cease to grow bulbils and develop flowers, with a threshold of 6.94 ± 0.04 mm bulb diameter (R 2 = 0.998). Since a flowering plant was never seen to form bulbils in this species, the proportion of plants forming bulbils decreases as the proportion of flowering plants increases. The maximum number of bulbils was formed in the class of 5.5–6.0 mm parent bulb diameter (n = 27, 10.2 bulbils on an average, Fig. 2a). The maximum number of flowers was formed by the largest plants (13.0–13.5 mm diameter of parent bulb, n = 2, average eight flowers).

Numbers of flowers (closed circles) and daughter bulbils (open circles) produced in dependence of parent bulb diameter (classes of 0.5 mm) in Gagea lutea (a) and G. spathacea (b). Given is the mean number per diameter class and the standard error of means. Proportions of plants that developed bulbils and/or flowers in Gagea lutea (c) and G. spathacea (d) in dependence of parent bulb diameter. The increasing parts of the graphs are fitted according to the equation y = 1/[1 + e−(x−xo)/b] with xo as the threshold for the formation of the respective structure
The respective thresholds for G. spathacea are lower: 2.43 ± 0.12 mm diameter of the parent bulb for bulbil formation (R 2 = 0.977) and 5.34 ± 0.12 mm for flowers (R 2 = 0.992, Fig. 2d), respectively. Both numbers of bulbils and numbers of flowers increase with the diameter of the parent bulb, since all flowering plants of G. spathacea form bulbils as well. The maximum number of bulbils (average 33.3) was formed in the three classes >7 mm bulb diameter (n = 10, Fig. 2b). These plants developed also the maximum number of flowers observed (average 1.8).
Resource allocation
Table 3 shows carbon and nitrogen contents as well as their relation in five analyzed organs of the two species. All epigeous plant parts show significantly higher nitrogen contents than bulbs and bulbils, whereas the C-content remains roughly constant. Nitrogen contents in G. spathacea are lower than in G. lutea for all plant parts, reaching between 72% (flowers) and 80% (daughter bulbs) of the values recorded for G. lutea. In contrast, carbon contents are higher (ranging from 103% in flowers to 113% in bulbs).
For 82 plants of G. lutea and 64 plants of G. spathacea, all plant parts were weighted and total nitrogen content was calculated using the contents per milligram dry tissue (Fig. 3). In all cases, leaves and assimilating plant parts had the highest proportion of nitrogen. Sterile plants of G. lutea allocated an average of 5.9% of their nitrogen resources to bulbils, those of G. spathacea 10.9%. On an average, flowering plants of G. lutea diverted 14.8% of their nitrogen resources to flowers, whereas flowering plants of G. spathacea had only 6.1% N allocated into flowers, but 18.4% into daughter bulbils, i.e. all together 24.5% to reproductive structures. However, these proportions seem to change with increasing total dry mass of a plant. Figure 4 shows the relations for dry mass and nitrogen content in logit-log plots. For both species, the investment in assimilating plant parts remains constant or decreases slightly with increasing size (G. lutea: dry mass, slope of the regression line 0.98, R 2 = 0.97; nitrogen content 0.97, R 2 = 0.99; G. spathacea: dry mass 0.99, R 2 = 0.94; nitrogen content 0.96, R 2 = 0.97). Investment into flowers increases in both species if plants become stronger, but this tendency is pronounced more in G. spathacea (G. lutea: dry mass 1.15, R 2 = 0.76; nitrogen content 1.24, R 2 = 0.76; G. spathacea: dry mass 1.66, R 2 = 0.41; nitrogen content 1.71, R 2 = 0.60). The most striking differences are found for bulbs and bulbils: whereas investment into bulbs remains almost constant in G. lutea, it decreases for G. spathacea (G. lutea: dry mass 0.99, R 2 = 0.98; nitrogen content 0.98, R 2 = 0.96; G. spathacea: dry mass 0.86, R 2 = 0.98; nitrogen content 0.83, R 2 = 0.96). In contrast, investment in bulbils increases only slightly for G. lutea, but strongly for G. spathacea (G. lutea: dry mass 1.15, R 2 = 0.76; nitrogen content 1.07, R 2 = 0.74; G. spathacea: dry mass 1.60, R 2 = 0.80; nitrogen content 1.47, R 2 = 0.76).

Distribution of nitrogen (per cent of total content) in sterile (n = 68) and flowering (n = 29) plants of Gagea lutea and sterile (n = 46) and flowering (n = 18) G. spathacea. Investigated were parent bulbs (open bars), bulbils (light grey), leaves (hatched), assimilating parts of inflorescences (reversely hatched), and flowers (dark grey)

Logarithmic plots showing the allocation of dry mass (a, b) and nitrogen (c, d) as a function of total dry mass and nitrogen content for Gagea lutea (a, c) and G. spathacea (b, d). Shown are values for assimilating plant parts (leaves and all inflorescence parts except for flowers, filled triangles), bulbs (filled circles), bulbils (open circles) and flowers (stars). Lines indicate the results of a regression analysis (see text)
Nitrogen content in seeds and bulbils differs strongly for G. lutea: seeds have a relatively constant dry mass (range 1.01–2.55 mg, mean 1.65 ± 0.34 mg, n = 5,392 from 93 plants) and contain 25.7 μg N per mg dry weight. The dry mass of bulbils fluctuates much more (range 0.22–3.22 mg, mean 1.30 ± 0.63 mg, n = 385), since their diameter reaches from 0.30 to 2.90 mm (mean 1.49 ± 0.42 mm, n = 1,955). With 13.8 μg N per mg dry mass, bulbils contain about half the amount of nitrogen, as compared to seeds. For G. spathacea we could not find seeds, but bulbils are comparable in size (diameter 0.11–3.61 mm, mean 1.27 ± 0.36, n = 3,572) and dry mass (range 0.09–2.62 mg, mean 1.46 ± 0.52 mg, n = 596); they contain 11.1 μg N per mg dry mass.
Using the values for nitrogen content of seeds and bulbils and the average number bulbils and/or seeds developed in plants of size classes of 0.5 mm parent bulb diameter, we estimated the total resources allocated to reproduction in both species (Fig. 5, potential seed for G. lutea set was derived from numbers of flowers, see below). In small stages, G. spathacea seems to allocate more resources to reproduction than G. lutea, for larger plants the ratio reverses strongly since large plants of G. lutea produce a high number of seeds.

Estimates for the amount of resources used for reproduction in cohorts of 0.5 mm parent bulb diameter for Gagea lutea (black and white bars) and G. spathacea (grey bars). Bars for G. lutea show the amount of resources (expressed as mg nitrogen) for bulbils (black parts) and seeds (white parts), whereas G. spathacea reproduces only by bulbils (grey bars). Total nitrogen content was calculated from the mean number of propagules developed by plants of the respective size class, with the potential seed set for G. lutea derived from the average number of flowers (compare Fig. 6). Average nitrogen contents found for seeds and bulbils of G. lutea (42.4 and 17.9 μg, respectively) and bulbils of G. spathacea (16.2 μg) were used
Pollen viability and seed set
For plants of both species, pollen grains are usually broadly obovate in shape. For G. lutea, viable grains measure 80.3 ± 8.1 μm × 57.5 ± 3.3 μm, corresponding to a volume of 0.141 ± 0.025 nl (120 grains measured). Non-viable grains are smaller (65.9 ± 8.4 μm × 49.9 ± 7.1 μm, 0.089 ± 0.030 nl, n = 123). The respective values for G. spathacea are 76.0 ± 11.3 μm × 60.2 ± 8.4 μm (0.151 ± 0.065 nl, alive, n = 68) versus 69.4 ± 10.4 μm × 53.0 ± 12.0 μm (0.112 ± 0.067 nl, non-viable, n = 155). For G. lutea, 60.7 ± 28.5% (SD, variation between individuals) of the pollen grains were found to be viable (1,829 grains of 9 individuals counted). In G. spathacea, this proportion was lower; but fluctuations between individuals were high (16.3 ± 22.8%, 1,230 grains of 16 individuals counted).
A total of 217 plants from five populations were checked for seed set in G. lutea (695 capsules with 8,926 seeds). Of these, only three plants were sterile; the mean seed set per plant was 40.8 ± 29.9 seeds. Each plant develop an average of 3.2 capsules with 12.7 ± 5.9 (maximum 33) seeds per capsule. For a cohort of 120 plants with 281 capsules, seeds were counted separately for each capsule, and only 26 capsules (9.3%) failed to develop seeds. As shown in Fig. 6, seed set increases steadily with capsule number. For G. spathacea, 177 plants from four populations, including 268 flowers, were surveyed repeatedly in the years 2006–2008. Without any exception, capsules did not grow further and failed to develop seeds.

Seed set per plant in dependence on capsule number for 217 plants of Gagea lutea, including standard error of means. Figures near the points indicate the number of plants with the respective number of capsules
Discussion
Including numerous species with slightly different growth patterns that can reproduce both vegetatively and sexually, the genus Gagea is especially interesting to compare resource allocation for vegetative and generative reproduction. As typical for monocotyledonous plants, all species follow a rather strict growth pattern. Already Irmisch (1850) analyzed these patterns and recognized the sympodial growth and the total replacement of all plant parts except for the parent bulb in a given year. However, as shown by Levichev (1999b), slight modifications in number and location of bulbs and bulbils can change tremendously the mode of vegetative reproduction and thus influence the spatial distribution of a population. For instance, replacement bulbs can be formed inside the parent bulb (like for the two species investigated herein), or outside [like in G. pratensis (Pers.) Dumort.]; the number of bulbils reaches from one to three per year (typical for G. pratensis), several (G. lutea) to many (G. spathacea); bulbils can be formed hypogeous (most often) or epigeous (some Central Asian species, like G. bergii Litv.); they can develop in the axil of their carrying leaf or on stolons of up to 10 cm length (Central Asian G. calyptriifolia Levichev); and finally, bulbils can germinate immediately or become dormant for one to several years. Such species-specific growth patterns seem to be typical for many Liliaceae of the tribe Tulipeae, as shown by Jäger (1973) for Tulipa sylvestris L.
Since all parts of a Gagea plant are renewed each year, an exact age determination of plants in nature is impossible, although age and size of the bulb are certainly correlated. Already a very rough estimation of the area of the cauline leaves is sufficient to produce a relatively good correlation with parent bulb diameter (Fig. 1). It can, therefore, be assumed that this leaf (G. lutea) or these two leaves (older plants of G. spathacea) are mainly responsible for accumulation of resources for the replacement bulb. If a surplus of resources is available, plants of both species start to develop bulbils. The thresholds in terms of diameter of the parent bulb are similar (2.63 mm in G. lutea vs. 2.43 mm in G. spathacea). When it comes to flowering, the threshold for G. lutea is considerably higher (6.94 vs. 5.34 mm). The most striking difference, however, is the existence of a “switch” in G. lutea: plants grow either bulbils or flowers in a given year, but never both structures together. Among 226 analyzed plants with bulbils and 238 with flowers, we did not find a single exception from this rule. Although not making an explicit statement, Peruzzi and Tison (2004) confirmed this by drawing a flowering plant without and a non-flowering plant with bulbils in their biosystematic study on the Italian species of Gagea. In contrast, G. spathacea continues to develop bulbils even if the critical bulb size to produce flowers is reached. This difference explains readily (1) the paucity of flowering plants in natural populations of the latter species and (2) the lower number of flowers per plant (1.51 in G. spathacea, 3.96 in G. lutea). In G. spathacea, the number of bulbils developed increases steadily with the diameter of the parent bulb (Fig. 2b). As a consequence, older plants of G. spathacea develop significantly more bulbils than in G. lutea; the maximum recorded number was 54 bulbils for a plant with two flowers. It seems to be the breakdown of the switch from vegetative to generative reproduction that allows G. spathacea to survive without producing seeds. The statement of Peterson et al. (2008b, p. 449) “in advanced age, the vegetative reproduction is absent” made for the sect. Spathaceae (monotypic, with G. spathacea as the single species) can thus not be maintained in the light of this study. In general, reproductive behavior seems to be under high selective pressure, since the species-rich section Gagea recognized by Peterson et al. (2008b) seems to include both strategies: species that continue vegetative reproduction parallel to flowering (like the mostly sterile G. pratensis, unpublished observations) and species where the two reproductive modes exclude each other mutually (as shown herein for the fertile G. lutea).
The mutual exclusion of seeds and bulbils in G. lutea seems to be in contradiction to the reproductive system in the genus Allium, where many species produce bulbils and seeds simultaneously. Ceplitis (2001) found evidence that annual fluctuations in fecundity of seed or bulbil production cause a stable balance between the two kinds of diaspores produced in the inflorescence of Allium vineale L., although the ratio seed to bulbils may fluctuate considerably between years. However, plants of G. lutea may also be able to switch between both modes during their individual lives. For instance, a bad year with only a weak accumulation of resources may cause a formerly flowering plant to fall below the respective threshold for flowering and hence to relapse to vegetative reproduction in the next year(s). In G. lutea, but probably in A. vineale as well, a plant cannot adapt to a given year, since the fate of a bud (forming flowers or bulbils) should be decided in the previous year, when the bud is initiated. This pattern was confirmed for Mayapple (Podophyllum peltatum L.) by Geber et al. (1997), where the rhizome forms a single shoot each year, with or without a flower. Thus, for Gagea as well as for Allium the average resource status of all plants achieved in the previous year should determine the ratio bulbils to seeds in a population for a given year. However, in G. lutea an individual plant reacts according to an either seeds or bulbils model, whereas an individual plant of A. vineale can grow a mixture of both.
The analysis of dry mass and nitrogen content, both measures of resources allocated to the respective structures, can help to elucidate the consequences of the superficially similar, but biologically quite different growth patterns in the two studied species of Gagea. When comparing cohorts of non-flowering and flowering plants in both species, already non-flowering plants of G. spathacea invest proportionally 1.9 times more nitrogen into vegetative reproduction (Fig. 3), and that does hardly change for flowering plants which invest 1.8 times more than individuals of G. lutea. Since the ratios given in Fig. 3 were recorded at flowering time, they may decrease to a certain extent by the time the plants of G. lutea develop seeds. This needs a considerable amount of resources for seeds, but inflorescence structures like bracts and the enduring tepals turning green after pollination contribute to provide resources. Looking at the absolute amount of resources (nitrogen), G. spathacea invests more into reproductive structures in earlier stages than plants of G. lutea of a similar size. Probably because of this reason G. spathacea falls later back behind G. lutea, since this species finally grows much larger bulbs (Fig. 5). This is reflected by the differences between the two species in resource allocation with increasing plant size (Fig. 4), whereas in G. lutea the proportion of resources allocated to bulbils remains roughly constant, it increases in G. spathacea with increasing plant size. This means that an individual invests proportionally more resources into vegetative reproduction the older it gets—which ultimately depletes its own resources. This explains the smaller bulb size of G. spathacea in comparison to G. lutea and may be a hint that individuals of G. lutea can reach a higher individual age.
Although not yet completely confirmed by quantitative observations, a number of species with odd chromosome sets in the genus Gagea seem to have lost the ability for generative reproduction. Gargano et al. (2007) found a greatly reduced seed set for the triploid Italian species G. chrysantha Schult. and Schult. f. s.l. and G. granatellii (Parl.) Parl., but also for G. fragifera (Vill.) Ehr. Bayer and G. López (reported as heptaploid) if compared to G. lutea with an even (hexaploid) chromosome set. For G. pratensis, most investigated populations of northern Germany had plants with 60 chromosomes (Henker 2005) and are mentioned to be sterile by this author. With x = 12 as the basic number of chromosomes for the genus, this would translate to a pentaploid level. In five populations of G. pratensis from Western Pomerania (182 plants with a total of 356 flowers) only three plants were found to develop a few seeds (M. Schnittler, personal observation). Peterson et al. (2008b) also found G. pratensis to be completely sterile (three investigated populations around Halle, Germany).
For G. spathacea, most authors cite sporophytic chromosome counts between 102 and 108 (Westergård 1936; Měsíček and Hrouda 1977; Peruzzi 2003; Henker 2005), fitting best the nonaploid level (9x = 108). In this study, all investigated populations of G. spathacea were found to be completely sterile; and Gustafsson (1946) observed a high degree of seed sterility for southern Sweden as well. Only Tomović and Niketić (2005) mention a “capsule with many sub-globose seeds” in their general description of the species, but do not give detailed observations for the two populations reported from Serbia. We assume that in (almost) all seed-sterile species the odd chromosome sets cause irregular meiosis and do not or rarely allow the formation of viable seeds (compare Westergård 1936). In contrast, seed set in the hexaploid G. lutea is normal and seems to increase linear with the number of capsules (Fig. 6). Although we did not observe the flowering sequence, our observations do not confirm those of Nishikawa (1998) who found in this species a lower seed output for later flowers than for early ones.
Similar to our results for G. spathacea, Caparelli et al. (2006) found for the triploid G. granatellii that lack of seed production correlates with a high percentage of malformed pollen. Pollen fertility is generally reduced in species of Gagea with higher ploidy levels (Zarrei and Zarre 2005), but odd ploidy levels seem to be especially detrimental to pollen fertility (Gargano et al. 2007). However, our results for G. spathacea suggest that even for species that fail to produce seeds, pollen still contains a small proportion of viable grains. In G. spathacea, the number of pollen grains per anther seems to be smaller, grains are often malformed and stick together, whereas pollen grains of G. lutea are more numerous, better separated from each other and appear to be more regular in size. In both species, living (fluorescing) grains tend to be larger than dead grains.
All these observations on seed and pollen fertility of Gagea species may be explained best by the following assumptions: (1) seed set, to a lesser extent also pollen fertility, is generally negatively affected by increasing ploidy levels. (2) Odd levels (tri-, penta-, hepta- and nonaploid states) reduce pollen viability and cause at least a partial seed sterility. However, even highly polyploid species with odd chromosome sets seem to be able to produce at least some viable pollen, as indicated by our results for G. spathacea. Pollen may cross-pollinate related species to form hybridogenous seeds. We thus agree with Peterson et al. (2004, 2008a) and Peruzzi (2008b) that hybridization is a major cause for speciation in the genus. Vegetative reproduction via bulbils allows primary hybrids with odd chromosome sets to survive and even multiply. This is likely the reason for the many different ploidy levels recorded in the genus Gagea (known are all levels from 2x to 9x in Europe, compare Peruzzi and Aquaro 2005) as well as in the genus Ornithogalum showing a similar life history (basic number 9, levels 2x – 10x, van Raamsdonk 1985; in addition dysploid forms exist).
From an ecological point of view, bulbils have different features in comparison to seeds. First, seeds are generative diaspores, which create a higher genetic diversity in offspring than the bulbils which are genetically identical with their mother plants. Second, seeds are formed epigeous and are more durable than bulbils. As an obvious short term-advantage, the elaiosome in many European species of sect. Gagea allows dispersal over some distance, which should only exceptionally happen for the subterranean bulbils, e.g., via digging animals like wild boar (Sus scrofa) observed in one population of G. spathacea in 2008. On the other hand, seeds confer a higher cost of reproduction. A seed of the investigated G. lutea carries more than twice the amount of resources of a bulbil, if seen in terms of nitrogen content: 42.4 μg for an average-sized seed but 17.9 μg for an average-sized bulbil. Developing flowers infers additional costs, but these may be (at least partly) counterbalanced by the additional assimilation of flowering structures during seed maturation. However, in a study in the genus Allium, capable as well to reproduce with both epigeous seeds and bulbils, Ceplitis and Bengtsson (2004) also identified advantages for bulbils. First, they seem to be more robust than seeds and have a higher germination rate. Second, they lead to whole cohorts of plants belonging to a single genet, which were observed to be less competitive to each other than populations of genetically diverse individuals. Finally, germination from bulbils produced fewer plants with developmental abnormities than generative seedlings. In contrast to the bulbils of Allium that are formed instead of flowers, the two species of Gagea studied by us develop subterranean bulbils. Regarding the germination rate, this could well be an advantage in comparison to seeds, but almost certainly it hampers dispersal compared with aboveground diaspores. Seeds of G. lutea have a deep simple epicotyl morphophysiological dormancy (Kondo et al. 2004), and seedlings appear at the same time as bulbils sprout (personal observation of the authors).
Not yet investigated are the genetic consequences of the two different reproductive strategies. At least for G. lutea, the regular generative reproduction via seeds should create a rather high level of genetic diversity. Seeds seem to be effective founders of new populations in the species. An indirect proof is the high level of genetic polymorphism (71.4%) found with a RAPD analysis between populations of G. lutea (Peterson and Peterson 1999). So far, no molecular analyses are available for G. spathacea. If our results are valid for the species as a whole, it should be a truly asexual species with little genetic divergence, relying entirely on vegetative reproduction via bulbils for survival. According to prevalent theory, obligate vegetative reproduction should be selected against due to the mutational load that may accumulate (see Silvertown 2008 for review). However, polyploidy may protect such plants from detrimental effects (Archetti 2004). If this holds true, G. spathacea is one of the rare examples of plants showing an odd set of chromosomes connected with a partial or complete loss of sexual reproduction. Most of these species reproduce asexually by agamospermy (e.g., many species of Taraxacum) and tend to be polyploid (Richards 2003). For G. spathacea, vegetative reproduction without seeds may be able to compensate the loss of sexual reproduction as well, but seems to have its prize in terms of individual fitness.
References
Archetti M (2004) Recombination and loss of complementation: a more than two-fold cost for parthenogenesis. J Evol Biol 17:1084–1097
Ashman TL (1994) Reproductive allocation in hermaphrodite and female plants of Sidalcea oregana ssp. spicata (Malvaceae) using four currencies. Am J Bot 81:433–438
Caparelli K, Peruzzi L, Cesca G (2006) A comparative analysis of embryo-sac development in three closely-related Gagea species (Liliaceae), with some considerations on their reproductive strategies. Plant Biosyst 140:115–122
Ceplitis A (2001) Genetic and environmental factors affecting reproductive variation in Allium vineale. J Evol Biol 14:721–730
Ceplitis A, Bengtsson BO (2004) Genetic variation, disequilibrium and natural selection on reproductive traits in Allium vineale. J Evol Biol 17:302–311
Gargano D, Peruzzi L, Caparelli K, Cesca G (2007) Preliminary observations on the reproductive strategies in five early-flowering species of Gagea Salisb. (Liliaceae). Bocconea 21:349–358
Geber MA, de Kroon H, Watson MA (1997) Organ preformation in mayapple as a mechanism for historical effects on demography. J Ecol 85:211–223
Gustafsson A (1946) Apomixis in higher plants. Part I. The mechanisms of apomixis. Lunds Univ Arsskr Adv 2(42):1–67
Haeupler H (1969) Bestimmungsschlüssel der Gagea-Arten im südlichen Niedersachsen im blütenlosen Zustand. Mitt. Florist-soziol Arbeitsgem 14:36–46
Henker H (2005) Goldsterne und Stinsenpflanzen in Mecklenburg-Vorpommern. Botanischer Rundbrief 39:14–37
Hultén E, Fries M (1986) Atlas of North European vascular plants: north of the Tropic of Cancer I–III. Koeltz Scientific Books, Königstein. http://linnaeus.nrm.se/flora. Accessed 20 Dec 2007
Irmisch Th (1850) Zur Morphologie der monokotylischen Knollen- und Zwiebelgewächse. Berlin
Jäger E (1973) Zur Verbreitung und Lebensgeschichte der Wildtulpe (Tulipa sylvestris L.) und Bemerkungen zur Chorologie der Gattung Tulipa L. Hercynia NF 10:429–448
Kearns CA, Inouye DW (1993) Techniques for pollination biologists. University Press of Colorado, Boulder
Kondo T, Miura T, Okubo N, Shimada M, Baskin C, Baskin J (2004) Ecophysiology of deep simple epicotyl morphophysiological dormancy in seeds of Gagea lutea (Liliaceae). Seed Sci Res 14:371–378
Korneck D, Schnittler M, Vollmer I (1996) Rote Listen und Florenlisten gefährdeter Pflanzen in Deutschland: Florenliste und Rote Liste der Farn- und Blütenpflanzen (Pteridophyta et Spermatophyta). Schriftenreihe Vegetationsk. 28:21–187
Levichev IG (1999a) Phytogeographical analysis of the genus Gagea Salisb. (Liliaceae). Komarovia 1:45–47
Levichev IG (1999b) The morphology of Gagea Saliasb. (Liliaceae) I. Subterranean organs. Flora 194:379–392
Levichev IG (2008) Historia Gagearum. http://www.binran.ru/infsys/gagea/index-eng.html. Accessed 10 Nov 2008)
Měsíček J, Hrouda L (1977) Chromosome numbers in Czechoslovak species of Gagea (Liliaceae). Folia Geobot Phytotax 9:359–368
Meusel HE, Jäger EJ, Weinert E (1965) Vergleichende Chorologie der zentraleuropäischen Flora, vol 1. Fischer, Jena
Nishikawa Y (1998) The function of multiple flowers of a spring ephemeral, Gagea lutea (Liliaceae), with reference to blooming order. Canad J Bot 76:1404–1411
Patterson TB, Givnish TJ (2002) Phylogeny, concerted convergence, and phylogenetic niche conservatism in the core Liliales: insights from rbcL and ndhF sequence data. Evolution 56:233–252
Peterson A, Peterson J (1999) Genotypic differentiation on an individual level in Gagea villosa (M. Bieb.) Duby, Gagea lutea (L.) Ker-Gawler and Gagea bohemica subsp. saxatilis (Koch) Pascher in Saxony-Anhalt using RAPD markers. J Appl Bot 73:228–233
Peterson A, Hohn H, Koch E, Peterson J (2004) A molecular phylogeny of the genus Gagea (Liliaceae) in Germany inferred from non-coding chloroplast and nuclear DNA sequences. Plant Syst Evol 245:145–162
Peterson A, Harpke D, Peruzzi L, Tison J-M, John H, Peterson J (2008a) Hybridization drives speciation in Gagea (Liliaceae). Plant Syst. Evol. doi: 10.1007/s00606-008-0102-3 (in press)
Peterson A, Levichev IG, Peterson J (2008b) Systematics of Gagea and Lloydia (Liliaceae) and infrageneric classification of Gagea based on molecular and morphological data. Mol Phylogen Evol 46:446–465
Peruzzi L (2003) Contribution to the cytotaxonomical knowledge of Gagea Salisb. (Liliaceae) sect. Foliatae A. Terracc. and synthesis of karyological data. Caryologia 56:115–128
Peruzzi L (2008a) Contribution to the cytotaxonomical knowledge of the genus Gagea Salisb. (Liliaceae). III. New karyological data from the central Mediterranean area. Caryologia 61:92–106
Peruzzi L (2008b) Hybridity as a main evolutionary force in the genus Gagea Salisb. (Liliaceae). Plant Biosyst 142:179–184
Peruzzi L, Aquaro G (2005) Contribution to the cytotaxonomical knowledge of Gagea Salisb. (Liliaceae). II. Further karyological studies on Italian populations. Candollea 60:237–253
Peruzzi L, Tison JM (2004) Verso una revisione biosistematica del genere Gagea Salisb. (Liliaceae) in Italia. Un nuovo tipo di approccio. Inform Bot Ital 36:470–475
Peruzzi L, Peterson A, Tison J-M, Peterson J (2008) Phylogenetic relationships of Gagea Salisb. (Liliaceae) in Italy, inferred from molecular and morphological data matrixes. Plant Syst Evol. doi: 10.1007/s00606-008-0081-4 (in press)
Richards AJ (2003) Apomixis in flowering plants: an overview. Philos Trans Ser B 358:1085–1093
van Raamsdonk LWD (1985) Pollen fertility and seed formation in the Ornithogalum umbellatum/angustifolium complex (Liliaceae/Scilloideae). Plant Syst Evol 149:287–297
Rothmaler W (founder), Jäger E, Werner K (eds) (2005). Exkursionsflora von Deutschland, vol 4. Gefäßpflanzen: Kritischer Band, 10th edn. Spektrum Akademischer Verlag, Munich
Schnittler M, Günther K-F (1999) Central European vascular plants requiring priority conservation measures—an analysis from national Red Lists and distribution maps. Biodivers and Conserv 8:891–925
Silvertown J (2008) The evolutionary maintenance of sexual reproduction: evidence from the ecological distribution of asexual reproduction in clonal plants. Int J Plant Sci 169:157–168
Tomović G, Niketić M (2005) Gagea spathacea (Hayne) Salisb. (Liliaceae)—a new species for the flora of Serbia. Ann Biol Sci Belgrad 57(4):291–294
Uphof JCT (1958–1960) A review of the genus Gagea Salisb. Plant Life 14: 124–132; 15: 151–161; 19: 163–176
Westergård M (1936) A cytological study of Gagea spathacea with a note on the chromosome number and embryo-sac formation in Gagea minima. CR Trav Carlsberg Lab Sér Physiol 21:437–451
Zarrei M, Zarre S (2005) Pollen morphology of the genus Gagea (Liliaceae) in Iran. Flora 200:96–108
Acknowledgments
Most of the measurements for morphological data were carried out within several student courses, and we wish to thank all participants, especially C. Brummond, J. Dobbert, N. Formella, H. Kasten, A. Krüger, M. Liermann, L. Lohmann, J. Mohr, L. Niehusen, A. Roschanski, S. Schmidt, and A.-K. Zießnitz. For permission and advice to use the N/C analyzer we are indebted to U. Möbius and S. Zerbe; for use of a fluorescence microscope to S. Stremlau and Ch. Stöhr. Search for localities, but also the organization of field work was supported by S. Starke and A. Klahr (all Institute of Botany and Landscape Ecology). For advices, especially concerning localities, we are grateful to H. Henker (Wismar). We wish to thank the “Staatliches Amt für Umwelt und Natur Ueckermünde” for permission to collect plants and two anonymous reviewers for useful comments and advices.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Schnittler, M., Pfeiffer, T., Harter, D. et al. Bulbils contra seeds: reproductive investment in two species of Gagea (Liliaceae). Plant Syst Evol 279, 29–40 (2009). https://doi.org/10.1007/s00606-008-0143-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00606-008-0143-7