
Abstract
Graphite-rich metamorphic limestones included within low-grade black schists of the lowest Nevado-Filábride tectonic unit in the Sierra de Baza (Bodurria Unit) provided the first conodonts found in this complex (Declinognathodus bernesgae, D. inaequalis. D. cf. praenoduliferus, and Idioprioniodus sp.). This demonstrates the early Bashkirian age of the sedimentary protoliths, and their deposition in open marine anoxic environments of a continental margin that opened towards the E and that was related to the Palaeotethys. This breakthrough offers strong support for the hypothesis that the lower Nevado-Filábride units show stratigraphic and palaeogeographic affinities with the external domains of the Iberian Massif (Palentian Domain of the Cantabrian Zone in particular). Consequently, this part of the Betic Internal Zones must be related to the basement of the South-Iberian Palaeomargin and excluded from the Alborán Domain tectonostratigraphic terrane.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
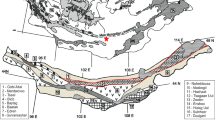
The Variscan–Alleghenian Orogen in Spain (Fig. 1) includes the Iberian Massif (Martínez Catalán 2011; Simancas et al. 2013), but also fragments that were detached from it during the Mesozoic breakup of Pangea and that were later involved in the Alpine Orogen (Fig. 2A). Several of these fragments make part, at present, of the three main complexes of the Internal Zones of the Betic Cordillera (Nevado-Filábride, Alpujárride and Maláguide) where pre-Mesozoic sedimentary successions and Palaeozoic magmatic rocks associated with them, frequently affected by intense metamorphism, are widely present (Fig. 2B). The location of one of these fragments, the Nevado-Filábride Complex (Gómez-Pugnaire et al. 2004, 2012), makes it a key terrane to interpret the geometry and evolution of the Variscan Orogen in SE Spain and the overall picture of the Pangea amalgamation and later breakup.

(modified from Martínez Catalán 2011)
Tectonic map of the Iberian Massif including the different palaeogeographic zones

Tectonic map of the Betic Cordillera. Black areas in A indicate the main outcrops of Palaeozoic rocks in the Internal Domains of the Western Mediterranean Alpine Belts
The stratigraphy of the Nevado-Filábride Palaeozoic is poorly known due to metamorphic overprint and deformation during Variscan and Alpine orogenies. Consequently, identifiable fossils are rare and provide imprecise biostratigraphic information. The present study reports the first conodonts, Bashkirian in age, found in the metamorphic succession of the lowest tectonic unit of the Nevado-Filábride Complex. This allows us: (1) to determine the depositional environment of the protoliths; (2) to correlate the stratigraphic succession of the studied sediments to the outermost sectors of the Variscan Orogen in Northern Spain; and (3) to discuss their significance for the geometry and zoning of the Variscan belt SE-wards of the Iberian Massif in the late Carboniferous palaeogeographic framework.
Geological setting
Iberian Massif
The Iberian Massif features an S-shaped double-vergent orocline (Fig. 1) formed when several oceanic domains (like the Rheic and Rhenohercynian oceans) closed and Pangea was assembled by collision of Gondwana, Laurussia, and other intervening terranes (Weil et al. 2000; Matte 2001; Murphy et al. 2006, 2009; Martínez Catalán et al. 2007; Nance et al. 2010; Pereira et al. 2010, 2012; Martínez Catalán 2011, 2012; Shaw et al. 2012; Kroner and Romer 2013; Franke et al. 2017). Stratigraphic, tectonometamorphic, and magmatic criteria allow a division of the Iberian Massif into the Cantabrian, West Asturian-Leonese, Central Iberian, Galicia Tras-os-Montes, Ossa-Morena, and South-Portuguese Zones (Lotze 1945; Julivert et al. 1972). The latter zone is the most external domain in the Southern Iberian Massif and was affected by SW-directed thin-skinned tectonics involving uppermost Devonian-Lower Carboniferous successions (Simancas et al. 2003). The boundaries between the South-Portuguese and Ossa-Morena Zones and between the Galicia Tras-Os-Montes and Central Iberian Zones are generally interpreted as the suture of the Rheic Ocean, but with opposite subduction polarity (Martínez Catalán et al. 1996, 1997). The Galicia Tras-Os-Montes Zone is made of oceanic units derived from the Rheic Ocean and of continental units with an “out-of-Gondwana” provenance. Both were thrusted onto the Central Iberian Zone that, together with the West Asturian-Leonese and Cantabrian zones, made part of N-Gondwana. The Ossa-Morena/Central Iberian boundary is also interpreted as a suture, but related to the closure of a narrow (back-arc?) ocean that (like the Rhenohercynian Ocean of Central Europe, cf. Franke et al. 2017) was younger than the Rheic Ocean, independent of it (Gómez-Pugnaire et al. 2003; Simancas et al. 2013) and opened during incomplete breakup of the Armorican ribbon continent from N-Gondwana (Robardet 2003). Eastwards of the Central Iberian Zone, the West Asturian Leonese Zone represents a domain affected by E-vergent ductile deformation and low-to-medium-grade metamorphism. Finally, the Cantabrian Zone is the most external zone of the orogen that is preserved within the Iberian Massif, and is characterized by thin-skinned tectonics and oroclinal deformation involving Palaeozoic sedimentary rocks (Pérez-Estaún et al. 1988).
The orogenic evolution of the outer zones of the Iberian Massif started with the latest Devonian-earliest Carboniferous closure of the Rhenohercynian Ocean, but its final (postcollisional) evolution was Late Carboniferous–Early Permian, when deformation ceased after formation of the Cantabrian orocline (Pastor-Galán et al. 2014). A persisting active subduction yet remained, however, eastwards of the Iberian Massif in the northern margin of the Palaeotethys Ocean. Actually, the onset of Palaeotethyan subduction below Iberia in latest Carboniferous–earliest Permian time has been recently invoked as the best favourable geodynamic setting to explain the features and age of the omnipresent calc-alkaline late-Variscan postcollisional magmatism in Iberia, which is several millions of years younger than the complete closure of the Rhenohercynian and other oceans (Pereira et al. 2015).
The Palaeotethys Ocean closed during the Permian–Triassic, while coeval rifting and seafloor spreading in northern Gondwana gradually opened the Neotethys (Hsü and Bernoulli 1978; Şengör 1990; Şengör et al. 1988; Stampfli 2000). Some authors interpret that, from Silurian to Carboniferous time, the Palaeotethys Ocean was continuous from the Alpine–Himalayan region towards the SW up to Central America, through the area between southern Iberia and N Africa (Stampfli and Borel 2002, 2004; Cocks and Torsvik 2006; Stampfli and Kozur 2006; Stampfli et al. 2003, 2013; Von Raumer et al. 2002, 2003). However, most regional studies on Iberian and N African Palaeozoic successions indicate that the Palaeotethys did not continue towards the W (Michard et al. 2008; Simancas et al. 2009), and that it ended somewhere within the Palaeozoic terranes now involved in the Internal Zones of the Western Mediterranean Alpine Orogen and/or in the Moroccan Mesetas, where there is no evidence for oceanic crust or related volcanism. This is further supported by stratigraphic studies on Palaeozoic Alpine terranes in Italy and Spain (Navas-Parejo et al. 2009a , b, 2015; Navas-Parejo 2012; Rodríguez-Cañero and Martín-Algarra 2014).
Nevado-Filábride Complex of the Betic Cordillera
The Betic Cordillera is part of Western Mediterranean Alpine Orogen. The Nevado-Filábride, Alpujárride, and Maláguide tectonometamorphic complexes (Fig. 2B), together with detached continental margin Meso-Cenozoic covers (Frontal Units), constitute its Internal Zones (Vera 2004) or Alborán Domain (Balanyá and García-Dueñas 1987). This allochthonous terrane has been recently redefined by excluding from it the Nevado-Filábride Complex (Gómez-Pugnaire et al. 2004, 2012).
The Betic Orogen was formed by the early Miocene subduction of the oceanic floor that constituted the basement of the Betic–Rifian Flysch complexes (Fig. 2B) below the Alborán Domain, which collided against the S Iberia and N Africa palaeomargins later in the Miocene. At that time, the Nevado-Filábride Complex (NFC) was affected by HP metamorphism (López Sánchez-Vizcaíno et al. 2001), because it subducted and finally accreted below the Alborán Domain as it would probably make up part of the Southern Iberian continental crust and margin (Gómez-Pugnaire et al. 2012).
The NFC outcrops exclusively in the central–eastern Betic Cordillera and is located in three mayor E–W trending antiforms: the Sierras Alhamilla, Nevada, and Filabres (Fig. 3). Eastwards, left-lateral strike-slip faults rotate and translate them towards the N to form the Águilas arc (Fig. 3). Two major tectonic ensembles called Veleta (bottom) and Mulhacén (top) are commonly distinguished within the NFC in Sierra Nevada (Fig. 3). The Veleta-type successions are several km thick and composed of pre-Mesozoic black schists (locally including thin marble layers) and quartzites (Aulago Formation: Martínez-Martínez 1986) that underwent metamorphism in greenschist facies (Gómez-Pugnaire and Franz 1988).

Nevado-Filábride Complex, and its division in Veleta and Mulhacén successions
The Mulhacén successions constitute a tectonic amalgamation of several thin rock units (see Jabaloy et al. 2015), including continental-crust successions of dark (graphite-rich) pre-Mesozoic metapelites (Montenegro Schists: Martínez-Martínez 1986), younger light-coloured metasediments (siliciclastic and carbonatic), and other units with ultramafic rocks. Both the continental and ultramafic units contain metabasite bodies. A succession of controversial age, formed by metaevaporites, calcitic and dolomitic marbles, quartzites, and more or less calcareous chloritic–amphibolic schists, comprises the uppermost part of the Mulhacén successions (Puga et al. 2011; Gómez-Pugnaire et al. 2012). All the successions underwent intense, subduction-related HP Alpine metamorphism (Puga et al. 2002, 2017; Jabaloy et al. 2015).
The Veleta (Aulago) and Mulhacén (Montenegro) schists are quite similar, but the Montenegro schists were much more intensely affected by metamorphism, both Alpine and pre-Alpine. The pre-Alpine metamorphism in the Montenegro schists caused static blastesis of large porphyroblasts (garnet, chloritoid, staurolite, and chiastolitic andalusite) that are texturally very well preserved locally, although they are systematically overprinted by the HP Alpine metamorphism (Puga et al. 1975). A cartographic unconformity bounds the Montenegro schists and quartzites from the overlying Tahal Formation, probably of Permian age and made up of metapsammites with metaconglomerate lenses and of light-coloured chloritoid-bearing albitic schists (Gómez-Pugnaire 1981; Jabaloy 1993). Acid to intermediate Late-Variscan granitoids locally intrude on the continental sequences of the Mulhacén successions, with zircons yielding latest Carboniferous—early Permian U–Pb SHRIMP ages (Gómez-Pugnaire et al. 2004, 2012).
The first NFC fossils come from Veleta-type rocks of the Águilas Arc (Fig. 3): the Lomo de Bas succession (Álvarez-Lobato and Aldaya 1985; Álvarez-Lobato 1987). These are Eifelian chaetetids (Chaetetes cf. salairicus Dubatolov: Lafuste and Pavillon 1976). Recently, Laborda-Lopez et al. (2013, 2015a, b) proposed an Emsian age for crinoids (Bystrowicrinus sp. and Pentagonopentagonalis sp.) and unclassifiable macrofaunal remnants (phillipsastreid rugose corals, cephalopods, gastropods, brachiopods, trilobites, benthic foraminifera, and possible algal thali) that they found in the same area. The new litho- and biostratigraphic data provided here come from one Veleta-type unit in Sierra de Baza, the westernmost part of the Filabres antiform (Fig. 4A). In this area, Gómez-Pugnaire et al. (1982) previously found Neoproterozoic acritarchs (Gloecapsomorpha sp. and Trematosphaeridium sp.).

A Geological map of the Nevado-Filábride Complex in northern Sierra de Baza. B Maps and cross sections of the four tectonic windows of the Bodurria Unit. Red star: Collado de Floranes section
Bodurria unit
Most NFC rocks in Sierra de Baza correspond to a Mulhacén-type succession (Filabres Unit) that overthrusts a lower set of Veleta-type tectonic successions by means of a brittle–ductile shear zone with a sense of transport towards the W (Jabaloy 1993). The Bodurria Unit, of Veleta-type, outcrops below the Filabres Unit in four small tectonic windows called, from E to W: Petronila, Bodurria, Zar, and Rambla del Agua (Fig. 4B). It is made of low-to-very low-grade metamorphic rocks with very weak strain, quite similar to those of the lower part of the Filabres Unit and affected by S-verging folds.
All tectonic windows of the Bodurria Unit show equivalent stratigraphic successions, with two main parts (Fig. 5). The lower part is formed by a few dozen metres of low-grade graphite-rich black schists and slates that alternate with thinly-to-medium layered (occasionally thickly layered) quartzitic horizons that become progressively more abundant upwards. The dark schists include a decametre-thick laterally discontinuous intercalation of finely detrital and well-laminated, black-to-brownish thin carbonate layers alternating with black slates (Figs. 5 and 6A–E).

Left Synthetic lithostratigraphy of the Bodurria Unit. Righ t Section measured and sampled for conodonts in the Collado Floranes area, with location of the productive samples (stars)
The upper part of the succession is lighter coloured and several hundred of metres thick. It is dominated by thickly-to-very thickly layered quartz-rich psammites with subordinate black-to-dark grey schists similar to those of the underlying succession. This stratigraphic upper part of the Bodurria Unit is made of ca. 250 m of alternating 1–10-m-thick packets of medium-to-very thickly layered and medium-sand-to-coarse-sand-grained whitish-to-greyish quartz-rich psammites (quartzwackes) separated by thin layers of black slates (Fig. 6F), and packets made of finer-grained psammites alternating with some black-to-grey slates. Upwards, quartzitic packets become thicker. Intense folding makes it generally difficult to recognize upward trends in the evolution of bed thickness, but thinning-upwards sequences are locally visible within a general thickening-upwards trend (Bodurria and Petronila windows). The psammites locally preserve sedimentary structures, including: scour surfaces, graded bedding, parallel and cross lamination, undulatory to linguoid ripples, flaser structures, and burrows (Fig. 6G, H).

Field views of the Bodurria Unit rocks. A, B Lower and middle parts of the section of alternating black-brownish metalimestones and schists (slates) measured in the Petronila window (Floranes outcrop: 37°18′49″N, 2°46′50″W); numbers indicate the position of the samples (mostly barren of conodonts) with arrows pointing to the location of two conodont-bearing samples (15A-34 and 15A-37). C Black metalimestones of the core of the Bodurria window antiform, with numbers indicating the location of samples for conodonts (barren at that site: 31°17′46″N, 2°49′31″W); note the fold vergence towards the S, and the presence of two blind fold-thrusts in its very core (arrows). D Close up of one parallel-laminated metalimestone bed between black slates (Floranes section). E Laminated and slightly bioturbated black metalimestone bed (Bodurria window). F Well-bedded black slates stratigraphically overlying the metalimestones shown in picture (c). G Rippled metapsammites of the stratigraphic upper part of the Bodurria Unit in the northern limb of the Bodurria window antiform (37°17′57″N, 2°49′34″W): note linguoid to undulating asymmetric ripples with sinuous crests that, after restoring bedding to horizontal, indicate palaeocurrents directed towards the SE (arrow). H Cross lamination of one metapsammite bed in the same outcrops as G; note migration of ripples towards the SE (right)
First conodonts of the NFC
Materials and methods
Three carbonate-bearing sections were sampled for conodont studies (Figs. 4B, 5): (1) the Collado de Floranes area near to the eastern end of the Petronila window; (2) the core of an antiform in the central part of the Bodurria window; and (3) the central part of the Zar window. In these outcrops, fine-grained black slates alternate with fine sandy-to-silty, parallel or locally cross-laminated and occasionally burrowed, very thinly-to-thinly bedded (5–20 cm) black metalimestones very rich in graphite, with brownish weathering colours (Fig. 6A–E). Under the microscope, the organic matter concentrates in well-defined parallel laminae. Lighter-coloured lithotypes poorer in organic matter exhibit brownish–yellowish hues due to the presence of Fe oxyhydroxides and/or phosphate. The metalimestones contain small amounts of silt-sized-to-very fine-sand-sized detrital quartz grains and mica-chlorite flakes.
Finding this conodont fauna required patient and thorough work, because conodont elements from these levels are very scarce as only 3 of 56 samples processed (weighting > 100 kg in total) yielded a few conodont elements reliable for taxonomic identification. Samples were processed using standard dissolution techniques with both acetic (Jeppsson et al. 1999) and formic (Ellison and Graves 1941) acids and heavy liquid separation using sodium metatungstate (Anderson et al. 1995).
The conodont elements collected are not very well preserved, being texturally altered and partly broken. They have a uniform CAI value of 5 indicating temperatures of 300–480 °C consistent with chlorite grade regional metamorphism (Epstein et al. 1977; Rejebian et al. 1987), which agrees with the metamorphic conditions deduced from the schists associated with the metalimestones (Jabaloy 1993). The conodont specimens are stored in the Dpto. de Estratigrafía y Paleontología of Granada University.
The conodont association found has a very low diversity, with only two genera and three species. Table 1 summarizes the number of conodont elements for each taxon recognized: Declinognathodus bernesgae Sanz-López, Blanco-Ferrera, García-López, and Sánchez de Posada 2006, D. inaequalis (Higgins 1975), D. praenoduliferous Nigmadganov and Nemirovskaya 1992, plus Idioprioniodus sp. and ramiform elements that correspond to P2 elements of Declinognathodus sp. and to S elements of Gnatodonthid species (see also Fig. 7).

Bashkirian conodonts of the Bodurria Unit. (1) Declinognathodus bernesgae Sanz-López, Blanco-Ferrera, García-López, and Sánchez de Posada 2006. P1 element; SF1-10. (2) Declinognathodus bernesgae Sanz-López, Blanco-Ferrera, García-López, and Sánchez de Posada 2006. P1 element; SF1-7. (3) Declinognathodus cf. D. praenoduliferous Nigmadganov and Nemirovskaya 1992. P1 element; 15A37-1. (4) Declinognathodus inaequalis (Higgins 1975). P1 element; SF1-6. (5) Declinognathodus bernesgae Sanz-López, Blanco-Ferrera, García-López, and Sánchez de Posada 2006. P1 element; SF1-4. (6) Declinognathodus cf. D. bernesgae Sanz-López, Blanco-Ferrera, García-López, and Sánchez de Posada 2006. P1 element; 15A37-4. (7) Declinognathodus cf. D. praenoduliferous Nigmadganov and Nemirovskaya 1992. P1 element; 15A37-6. (8) Idioprioniodus sp. element; 15A37-2. (9) Idioprioniodus sp. element; 15A37-3. (10) Declinognathodus sp. P2 element; SF1-13. (11) Declinognathodus sp. P2 element; SF1- 12. (12) Gnathodontid S element; 15A37-7. (13) Gnathodontid S element; 15A34-1
Systematic palaeontology
Class Conodonti Branson, 1938
Order Ozarkodinida Dzik, 1976
Family Idiognathodontidae Harris and Hollingsworth, 1933
Genus Declinognathodus Dunn, 1966
Type species: Cavusgnathus nodulifera Ellison and Graves, 1941
Diagnosis P1 elements mostly show Class II symmetry (Lane 1968). P1 element of Declinognathodus is carminiscaphate with an elongate, narrow platform divided into two unequal parapets separated by a median, longitudinal, more or less developed trough. The ornamentation of the parapets is made of nodules or ridges. The free blade joins platform in a medial position; its dorsal continuation, the medial carina, is declined to one side, merges and continues dorsally as the rostral (external) platform parapet. One or several nodules appear along the ventral–rostral (anterior–external) margin of the platform. Conodont apparatus is unknown.
Remarks Some authors considered Declinognathodus and Idiognathoides as synonyms (Lane 1967; Lane and Straka 1974; Grayson et al. 1990). The original definition of Declinognathodus by Dunn (1966) specified the differences to Cavusgnathus and Streptognathodus, from which it clearly differs, but not from Idiognathoides. As Grayson et al. (1990) stated, although the apparatus of Declinognathodus has not been established yet, some elements of Idiognathoides, Declinognathodus, and Gnathodus, in particular P2 and S elements, are very similar.
Declinognathodus bernesgae Sanz-López, Blanco-Ferrera, García-López, and Sánchez de Posada, 2006
Figs. 7.1, 7.2, 7.5, 7.6
*2006 Declinognathodus noduliferous bernesgae Sanz-López, Blanco-Ferrera, García-López, and Sánchez de Posada, pl. 1, figs. 8–18.
2011 Declinognathodus bernesgae. (Nemyrovska et al., pl. 3, figs. 11, 13, 14; pl. 4, figs. 4, 16, 17, 25, 26).
2013 Declinognathodus bernesgae. (Sanz-López et al., fig. 7G–Y).
Material Nine P1 elements.
Diagnosis P1 element has two parapets with transverse nodes on both sides of the platform separated by a shallow groove that disappears in the dorsal (posterior) part of the platform, where up to four transverse ridges occur. Carina is deflected to the rostral (external) side and there are one or two nodules on the ventral–rostral (anterior–external) margin of the platform. Nigmadganov and Nemyrovskaya (1992) and Nemyrovska et al. (2011) did not always agree with Sanz-López et al. (2006), and Sanz-López and Blanco-Ferrera (2013) concerning the accurate identification of this species.
Remarks The elements SF1-10 (Fig. 7.1) and SF1-7 (Fig. 7.2) are quite well preserved and they agree with the diagnosis of this species. The element SF1-4 (Fig. 7.5) does not have a free blade, but the platform shows well-diagnostic features. The element 15A-37-4 (Fig. 7.6) is assigned to this species without confidence, because the dorsal platform and the free blade are broken. However, the blade is in a middle position, the short carina is declined to the rostral side, a groove exists between both parapets, and also a little nodule on the ventral–rostral margin of the platform.
Range Lowermost Bashkirian (Nemyrovska et al. 2011); Upper Serpukhovian-Lower Bashkirian (Sanz-López and Blanco-Ferrera 2013).
Declinognathodus inaequalis (Higgins 1975)
Fig. 7.4
*1975 Idiognathoides noduliferous inaequalis Higgins, pl.12, figs. 1–7,12; pl.14, figs. 11–13; pl. 15, figs. 10,14
2013 Declinognathodus inaequalis. (Sanz-López and Blanco-Ferrera, Fig. 5G–O).
Material One P1 element.
Diagnosis P1 element has a prominent carina joined to the rostral (external) parapet in the middle of the platform and separated from the ventral (anterior) part of the parapet by a long, deep through. The rostral (external) parapet consists of a ridge-like, only present in the ventral part of the platform.
Remarks The element SF1-6 is not badly preserved and it shows clearly the prominent carina joining the parapet in the middle of the platform.
Range Lower Bashkirian (Chokierian-Kinderscoutian Western Europe regional substages: Higgins 1975). The Declinognathodus inaequalis first appearance datum (FAD) should be used for the recognition of the mid-Carboniferous boundary according to Sanz-López et al. (2006, 2013), because the specimens from the Global Boundary Stratotype Section and Point (GSSP) at the Arrow Canyon in Nevada illustrated in Brenckle et al. (1997) seem to be D. inaequalis (Higgins 1975). The latter is the nominal species for the first Pennsylvanian conodont biozone, the Declinognathodus inaequalis Zone, which was defined in the deep-water successions of Eurasia (Sanz-López et al. 2013 and references therein).
Declinognathodus praenoduliferous Nigmadganov and Nemirovskaya 1992
Figs. 7.3, 7.7
*1992 Declinognathodus praenoduliferous Nigmadganov and Nemirovskaya, pl. 2, fig. 6–9, pl. 3, fig. 2.
2011 Declinognathodus praenoduliferous. (Nemyrovska et al., pl.4, figs. 1–3, 5, 9–11, 15, 18, 20).
2013 Declinognathodus praenoduliferous. (Sanz-López and Blanco-Ferrera, fig. 5P–S).
2013 Declinognathodus praenoduliferous. (Sanz-López et al., fig. 4L).
Material Two P1 elements.
Diagnosis P1 element is characterized by a short carina and a platform covered by separate, widely transverse ridges and lack of a long, medial trough; only a split may occur at the most ventral part.
Remarks The studied elements 15A37-1 (Fig. 7.3) and 15A37-6 (Fig. 7.7) assigned to this species are very badly preserved. Nevertheless, they have a short carina and their platform looks as it is totally covered with transverse nodes and there is no trough.
Range Lowermost Bashkirian (Nemyrovska et al. 2011) and Upper Serpukhovian–Lower Bashkirian (Sanz-López et al. 2013).
Discussion
Biofacies and age of the Bodurria conodonts
The biofacies study of the Bodurria fauna is not very reliable because of the scarcity of conodonts. However, the relative abundance of Declinognathodus in the samples studied may indicate offshore but shallow-marine conditions, as defined in the Declinognathodus/Idiognathoides Biofacies of the shallow Mid-Continental Sea in the USA (Davis and Webster 1985). Nonetheless, this is not conclusive, because early forms of Declinognathodus have also been reported from deeper water facies (Nigmadganov and Nemirovskaya 1992; Nemirovskaya and Nigmadganov 1994; Sanz-López and Blanco-Ferrera 2009). In fact, during the latest Mississippian, Declinognathodus evolved from Gnathodus (Nemirovskaya and Nigmadganov 1994; Nemyrovska et al. 2011), which is a typical deep-water genus (Gnathodus Biofacies: Dreesen et al. 1986), although some Gnathodus species seem to have a shallower distribution (Orchard 1991) before their extinction at the beginning of the Pennsylvanian.
Lane et al. (1999) characterized Declinognathodus noduliferous as a cosmopolitan group that appears in most marine environments, allowing global correlations. These authors proposed the D. noduliferous s.l. FAD as the worldwide index for the mid-Carboniferous boundary GSSP. At the time of this definition, this species included several subspecies, such as D. noduliferous noduliferous (Ellison and Graves 1941), D. n. inaequalis (Higgins 1975), D. n. lateralis (Higgins and Bouckaert 1968), and D. n. japonicus (Igo and Koike 1964), which some authors now consider as independent species (Mizuno 1997; Sanz-López et al. 2006, 2013). According to Sanz-López et al. (2006), the D. n. inaequalis FAD should be used as the mid-Carboniferous boundary marker. Actually, the inaequalis Zone (Sanz-López et al. 2013) is the first Pennsylvanian conodont biozone in deep-water successions of northern Spain and Eurasia.
Gnathodus and Lochriea are commonly found in Visean-Serpukhovian beds of other Iberian sites (Nemyrovska et al. 2011; Sanz-López et al. 2013) and of the Maláguide Complex of the Betic Cordillera (Rodríguez-Cañero 1993; Rodríguez-Cañero and Guerra-Merchán 1996; O’Dogherty et al. 2000; Navas-Parejo 2012; Navas-Parejo et al. 2015). Nevertheless, some authors report the survival of some Gnathodus and Lochriea species in deep- and shallow-water lowermost Pennsylvanian beds of the Eurasian province and of the Palaeotethys (Higgins 1981; Sanz-López et al. 2006, and references therein). Therefore, despite its very low diversity and paucity of conodont elements, the association found in the Bodurria Unit indicates the earliest Bashkirian, when Gnathodus and Lochriea went extinct and other new species belonging to Upper Carboniferous had not yet appeared. According to the presence of the index species, the NFC rocks studied are assigned to the D. inaequalis Zone.
The same conodont association as that found in the Bodurria Unit is also present in the Barcaliente Formation (Wagner et al. 1971) of the Cantabrian Zone, northern Spain: a well-bedded black laminated limestone with high organic matter content, commonly shaly and laminated (González Lastra 1978). Nemyrovska et al. (2011) and Sanz-López and Blanco-Ferrera (2013, Appendix S1) provide information on several samples from Barcaliente Formation containing scarce D. bernesgae, D. inaequalis, and D. praenoduliferous, with exactly the same fauna as those found here. A very similar fauna was also reported from the western Pyrenees (Iraty Formation: Sanz-López and Blanco-Ferrera 2012). The Bodurria association also allows correlation with one conodont-bearing Bashkirian succession recently discovered in the Maláguide Complex (Cortijo de los González: Navas-Parejo 2012; Navas-Parejo et al. 2012). It is characterized by Idiognathoides corrugatus (Harris and Hollingsworth 1933), Declinognathodus noduliferous (Ellison and Graves 1941), and Lochriea aff. commutata (Branson and Mehl 1941), which probably were morphotypes transitional to D. orphanus (Van den Boogaard and Bless 1985).
Depositional environment of the Bodurria succession
Lithological evidence indicates that the sedimentary protoliths of the Bodurria succession were fine-grained siliciclastics very rich in organic matter (black shales), associated with fine-grained carbonates with low-energy current structures (parallel lamination and, occasionally, cross lamination). This suggests anoxic, moderately deep, and calm depositional environments, sporadically affected by weak bottom currents. The conodont fauna supports deposition of the rocks studied in distal open marine and moderately deep settings.
Upwards in the succession, the steadily more abundant psammitic beds become predominant, being the evidence of a gradual increase in siliciclastic supply (fine-to-medium-grained, rarely coarse-grained sand) under progressively higher energy hydrodynamic conditions, and deposition with higher sedimentary rate and in shallower environments than for the lower beds. The common presence of sedimentary structures in the psammites suggests sedimentation under higher energy bottom currents, acting episodically within quartz-rich sandy depositional systems. These were probably marine clastic wedges prograding from shallower marine regions towards the deeper basin interior, thus producing gradually thickening upwards arenite bodies interfingering with black shales, with local thinning-upwards trends probably related to lateral migration of feeding channels.
The above-mentioned data, together with available stratigraphic information on the NFC (Lafuste and Pavillon 1976; Gómez-Pugnaire et al. 1982; Laborda-Lopez et al. 2015a, b), allow an interpretation of the NFC pre-Mesozoic successions as deposited in marine, mainly terrigenous and subordinately carbonatic environments from the latest Precambrian to the late Palaeozoic. They also offer a glimpse of certain features of the Betic Palaeozoic palaeogeographic zoning: (1) in the eastern NFC areas (Lomo de Bas) shallow-marine platforms above the storm base level existed during the Devonian; (2) in the Sierra de Baza, the sedimentation accumulated in distal and deeper, calm, poorly oxygenated-to-anoxic environments at the beginning of the late Carboniferous.
Correlations with other Variscan terranes
It is worth mentioning that the Bashkirian conodonts found in the Maláguide Complex (Cortijo de los González) come from a different lithological succession to that of the Bodurria Unit. The Cortijo de los González section is much thinner as it is formed by only a few metres thick, strongly folded, bluish beds of recrystallized limestones locally including abundant crinoids and alternating with brownish-grey calcareous slates (Navas-Parejo et al. 2012). On the other hand, quartzitic sandstones do not appear anywhere in this succession or in other Maláguide Carboniferous outcrops; and both the pelites and the carbonates of this succession are much less rich in organic matter than their Bodurria counterparts. In addition, in the Maláguide Complex, there are also remnants of Bashkirian shallow-marine carbonate platforms, revealing a yet diversified palaeogeography in this realm during the Late Carboniferous (Herbig 1989). These remnants are limestone boulders, bearing abundant shallow-water faunas and floras, which are systematically included within the youngest Maláguide Palaeozoic formation, the Marbella Conglomerate (Geel 1973; Buchroithner et al. 1980; Herbig 1983, 1984, 1986; Herbig and Mamet 1985; Mamet and Herbig 1990). This platform was totally dismantled and the clasts redeposited in the synorogenic, deep clastic Maláguide basin later during the Late Carboniferous (Herbig 1984). All these facts suggest deposition of the Maláguide Bashkirian succession in more distal depositional settings and less restricted to ocean circulation than those where the Bodurria succession was deposited.
The thick Upper Carboniferous succession studied in the Bodurria Unit, with black pelites including subordinate conodont-bearing carbonate horizons both very rich in organic matter, associated with thick quartzitic sandstones, and showing moderately distal deep marine facies and open marine fauna, represents the first robust stratigraphic support for identifying the tectonostratigraphic and palaeogeographic affinities of at least part of the NFC, to external tectonostratigraphic terranes of the Iberian Massif, Palentian Domain of the Cantabrian Zone, in particular (Wagner et al. 1971; González Lastra 1978; Nemyrovska et al. 2011). This hypothesis is further supported by the absence of marine Upper Carboniferous sediments in the Central Iberian, Galicia Tras-Os-Montes, and Ossa Morena zones (Martínez Catalán 2011, 2012; Simancas et al. 2009). The proposed correlation (Fig. 8) indicates that the boundary between the Variscan Iberian palaeogeographic zones can be traced south-eastwards, below the Alpine deformational front, and provides a reasonable tectonopalaeogeographic explanation for the absence of the NFC in the western sector of the Betic Internal Zones.
The new stratigraphic data provided herein confirm the Palaeotethyan (instead of Rheic) palaeogeographic affinities for the NFC as previously proposed for other Betic Palaeozoic terranes, the Maláguide Complex, in particular (cf. Rodríguez-Cañero and Martín-Algarra 2014). The Late Carboniferous palaeogeographic reconstruction presented in Fig. 8 illustrates the possible relationships of the NFC with other elements of the Variscan belt. The geology of Western Europe is modified from Martínez Catalán (2011) to include the Y geometry of the foredeep and foreland thrust belt proposed by Matte (2001). In this model, the NFC is located at the southern end of the Variscan foredeep basin (to the SE of the Cantabrian Zone) and the Maláguide Complex to the E of it, in the same margin but in a more distal position and closer to Alpine areas. Our main reason is that the Maláguide conodont-bearing successions show stronger palaeobiogeographic and lithostratigraphic affinities with the Alps than with Iberia since, at least, the Late Ordovician (Rodríguez-Cañero et al. 2010).

Early Bashkirian eastern Variscan belt, modified from Martínez Catalán (2011), with the hypothetical location of the Nevado-Filábride Complex with respect to other Variscan Iberian Terranes and of the position of the localities bearing lowermost Bashkirian conodonts mentioned in the discussion. Zones of the Variscan Chain: CIZ Central Iberian, CZ Cantabrian, GTMZ Galicia-Trás-os-Montes, MGCZ Mid-German Crystalline, MZ Moldanubian, OMZ Ossa-Morena, RHZ RhenoHercynian, SPZ South-Portuguese, STZ Saxo-Thuringian, TBZ Teplá-Barrandian, WALZ West Asturian-Leonese
Conclusions
The main conclusion of this paper are summarized as it follows:
-
1.
The first conodonts found in the Bodurria Unit allow the first precise biochronostratigraphic dating of a Palaeozoic succession in the Nevado-Filábride Complex, which includes Upper Carboniferous beds, in particular, the lower Bashkirian, Declinognathodus inaequalis Zone.
-
2.
The age of the Bodurria succession demonstrates that the Variscan deformation of the lowest Nevado-Filábride Complex took place in late Carboniferous time, as in the outermost sectors of the Iberian Variscan Massif or in Palaeotethys-related areas to the E of it now found within the Alpine Chain.
-
3.
The lithostratigraphic features and conodont content of the Bodurria succession indicate its deposition in open marine, moderately deep anoxic environments, very similar to those recognized in the Palentian Domain of the Cantabrian Zone during the Bashkirian.
-
4.
The proposed correlation provides strong support to the classic idea that attributes the NFC to Iberia-Europe. It also demonstrates that at least some tectonopalaeogeographic zones of the Variscan Iberian Massif can be traced to the Betic Cordillera below the Alpine deformational front.
-
5.
This model also allows explaining why the Nevado-Filábride Complex is absent in the Western Betic Cordillera and in the Rif, and why the pre-Mesozoic basements of the Alborán Domain units (Alpujárride and Maláguide complexes) thrust directly onto the western Subbetic units of the South-Iberian Palaeomargin.
-
6.
Finally, this new evidence provides further support to exclude the Nevado-Filábride Complex from the Alborán Domain concept and to restrict it, essentially, to the Alpujárride and Maláguide complexes and related covers (Frontal Units) after their main Alpine orogenic deformation during the Aquitanian–Burdigalian (cf. Guerrera et al. 1993; Gómez Pugnaire et al. 2012).
References
Álvarez-Lobato F (1987) Subhorizontal shear zones and their relation to nappe movements in the Cantal and Miñarros units, Eastern Betic Zone (Spain). Geol Mijnbouw 66:101–111
Álvarez-Lobato F, Aldaya F (1985) Las unidades de la Zona Bética en la región de Águilas-Mazarrón (Prov. de Murcia). Estud Geol 41:139–146
Anderson MA, Dargan G, Brock GA, Talent JA, Mawson R (1995) Maximising efficiency of conodont separations using sodium polytungstate solution. Cour Forsh Senck 182:515–521
Balanyá JC, García-Dueñas V (1987) Les directions structurales dans le Domaine d’Alborán de part et d’autre du Détroit de Gibraltar. CR Acad Sci 304:929–932
Branson EB (1938) Stratigraphy and paleontology of the lower Mississippian of Missouri, Pt. 1. Univ Mo Stud 13:1–208
Branson EB, Mehl MG (1941) New and little known Carboniferous genera. J Paleontol 15:97–106
Brenckle PL, Baesemann JF, Lane HR, West RR, Webster GD, Langenheim RL, Brand U, Richards BC (1997) Arrow Canyon, the Mid-Carboniferous boundary stratotype. In: Brenckle PL, Page WR (eds) Guidebook: arrow Canyon range, Nevada. Cushman Foundation for foraminifera research, 36 Suppl. Special Publication, Washington, pp 13–32
Buchroithner MF, Flügel E, Flügel HW, Stattegger K (1980) Mikrofazies, Fossilien und Herkunft der Kalk-Gerölle im Karbon-"Flysch” der Betischen Kordilleren. Span Facies 2:1–54
Cocks LRM, Torsvik TH (2006) European geography in a global context from the Vendian to the end of the Palaeozoic. In: Gee DG, Stephenson RA (eds) European lithosphere dynamics, Memoirs 32. Geological Society, London, pp 83–95
Davis LE, Webster GD (1985) Late Mississippian to Early Pennsylvanian conodont biofacies in central Montana. Lethaia 18:67–72
Dreesen R, Sandberg C, Ziegler W (1986) Review of late Devonian and early Carboniferous conodont biostratigraphy and biofacies models as applied to the Ardenne shelf. Ann Soc Géol Belg 109:27–42
Dunn DL (1966) New Pennsylvanian platform conodonts from southwestern United States. J Paleontol 40:1294–1303
Dzik J (1976) Remarks on the evolution of Ordovician conodonts. Acta Palaeontol Pol 21(4):395–458
Ellison S, Graves RW Jr (1941) Lower Pennsylvanian (Dimple Limestone) conodonts of the Marathon region, Texas. Mo Univ Sch Mines Metall Bull 14:1–21
Epstein AG, Epstein JB, Harris LD (1977) Conodont color alteration—an index to organic metamorphism. Geol Surv Prof Pap 995:1–27
Franke W, Cocks LRM, Torsvik TH (2017) The Palaeozoic Variscan oceans revisited. Gondwana Res 48:257–284
Geel T (1973) The geology of the Betic of Malaga, the Subbetic and the zone between these two units in the Velez Rubio area (Southern, Spain). GUA Pap Geol 5:179
Gómez-Pugnaire MT (1981) Evolución del metamorfismo alpino en el Complejo Nevado-Filábride de la Sierra de Baza (Cordilleras Béticas, España). Tecniterrae 4:130
Gómez-Pugnaire MT, Franz G (1988) Metamorphic evolution of the Paleozoic series of the Betic Cordilleras (Nevado-Filabride Complex, SE Spain) and its relationship with the Alpine orogeny. Geol Rundsch 77:619–640
Gómez-Pugnaire MT, Chacón J, Mitrofanov V, Timofeev V (1982) First report on the Precambrian rocks in the graphite-bearing series of the Nevado-Filábride Complex (Betic Cordilleras, Spain). Neues Jahrb Geol P M 3:176–180
Gómez-Pugnaire MT, Azor A, López Sánchez-Vizcaíno V, Soler M (2003) The amphibolites from the Ossa-Morena/Central Iberian Variscan suture (southwestern Iberian Massif): geochemistry and tectonic interpretation. Lithos 68:23–42
Gómez-Pugnaire MT, Galindo-Zaldívar J, Rubatto D, González-Lodeiro F, López Sánchez-Vizcaíno V, Jabaloy A (2004) A reinterpretation of the Nevado-Filábride and Alpujárride complex (Betic Cordillera): field, petrography and U–Pb ages from orthogneisses western Sierra Nevada, S Spain). Schweiz Mineral Petrogr Mitt 84:303–322
Gómez-Pugnaire MT, Rubatto D, Fernández-Soler JM, Jabaloy A, López Sánchez-Vizcaíno V, González-Lodeiro F, Galindo-Zaldívar J, Padrón-Navarta JA (2012) U–Pb geochronology of Nevado-Filábride gneisses: evidence for the Variscan nature of the deepest Betic complex (SE Spain). Lithos 146–147:93–111
González Lastra J (1978) Facies Salinas en la Caliza de Montaña (Cordillera Cantábrica). Trabajos de Geología 10:249–265
Graves RW, Ellison S (1941) Ordovician conodonts of the Marrathon Basin, Texas. Missouri University, School of Mines and Metallurgy. Bull Tech Ser 14:1–26
Grayson RC Jr, Merrill GK, Lambert LL (1990) Carboniferous gnathodontid conodont apparatuses: evidence of a dual origin for Pennsylvanian taxa. Cour Forsh Senck 118:353–396
Guerrera F, Martín-Algarra A, Perrone V (1993) Late Oligocene–Miocene syn-late-orogenic successions in Western and Central Mediterranean Chains from Betic Cordillera to Southern Apennine). Terra Nova 5:525–544
Harris RW, Hollingsworth RV (1933) New Pennsylvanian conodonts from Oklahoma. Am J Sci Ser 5 25(147):193–204
Herbig HG (1983) El Carbonífero de las Cordilleras Béticas. In: Martínez-Díaz C (ed) Carbonífero y Pérmico de España, X Congreso Internacional sobre la Estratigrafía y la Geología del Carbonífero, Instituto Geológico y Minero de España, Madrid, pp 343–356
Herbig HG (1984) Rekonstruktion eines nicht mehr existenten Sedimentations-Raums: die Kalk-Gerölle im Karbon Flysch der Malagiden (Betische Kordillere, Südspanien). Facies 11:1–108
Herbig HG (1986) Rugosa und Heterocorallia aus Obervisé-Geröllen der Marbella-Formation (Betische Kordillere, Südspanien). Palaeontol Z 60:189–225
Herbig HG (1989) Carboniferous paleogeography of the West-Mediterranean Paleotethys. In: 11th international congress on stratigraphy and geology of the carboniferous 4, pp 186–196
Herbig HG, Mamet B (1985) Stratigraphy of the limestone boulders, Marbella Formation (Betic Cordillera, Southern Spain). In: Comptes rendus 10ème Congrès Internationale sur la Stratigraphie et la Géologie du Carbonifère, Madrid 1983, 1, pp 199–212
Higgins AC (1975) Conodont zonation of the late Viséan-early Westphalian strata of the south and central Pennines of northern England. Bull Geol Surv Great Br 53:1–127
Higgins AC (1981) The distribution of conodonts in relation to the paleogeography of late Visean-Namurian time. In: Neale JW, Brasier MD (eds) Microfossils from recent fossil shelf seas. Ellis Horwood Limited, Chichester, pp 37–51
Higgins AC, Bouckaert J (1968) Conodont stratigraphy and palentology of the Namurian of Belgium. Mémoires Expl. Cartes geol. Minie Belgique 10:1–64
Hsü KJ, Bernouilli D (1978) Genesis of the Tethys and the mediterranean. In: Hsü KJ, Montadert L et al (eds) Init Rep DSDP, 42, Pt. 1. U.S. Gov. Printing Office, Washington, pp 943–949
Igo H, Koike T (1964) Carboniferous conodonts from the Omi Limestone, Niigata Prefecture, Central Japan/Studies of Asian conodonts, part I. Trans Proc Palaeont Soc Jpn 53:26–30
Jabaloy A (1993) La estructura de la región occidental de la Sierra de los Filabres (Cordilleras Béticas). Tierras del Sur, Spain, vol 9. Universidad de Granada, Granada, p 261
Jabaloy A, Gómez-Pugnaire MT, Padrón-Navarta JA, López Sánchez-Vizcaíno V, Garrido CJ (2015) Subduction- and exhumation-related structures preserved in metaserpentinites and associated metasediments from the Nevado-Filábride complex (Betic Cordillera, SE Spain). Tectonophysics 644–645:40–57
Jeppsson L, Anehus R, Fredholm D (1999) The optimal acetate buffered acetic acid technique for extracting phosphatic fossils. J Paleontol 73:964–972
Julivert M, Fontboté M, Ribeiro A, Conde LE (1972) Mapa y memoria explicativa del Mapa Tectónico de la Península Ibérica y Baleares, scale 1:1,000,000. Instituto Geológico y Minero de España, Madrid, p 113
Kroner U, Romer RL (2013) Two plates-many subduction zones: the Variscan orogeny reconsidered. Gondwana Res 24:298–329
Laborda-López C, Aguirre J, Donovan SK (2013) Asociaciones de macrofósiles en rocas metamórficas del Complejo Nevado-Filábride (Zonas Internas de la Cordillera Bética) en Águilas, Murcia (SE España). Tafonomía y biocronoestratigrafía. XXIX Jornadas de Paleontología, Abstracts, pp 83–84
Laborda-López C, Aguirre J, Donovan SK (2015a) Surviving metamorphism: taphonomy of fossil assemblages in marble and calc-silicate schist. Palaios 30:668–679
Laborda-López C, Aguirre J, Donovan SK, Navas-Parejo P, Rodríguez S (2015b) Fossil assemblages and biochronology of metamorphic carbonates of the Nevado-Filábride complex from the Águilas tectonic arc (SE Spain). Span J Palaeontol 30(2):275–292
Lafuste MLJ, Pavillon MJ (1976) Mise en évidence d’Eifélien daté au sein des terrains métamorphiques des zones internes des Cordillères bétiques. Intérêt de ce nouveau repère stratigraphique. CR Acad Sci 283:1015–1018
Lane HR (1967) Uppermost Mississippian and Lower Pennsylvanian conodonts from the type Morrowan region Arkansas. J Paleontol 41:920–943
Lane HR (1968) Symmetry in conodont element pairs. J Paleontol 42:1258–1263
Lane HR, Straka JJ (1974) Late Mississippian and early Pennsylvanian conodonts, Arkansas and Oklahoma. Geol Soc Am Spec Pap 152:144
Lane HR, Brenckle PL, Baesemann JF, Richards B (1999) The IUGS boundary in the middle of the Carboniferous: arrow Canyon, Nevada, USA. Episodes 22(4):272–283
López Sánchez-Vizcaíno V, Rubatto D, Gómez-Pugnaire MT, Tommsdorff V, Müntener O (2001) Middle Miocene high-pressure metamorphism and fast exhumation of the Nevado-Filábride complex, SE Spain, Terra Nova 13:327–332
Lotze F (1945) Zur gliederung der Varisziden der lberischen Meseta. Geotekt Forsch 6:78–92
Mamet BL, Herbig HG (1990) The algae Pseudodonezella n. gen. and Eovelebitella occitanica Vachard, 1974 from Southern Spain (Carboniferous, Betic Cordillera. Revista Española de Micropaleontología 22:199–211
Martínez Catalán JR (2011) Are the oroclines of the Variscan belt related to late Variscan strike-slip tectonics? Terra Nova 23(4):241–247
Martínez Catalán JR (2012) The Central Iberian arc, an orocline centered in the Iberian Massif and some implications for the Variscan belt. Int J Earth Sci 101:1299–1314
Martínez Catalán JR, Arenas R, Díaz-García F, Rubio-Pascual FJ, Abati J, Marquínez J (1996) Variscan exhumation of a subducted Paleozoic continental margin: the basal units of the Ordenes Complex, Galicia, NW Spain. Tectonics 15:106–121
Martínez Catalán JR, Arenas R, Díaz-García F, Abati J (1997) Variscan accretionary complex of northwest Iberia: Terrane correlation and succession of tectonothermal events. Geology 25:103–110
Martínez Catalán JR, Arenas R, Díaz-García F, Gómez-Barreiro J, González-Cuadra P, Abati J, Castiñeiras P, Fernández-Suárez J, Sánchez-Martínez S, Andonaegui P, Gónzalez-Clavijo E, Díez-Montes A, Rubio-Pascual FJ, Valle-Aguado B (2007) Space and time in the tectonic evolution of the northwestern Iberian Massif. Implications for the Variscan belt. In: Hatcher RD Jr, Carlson MP, McBride JH, Martínez Catalán JR (eds) 4–D framework of continental crust. Geological Society of America, Memoir 200, Boulder, Colorado, pp 403–423
Martínez-Martínez JM (1986) Evolución tectono-metamórfica del Complejo Nevado-Filábride en el sector entre Sierra Nevada y Sierra de los Filabres (Cordilleras Béticas). Cuadernos de Geología de la Universidad de Granada 13:1–194
Matte P (2001) The Variscan collage and orogeny (480–290 Ma) and the tectonic definition of the Armorica microplate: a review. Terra Nova 13:122–128
Michard A, Hoepffner C, Soulaimani A, Baidder L (2008) The Variscan belt. In: Michard A, Saddiqui O, Chalouan A, Frizon de Lamotte D (eds), Continental evolution: the geology of Morocco. Springer, Berlin, pp 65–132
Mizuno Y (1997) Conodont faunas across the Mid-Carboniferous boundary in the Hina Limestone, Southwest Japan. Paleontol Res 1:237–259
Murphy JB, Gutierrez-Alonso G, Nance RD, Fernandez-Suarez J, Keppie JD, Quesada C, Strachan J, Dostal J (2006) Origin of the Rheic Ocean: rifting along a Neoproterozoic suture? Geology 34:325–328
Murphy JB, Gutierrez-Alonso G, Nance RD, Fernandez-Suarez J, Keppie JD, Quesada C, Dostal J, Braid JA (2009) Rheic Ocean mafic complexes: overview and synthesis. Geol Soc Lond Spec Publ 327:343–369
Nance RD, Gutiérrez-Alonso G, Keppie JD, Linnemann U, Murphy JB, Quesada C, Strachan RA, Woodcock NH (2010) Evolution of the Rheic Ocean. Gondwana Res 17:194–222
Navas-Parejo P (2012) Paleozoic stratigraphy and palaeogeography of the Malaguide Complex (Betic Cordillera) and other Western Mediterranean related domains (Calabria–Peloritani Terrane). Ph.D. Thesis, University of Granada, p 235. http://hdl.handle.net/10481/23780. Accessed 12 Dec 2017
Navas-Parejo P, Somma R, Martín-Algarra A, Perrone V, Rodríguez-Cañero R (2009a) First record of Devonian Orthoceratid-bearing limestones in Southern Calabria (Italy). CR Palevol 8:365–373
Navas-Parejo P, Rodríguez-Cañero R, Somma R, Martín-Algarra A, Perrone V (2009b) The Frasnian Upper Kellwasser event and a lower Famennian stratigraphic gap in Calabria (southern Italy). Palaeobiodivers Palaeoenviron 89:111–118
Navas-Parejo P, Rodríguez-Cañero R, Martín-Algarra A (2012) Primer registro de un horizonte estratigráfico hemipelágico con conodontos del Carbonífero Superior en el Complejo Maláguide (Cordillera Bética oriental). Geogaceta 52:81–84
Navas-Parejo P, Rodríguez-Cañero R, Martín-Algarra A (2015) New conodont data from a Devonian-Carboniferous succession in the central sector of the Betic Cordillera (SE Spain). Span J Palaeontol 30:133–145
Nemirovskaya TI, Nigmadganov I (1994) The Mid-Carboniferous conodont event. Cour Forsh Senck 168:319–333
Nemyrovska TI, Wagner RH, Winkler-Prins CF, Montañez I (2011) Conodont faunas across the mid-Carboniferous boundary from the Barcaliente Formation at La Lastra (Palentian Zone, Cantabrian Mountains, northwest Spain); geological setting, sedimentological characters and faunal descriptions. Scripta Geol 143:127–183
Nigmadganov IM, Nemyrovskaya TI (1992) Mid-Carboniferous boundary conodonts from the Gissar Ridge, South Tienshan, Middle Asia. Cour Forsh Senck 154:253–275
O’Dogherty L, Rodríguez-Cañero R, Gursky H-J, Martín-Algarra A, Caridroit M (2000) New data on lower Carboniferous stratigraphy and palaeogeography of the Malaguide Complex (Betic Cordillera, Southern Spain). CR Acad Sci IIa 331:533–541
Orchard MJ (1991) Conodonts, time and terranes: an overview of the biostratigraphic record in the western Canadian Cordillera. In: Orchard MJ, McCracken AD (eds) Ordovician to Triassic Conodont paleontology of the Canadian Cordillera, vol 417. Geological Survey of Canada, Bulletin, Ottawa, pp 1–25
Pastor-Galán D, Martín-Merino G, Corrochano D (2014) Timing and structural evolution in the limb of an orocline: The Pisuerga–Carrión Unit (southern limb of the Cantabrian Orocline, NW Spain). Tectonophysics 622:110–121
Pereira MF, Apraiz A, Chichorro M, Silva JB, Armstrong RA (2010) Exhumation of high pressure rocks in northern Gondwana during the Early Carboniferous (Coimbra–Cordoba shear zone, SW Iberian Massif): tectonothermal analysis and U–Th–Pb SHRIMP in-situ zircon geochronology. Gondwana Res 17(2–3):440–460
Pereira MF, Chichorro M, Johnston ST, Gutiérrez-Alonso G, Silva JB, Linnemann U, Hofmann M, Drost K (2012) The missing Rheic Ocean magmatic arcs: provenance analysis of Late Paleozoic sedimentary clastic rocks of SW Iberia. Gondwana Res 22:882–891
Pereira MF, Castro A, Fernández C (2015) The inception of a Paleotethyan magmatic arc in Iberia. Geosci Front 6:297–306
Pérez-Estaún A, Bastida F, Alonso JL, Marquínez J, Aller J, Álvarez-Marrón J, Marcos A, Pulgar JA (1988) A thin-skinned tectonic model for an arcuate fold and thrust belt: the Cantabrian Zone (Variscan Ibero–Armorican Arc). Tectonics 7:517–537
Puga E, Fontboté JM, Martín Vivaldi JL (1975) Kyanite pseudomorphs after andalusite in polymetamorphic rocks of the Sierra Nevada (Betic Cordillera, Southern Spain). Schweiz Mineral Petrogr Mitt 55:227–241
Puga E, Díaz de Federico A, Nieto JM (2002) Tectonostratigraphic subdivision and petrological characterisation of the deepest complexes of the Betic zone: a review. Geodin Acta 15:23–43
Puga E, Fanning M, Díaz de Federico A, Nieto JM, Beccaluva L, Bianchini G, Díaz Puga MA (2011) Petrology, geochemistry and U–Pb geochronology of the Betic Ophiolites: inferences for Pangaea break-up and birth of the westernmost Tethys Ocean. Lithos 124:255–272
Puga E, Díaz de Federico A, Fanning M, Nieto JM, Rodríguez Martínez-Conde JA, Díaz Puga MA, Bianchini G, Natali C, Beccaluva L (2017) The Betic Ophiolites and the mesozoic evolution of the western tethys. Geosciences 7:31
Rejebian VA, Harris AO, Huebner JS (1987) Conodont color and textural alteration: an index to regional metamorphism, contact metamorphism, and hydrotermal alteration. Geol Soc Am Bull 99:471–479
Robardet M (2003) The Armorica ‘microplate’: fact or fiction? Critical review of the concept and contradictory palaeobiogeographical data. Palaeogeogr Palaeoclimatol Palaeoecol 195:125–148
Rodríguez-Cañero R (1993) Contribución al estudio de los conodontos del Paleozoico del Complejo Maláguide (Cordillera Bética). Ph.D. Thesis, University of Málaga, Spain, pp 1–474
Rodríguez-Cañero R, Guerra Merchan A (1996) Nuevos datos sobre la fauna de conodontos y la edad de la Formación Falcoña (Complejo Maláguide, Cordillera Bética, España). Revista de la Sociedad Española de Paleontología 11:235–246
Rodríguez-Cañero R, Martín-Algarra A (2014) Frasnian/Famennian crisis in the Malaguide Complex (Betic Cordillera, Spain). Terra Nova 26:38–54
Rodríguez-Cañero R, Martín-Algarra A, Sarmiento GN, Navas-Parejo P (2010) First Late Ordovician conodont fauna in the Betic Cordillera (South Spain): a palaeobiogeographical contribution. Terra Nova 22:330–340
Sanz-López J, Blanco-Ferrera S (2009) Probable presence of old species of Declinognathodus in the Mississippian and the correlation with the Mid-Carboniferous boundary in the Cantabrian Mountains (Spain). Permophiles 53(Supplement 1):45–46
Sanz-López J, Blanco-Ferrera S (2012) Lower Bashkirian conodonts from the Iraty Formation in the Alduides-Quinto Real Massif (Pyrenees, Spain). Geobios 45:397–411
Sanz-López J, Blanco-Ferrera S (2013) Early evolution of Declinognathodus close to the Mid-Carboniferous Boundary interval in the Barcaliente type section (Spain). Palaeontology 56(5):927–946
Sanz-López J, Blanco-Ferrera S, García-López S, Sánchez de Posada LC (2006) The mid-Carboniferous boundary in northern Spain: difficulties for correlation of the global stratotype section and point. Riv Ital Paleontol Stratigr 112:3–22
Sanz-López J, Blanco-Ferrera S, Sánchez de Posada LC (2013) Conodont chronostratigraphical resolution and Declinognathodus evolution close to the Mid-Carboniferous Boundary in the Barcaliente Formation type section, NW Spain. Lethaia 46:438–453
Şengör AMC (1990) Plate tectonics and orogenic research after 25 years: a Tethyan perspective. Earth Sci Rev 27:1–201
Şengör AMC, Altiner D, Cin A, Ustaömer T, Hsü KJ (1988) Origin and assembly of the Tethyside orogenic collage at the expense of Gondwana Land. In: Audley-Charles MG, Hallam A (eds) Gondwana and Tethys. Geol Soc Lond Spec Pub, 37, London, pp 119–181
Shaw J, Johnston ST, Gutiérrez-Alonso G, Weil AB (2012) Oroclines of the Variscan orogen of Iberia: paleocurrent analysis and paleogeographic implications. Earth Planet Sci Lett 329–330:60–70
Simancas JF, Carbonell R, González-Lodeiro F, Pérez-Estaún A, Juhlin C, Ayarza P, Kashubin A, Azor A, Martínez-Poyatos D, Almodóvar GR, Pascual E, Sáez R, Expósito I (2003) The crustal structure of the transpressional Variscan Orogen of SW Iberia: the IBERSEIS deep seismic reflection profile. Tectonics 22(6):1062–1074
Simancas JF, Azor A, Martínez-Poyatos D, Tahiri A, El Hadi H, González-Lodeiro F, Pérez-Estaún A, Carbonell R (2009) Tectonic relationships of Southwest Iberia with the allochthons of Northwest Iberia and the Moroccan Variscides. C R Geosci 341:103–113
Simancas JF, Ayarza P, Azo A, Carbonell R, Martínez-Poyatos D, Pérez-Estaún A, González-Lodeiro F (2013) A seismic geotraverse across the Iberian Variscides: orogenic shortening, collisional magmatism, and orocline development. Tectonics 32:417–432
Stampfli GM (2000) Tethyan oceans. In: Bozkurt E, Winchester JA, Piper JDA (eds) Tectonics and magmatism in Turkey and the surrounding area. Geol Soc Lond Spec Publ 173, London, pp 1–23
Stampfli GM, Borel GD (2002) A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrons. Earth Planet Sci Lett 196:17–33
Stampfli GM, Borel GD (2004) The TRANSMED transects in space and time: constraints on the paleotectonic evolution of the mediterranean domain. In: Cavazza W, Roure FM, Spakman W, Stampfli GM, Ziegler PA (eds) The TRANSMED Atlas: the mediterranean region from crust to mantle. Springer, Berlin, pp 53–80
Stampfli GM, Kozur H (2006) Europe from the Variscan to the Alpine cycles. In: Gee DG, Stephenson R (eds) European lithosphere dynamics, Memoir, vol 32. Geological Society, London, pp 57–82
Stampfli GM, Vavassis I, De Bono A, Rosselet F, Matti B, Bellini M (2003) Remnants of the Paleotethys oceanic suture-zone in the western Tethys area. Bollettino della Società Geologica Italiana, Volume Speciale 2:1–23
Stampfli GM, Hochard C, Vérard C, Wilhem C, Von Raumer JF (2013) The formation of Pangea. Tectonophysics 593:1–19
Van den Boogaard M, Bless MJM (1985) Some conodont fauna from the Aegiranum Marine Band. PK Ned Akad B Phys 88(2):133–154
Vera JA (ed) (2004) Geología de España. SGE-IGME, Madrid
Von Raumer JF, Stampfli GM, Borel G, Bussy F (2002) The organization of pre-Variscan basement areas at the Gondwana margin. Int J Earth Sci 91:35–52
Von Raumer JF, Stampfli GM, Bussy F (2003) Gondwana-derived microcontinents—the constituents of the Variscan and Alpine collisional orogens. Tectonophysics 365:7–22
Wagner RH, Winkler Prins CF, Riding RE (1971) Lithostratigraphic units of the lower part of the Carboniferous in northern Leon, Spain. Trabajos de Geología 4:603–663
Weil AB, Van der Voo R, van der Pluijm BA, Parés JM (2000) The formation of an orocline by multiphase deformation: a paleomagnetic investigation of the Cantabria–Asturias Arc (northern Spain). J Struct Geol 22:735–756
Acknowledgements
We acknowledge J.R. Martínez Catalán and an anonymous reviewer for their comments and suggestions that notably improved the paper. This work is supported by Grants CGL2016-75679P, CGL2016-75224-R, and CGL2015-71692-P (MINECO/FEDER, Spain) and RNM-208 (Junta de Andalucía, Spain). We acknowledge the help of Ángela Cabrera-Porras and Ana Vélez-Felipe for their help during laboratory work for conodont sample preparation, and Isabel Sánchez-Almazo (C.I.C.-UGR) for the assistance during the SEM study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Rodríguez-Cañero, R., Jabaloy-Sánchez, A., Navas-Parejo, P. et al. Linking Palaeozoic palaeogeography of the Betic Cordillera to the Variscan Iberian Massif: new insight through the first conodonts of the Nevado-Filábride Complex. Int J Earth Sci (Geol Rundsch) 107, 1791–1806 (2018). https://doi.org/10.1007/s00531-017-1572-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00531-017-1572-8