
Abstract
An invasive forest pathogen, Cronartium ribicola, white pine blister rust (WPBR), is believed to have arrived in the Sacramento Mountains of south-central New Mexico about 1970. Epidemiological and genetic evidence supports the hypothesis that introduction was the result of long-distance dispersal (LDD) by atmospheric transport from California. This study applies a method to identify the atmospheric conditions favorable for rust transport and infection. An upper level synoptic classification (ULSC) identifies patterns of upper-level flow favorable for the transport of rust spores from a source to a target. Transport data are coupled with data for surface conditions favorable for infection at a designated target. A resulting calendar lists likelihood classes for establishment by four-times-daily observations during a dispersal season from April through July in the years 1965 to 1974. The single most-favorable period for transport and infection at the New Mexico site was identified as 1–15 June 1969. Five additional sites in the western United States with susceptible white pine populations and known infestation status were then evaluated to verify the model. Only the infested sites exhibit an establishment likelihood of “high” or “very high.” This suggests that the methodology correctly identifies locations with elevated establishment likelihood. Finally, likelihoods at nine additional points in the southwestern United States are determined and used to map regional patterns of transport, infection and establishment. The ULSC combined with appropriate surface meteorological data could be used to further investigate transport and infection, identify other areas at risk, assess the potential for gene flow of WPBR and evaluate long-distance dispersal of other pathogens.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Invasions of exotic species pose serious economic and ecological threats to managed and natural ecosystems worldwide (Mack et al. 2000). The 100-year, expanding epidemic of white pine blister rust (WPBR), caused by Cronartium ribicola J.C. Fisch., in North America demonstrates how an important plant pathogen spreads to new environments (Geils 2001; Samman et al. 2003). WPBR is native to Asia on white pines (Pinus, subgenus Strobus), ribes (currants and gooseberries, Ribes), and several other plant genera. Hunt (2003) concludes the rust was probably moved west by trade—first to Europe (circa 1850) and later (late 1800s through early 1900s) to numerous locations in North America. Although human activities mediated and then mitigated the original North American introductions, the rust continues to spread (presumably) by atmospheric transport (Geils et al. 2003). The potential for epidemic expansion to other vulnerable regions and for continued gene flow remains an unresolved threat (Hamelin et al. 2000).
Numerous attempts to quantify and model the expansion of invasive species are documented. Such expansion may occur as either the sum of many short steps (through local spread) or by a few great leaps [long-distance dispersal (LDD)]. The diffusion concept developed by Gregory in 1945 (see Waggoner and Aylor 2000) remains at the core of models proposed by Andow et al. (1990), Caswell et al. (2003) and others. This approach relies primarily upon the study of demography and dispersal gradients. Clark et al. (2003) and Muller-Landau et al. (2003) suggest that although LDD may be infrequent and of variable length it merits closer examination. For many pathogenic fungi, LDD contributes significantly to the establishment of disease outbreaks (Brown and Hovmoller 2002; Nagarajan and Singh 1990). The critical stages of transport and infection are (1) production of spores at a source and their escape from the canopy, (2) aerial transport and survival, and (3) deposition, germination, infection and reproduction on a suitable host (Aylor 1986; Aylor 2003; Isard et al. 2005). Mims and Mims (2004) report several recent examples of viable fungal spores transported long distances in smoke. Other examples for LDD of plant pathogens include coffee leaf rust (Bowden et al. 1971), peanut rust (Van Arsdel 1973), sugarcane rust (Purdy et al. 1985), blue mold (Davis and Monahan 1991), soybean blister rust (Munoz et al. 2004) and soybean rust (Isard et al. 2005). These studies examine aerial dispersal by air-flow patterns across continental or oceanic distances. Other approaches for the study of LDD include climate-matching (e.g., Yonow et al. 2004) and landscape genetics (e.g., Hamelin et al. 2000).
The spread of WPBR displays a range of spatial scales indicative of different frequencies and processes. Although the spores produced on ribes, which infect white pines, are easily killed by light, heat and desiccation, they are still capable of spreading the rust as far as several kilometers (Van Arsdel 1967; Van Arsdel et al. 2006). The thick-walled and pigmented spores produced on white pines, which infect ribes, often disperse tens of kilometers and are even capable of surviving for days while traveling hundreds of kilometers (Mielke 1943). Recently discovered disjunct outbreaks of WPBR suggest LDD to south-central New Mexico (Hawksworth 1990), western South Dakota (Lundquist et al. 1992), central North Dakota (Draper and Walla 1993), northeastern Nevada (Vogler and Charlet 2004), southern Colorado (Blodgett and Sullivan 2004), central Utah (B.W. Geils and D.R. Vogler, manuscript in preparation) and several additional locations in New Mexico (D. Conklin, personal communication). These outbreaks are in relatively unpopulated regions and suggest introduction by atmospheric rather than anthropogenic transport. The surveys of Smith and Hoffman (2000) and Geils et al. (2003) reveal that WPBR populations in interior western North America are widely scattered, and that not all mountain ranges with host populations have detectable levels of WPBR. The remote and incomplete distribution of WPBR in this region implies that meteorological conditions related to either transport or infection may limit LDD of the rust.
Various synoptic climatological approaches are used to explain epidemic introductions. Many approaches focus on either environmental suitability at the deposition site or on the pathway of aerial transport. For example, Yonow et al. (2004) identify the potential world-wide range of a plant pathogen by matching critical climatic variables at sites where the pathogen occurs to others with climatic similarity. Fujioka’s (1996) failure to account for the distribution of WPBR in Oregon and California from climatic variables, however, indicates such an approach can be inhibited by lack of data at appropriate spatial and/or temporal scales for either the climate or the pathogen. In contrast, Van Arsdel et al. (1961) is one of many examples where climate (and other factors) significantly explains how variation in environmental suitability determines rust distribution and severity. Holly Kearns (2005) used observations of climate and WPBR distribution from Wyoming (where the disease has occurred for decades) to predict its potential distribution in Colorado (where it has recently invaded). Aylor (2003) incorporates transport and survival along with suitability into dispersal models for several plant pathogens. He concludes that LDD may be more often limited by establishment at the deposition site than by conditions of transport.
Although the Sacramento Mountains of south-central New Mexico had long been recognized as highly vulnerable to WPBR (Bureau of Plant Industry 1921), its first discovery was reported by Hawksworth (1990) several decades after the rust was known in the nearest populations of susceptible pines in California, Idaho and Wyoming. Genetic analysis (Hamelin et al. 2000) concluded that the New Mexico rust population had recently experienced a demographic bottleneck and was more similar to populations in California than other potential sources. Van Arsdel et al. (1998) reported the oldest known infections on pine in the Sacramento Mountains and dated introduction to circa 1970. Even though the rust was widely distributed in the Sacramento Mountains by 1985 (Geils 2000; Conklin 2004), information on its source, dispersal pathway and establishment would be useful for understanding the potential for other, disjunct outbreaks (Geils et al. 2003) and gene flow (Hamelin et al. 2000), and the impacts of climate change on rust ecology and evolution.
This study applied an upper level synoptic classification (ULSC) (Frank et al. 2008, this issue) to identify upper-level flow patterns likely to transport WPBR spores from Sierra and Cascade source regions to target regions in the Great Basin, the southwestern United States and southern Rocky Mountains and to determine when these patterns occur. The resulting upper-level likelihood calendar was coupled with surface data to determine when, and how often, conditions for transport are followed by conditions favorable for infection, the combination of which is required for establishment. Initially, the methodology was applied and tested with a single target site in the Sacramento Mountains, New Mexico. Subsequently, to validate the model, the methodology was applied to five additional sites in the southwestern United States with known WPBR infestation status. Finally, the susceptibility of nine more points was explored to provide a more complete view of the potential vulnerability of white pine populations in the region.
Methods
Criteria derived from information on the distribution and epidemiology of WPBR were applied to meteorological observations and used to group and rank periods for likelihood of LDD. Information on WPBR described the potential source and season of spore production, longevity, and temperature and humidity requirements for germination and infection (Van Arsdel et al. 2006). Patterns characterizing upper-level air flow were generated by the ULSC (Frank et al. 2008, this issue). Corresponding surface conditions were extracted from the NCEP/NCAR Reanalysis Project (Kalnay et al. 1996) for the April to July season of spore release in the years 1965 to 1974. Results of analyses of these data for likelihood of upper-level transport and of surface conditions favorable for infection were combined to produce a calendar of establishment likelihood and summary tables.
Upper-level likelihood scenarios
The ULSC considers four meteorological variables: geopotential height, specific humidity, and u- and v-wind components, at the 500 hPa level over North America from the NCEP/NCAR Reanalysis Project data set (Kalnay et al. 1996; NOAA-CIRES 2002). This data set is generated from the application of mathematical data assimilation and forecasting models to historical weather data from numerous sources (UCAR 2003). The result is a global, gridded (2.5°), four-times-daily data set with a period of record beginning 1 January 1948. The 500 hPa level is selected for this analysis because it is generally midway between the levels of divergence and non-divergence; this level is commonly thought of as representative of the steering circulation for surface systems. These data, for the period 1965–1974, were subjected to principal component analysis to standardize and reduce the data set, and an average linkage clustering algorithm then identified groups of observations with similar flow patterns. The procedure yielded 16 clusters. The flow patterns identified by the ULSC typify all patterns expected to be observed over the study area, and the resulting cluster calendar showed that the clusters are generally temporally continuous. Subsequent classification of additional observations through a z-score method produced acceptable results, indicating that additional observations may easily be incorporated into the ULSC calendar by this means. Data analysis that identified patterns of similar upper-level conditions and how the patterns were best fit to each observation are described further by Frank et al. (2008, this issue).
Using the ULSC, four-times-daily observations of upper-level flow patterns for the April to July season in the years 1965 to 1974 were ranked by their likelihood of air flow passing directly from a source region in the Sierra-Cascade ranges to the target location in the Sacramento Mountains. Additional ULSC patterns for the April to July season in the years 1975 to 1990 were considered for likelihood of air flow passing from source regions in the Sierra, Cascade and Bitterroot Ranges to the Sacramento Mountains and 14 additional targets in the area (Fig. 1). In this analysis, upper-level-flow patterns were classified and ranked on a 1–4 likelihood scale for flow leading from source regions to a target site using knowledge-based visual interpretation. Each ULSC pattern was classified from low to very high for likelihood of potential upper-level transport and assigned a corresponding ranking from 1 to 4. To represent the persistence of transport favorability, each observation was then assigned the value of the average of the three four-times-daily observations centered on the observation. Since these sums were divided by 3, the average could artificially inflate the score of an observation with an unlikely upper-level pattern in transition between flows with higher likelihoods from different sources. These transition observations were identified and the artificially inflated scores reduced.

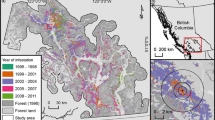
Fifteen points in the southwestern United States for which likelihood of white pine blister rust (WPBR) infestation was calculated are shown (triangles) over a map indicating WPBR infestation status. Data compiled by B.W. Geils, 19 June 2006
Surface likelihood scenarios
From the NCEP/NCAR Reanalysis Project, four-times-daily observations of surface temperature and relative humidity data were obtained for the spore season (April–July) in the years 1965 to 1974 at the grid point (32.5°N, 105°W; Fig. 1, Sierra Blanca) representative of the initial infection site identified in the Sacramento Mountains and which included the approximated time of initial infection (Van Arsdel et al. 1998). Correspondence between Reanalysis data at the surface (NOAA-CIRES 2002) and records from the Cloudcroft, New Mexico, COOP weather station (NESCIS 2003) confirmed that the Reanalysis data were representative of the actual surface conditions at areas where rust had established and intensified. Due to the remoteness of the target locations, meteorological data are generally not available. Reanalysis data were selected for use over other data sets because they consider historical weather data from numerous sources and are available on a fine temporal resolution. The data were obtained for dispersal seasons in the years 1975 to 1990 to expand the investigation to a longer period that might encompass potential establishment at all 15 target sites identified in Fig. 1.
Epidemiological criteria of time, temperature and humidity for germination and infection were applied to the surface data. WPBR spores deposited on ribes leaves might survive a maximum of 22 days, and spores might germinate and infect if maintained for 6–24 h in saturated air above 13°C (Van Arsdel et al. 2006). Because leaf-wetness instruments have serious limitations and other coarse-resolution weather models have provided acceptable estimates of wetness information at the micro-scale (Seem et al. 2000), a hypothesis that synoptic data could be used to assess micro-scale surface conditions was provisionally accepted. A relative humidity of 90% at the observation point was considered sufficient to indicate a saturated condition (necessary for germination) at the level of the ribes leaf.
Periods that satisfied meteorological criteria for the Sacramento site ranged in duration from a single observation (there were many) to 66 h (only one period of this duration was observed during the 1965–1974 study period). It was assumed that longer periods of favorable surface conditions are more conducive to spore germination/infection. Therefore, each observation falling within a favorable period of one observation or more was given a value equal to the reciprocal of a value calculated by the third-degree polynomial fit to the plot of the frequency of extended periods of favorable surface conditions. That is, there were 64 single-observation periods but only nine 18-h-long periods of favorable surface conditions during the study period, so the weight for a favorable period of 18 h in duration was approximately 1/9 while the weight for a period of 6 h in duration was only about 1/64. A third-degree polynomial was chosen over a linear or second-degree polynomial because it would more closely fit the generally log-normal distribution of the frequency plot.
Observations occurring soon after deposition were assumed to be more likely to result in infection than later observations, and no infection was considered possible 22 days or more after deposition, so each observation was assigned a value equal to the sum of the scores of the 21 subsequent days’ observations, weighting each observation’s score with a linearly decreasing multiplier, ranging from 1 for the immediate subsequent observation to 0 for the observation 22 days later. Finally, these weighted values were classified into four ranks similar to those of the upper-level values: (4) greater than 26.4 (four standard deviations): very high; (3) 26.4 to 13.2: high; (2) greater than 0 but less than 13.2: moderate; (1) 0: low.
Coupling of upper-level and surface likelihood scenarios
Scores for upper-level transport and surface infection were summed for each observation and classified to identify the relative likelihood of establishment for each observation during the examined dispersal seasons. Threshold values of 6, 5 and 4 were selected to place the summed transport and infection scores into four nominal classes for likelihood of establishment (the combination of transport and infection). These classes were very high for a score >6, high for a score 5.1–6, moderate for a score 4.1–5, and low for a score ≤4. The stipulation that a score be greater than 6 rather than equal to 6 was chosen to avoid classifying an observation as very high when either the upper level or surface term was less than high.
A sensitivity test was undertaken to determine if these thresholds were appropriate. Increasing the thresholds, from 5.0, 4.0 and 3.0, respectively, did not affect the number of observations placed in the ‘high’ and ‘very high’ classes but increasing the threshold by only 0.1 moved nearly 17% of the days from the ‘moderate’ to the ‘low’ class. Additional increase of the thresholds to 0.3 above the baseline had no effect on the observations placed in any of the classes. The frequency distribution for the 0.1 elevated thresholds was deemed to be the most robust and these thresholds were used for the final classification.
Indices for likelihood of conditions favorable for transport, infection, and establishment were computed for all 15 sites from observation frequencies for upper-level, surface, and coupled conditions, respectively, and grouped by favorability classes, very high, high, moderate and low (data for six sites are presented in Tables 2 and 3). This resulted in 12 frequencies for each point. A weighting scheme and summation was employed to combine the favorability class information and result in only three indices at each site: one for upper-level transport, one for surface conditions and one for likelihood of germination. In the weighting process, the frequency for a moderate class was multiplied by 1/7, high by 2/7, and very high by 4/7 (i.e., 1/7 + 2/7 + 4/7 = 7/7, preserving the relative frequency of moderate + high + very high observations to low observations while factoring in differences among moderate, high, and very high classes). The weighted class-frequencies were then added together to obtain, for each site, one index representing transport, another index for infection, and third for establishment (Table 5). Finally, for graphical displays that illustrate regional trends, indices were krieged and plotted as continuous gradient maps (Figs. 2, 3, 4).

Likelihood of upper-level transport of WPBR spores to the study area during the April–July spore season in the years 1975–1990

Likelihood of surface conditions favorable for infection by WPBR spores in the study area during the April–July spore season in the years 1975–1990

Likelihood of establishment of WPBR, the combination of spore transport at upper levels of the atmosphere and coincident surface conditions favorable for infection, in the study area during the April–July spore season in the years 1975–1990
Results
Assessing the model and likelihood of LDD
The synoptic model for likelihood of LDD of WPBR was developed and tested in three phases over several years. First, the procedure was implemented and evaluated for the only infestation in the region known at the time—the Sacramento Mountains. Model performance was judged by identification and ranking of likely episodes for establishment compared to the previously published age of oldest infections. Next, five additional targets were added: two sites that were subsequently discovered to be infested and three sites where searches as of 2007 have still failed to find established infestations. Model performance was judged by its discrimination between sites differing in presence/absence of pine infection. Finally, another nine sites were included to complete a regular grid across a region of interest. Very recent discoveries of young, spot infestations provided the opportunity to further assess model predictions against field observations using a case-history approach.
Sacramento mountains 1965–1974
The ULSC used cluster analysis to identify 16 patterns of similar upper level flow, named for dominant season and barometric structure (Frank et al. 2008, this issue). Only four of these clusters, however, are rated favorable for LDD of WPBR to the Sacramento Mountains (see Frank et al. 2008, this issue, Figs. 2 and 4). A cluster typified as a summer-trough (ST) pattern provides conditions with a very high likelihood for transport. A trough in the western United States generates a zone of higher wind speeds passing over the source region with a strong, direct flow to the target. This pattern occurs on 14.8% of the observations in the April–July spore season. Two other clusters, the winter-trough-ridge (WTR) and the summer-trough-ridge (STR), are rated as high for transport likelihood. The WTR pattern is also characterized by a trough in the western United States, but flow is weaker and traverses farther south in the transit from the source to the target. The WTR is observed 2.2% of the time during the spore season. Although the STR flow over the southwestern United States is more zonal than for ST or WTR, and the area of stronger winds is further north, transport likelihood is still rated as high. This pattern occurs on 19.5% of the observations during the spore season. The observations with a winter-zonal-warm (WZW) pattern are rated as only moderate for transport. This pattern occurs 7.9% of the time during the spore season and is distinguished by nearly direct upper-level flow from the west to east, but displays weaker upper-level winds that reduce the likelihood of timely source-to-target transport. All other ULSC patterns are rated low for transport likelihood because of either very weak flow or poor alignment.
Classification of four-times-daily meteorological observations for WPBR transport and infection indicates that unfavorable conditions occurred much more frequently than favorable conditions. For transport only, observations rated as very high were relatively frequent, accounting for 13% of those in the 1965–1974 study period. Coupling with surface infection requirements, however, greatly reduced the frequency of favorable conditions for rust establishment (transport and infection): 80% low, 13% moderate, 6% high, and only 0.7% very high. The observations with very high ratings occurred most frequently (>60%) in the later half of the dispersal season (June and July). The 33 observations rated as very high resulted from five different combinations of upper level and surface conditions, and occurred as five episodes of variable length in only four of the years in the study period (Table 1). There were brief episodes in June 1972 and July 1971, a longer episode beginning the end of June 1968, and a 2-week period in June 1969 when conditions remained very high for multiple periods exceeding 24 h.
The synoptic pattern for June 1969 (Table 1) began with a trough-ridge flow and moderate-to-high likelihood for transport; on 4 June the first of several transitions occurred. The following period of 5–7 June was dominated by an ST pattern with very high likelihood for transport. After a few days (8–10 June) when a period of low likelihood for transport prevailed, a short period (11 June) of ST developed with a high likelihood for transport. For the next few days (to 15 June), an STR persisted and maintained a high likelihood for transport. The patterns, duration, and sequences of synoptic conditions for June 1969 had also occurred in nearly every year examined. The distinctive feature of June 1969 was the coincidence of an extended period favorable for transport with development of unusually persistent surface meteorological conditions (66 h beginning at 0600 hours on 14 June) that provided nearly continuous favorable conditions for infection. The weather in the Sacramento Mountains for the following months of July, August and September was typical of the region and adequately cool and wet for the rust to infect white pines and thereby establish a persistent infestation, with infection alternating between ribes and pines.
Although canker aging can only approximate time since infection, it does provide an independent observation of infestation history (Kearns 2005). Considering the uncertainty of finding and correctly dating the oldest canker, the modal estimate of Van Arsdel et al. (1998) for the infestation origin of 1970 implies “sometime from 1965 to 1974.” If the dispersal model fits epidemiology and meteorology well, any of the observations rated very high for establishment could be the actual origin. Considering, however, the continuity of the June 1969 observations, this is most likely the origin of the Sacramento infestation. This demonstrates the utility of the modeling approach for identifying potential dates for origins of infestations.
Six target sites 1975–1990
Five more sites in the southwestern United States were identified for further evaluation of the model (Table 2). This provided three targets with pine populations known to be infested: the original target in the Sacramento Mountains (Sierra Blanca), a population in the Sangre de Cristo and Wet Mountains in southern Colorado (La Veta), and a population in the Jarbidge Mountains, Nevada (Elko), and three targets where intensive searches have failed to discover WPBR as of 2007: the plateaus of southwestern Utah (Bryce Canyon), the San Francisco Peaks, Arizona (Flagstaff), and the Spring and Sheep Creek Mountains, Nevada (Needles). Upper-level patterns were examined for likelihood of transport to the new points, and surface data were acquired for the period 1975–1990, inclusive of the time when the rust is believed to have first reached the La Veta and Elko target points.
The three infested sites had high or very high modeled likelihood of establishment for the period considered, whereas sites not known to be infested were rated to have only low or moderate likelihood (Table 2). A detailed examination of the calendar for the Elko site reveals nine consecutive observations beginning 2 July 1979 00Z with highly favorable establishment likelihood. The correspondence of this date with an independent estimate of the infestation origin by canker age, 1975–1984 (Vogler and Charlet 2004), suggests that coupled conditions rated as high (not just very high ratings) may be sufficient for successful rust dispersal. Although the possibility of establishment resulting from episodes rated moderate cannot be excluded, the infrequency of observations of even moderate rating at putatively non-infested sites demonstrates that the model distinguishes among sites differentiated by infestation status.
Examination of the surface and upper-level rankings before the coupling of the two scores shows that, separately, neither surface nor upper-level flow conditions are limiting factors for rust establishment (Table 3). Infested and un-infested points show similar frequencies of occurrence of surface and upper-level conditions. For example, the un-infested Bryce Canyon site has higher frequencies of very high likelihoods for both surface and upper-level flow than does the infested La Veta site. However, La Veta experienced 41 observations of high establishment likelihood during the study period while Bryce Canyon had none (Table 2). This suggests that the coincidence of favorable upper-level flow patterns with surface conditions conducive for germination is the unique condition that permits establishment of a population by LDD.
Nine additional sites 1975–1990
To further examine the susceptibility of white pine populations in the southwestern United States, nine additional target points were selected from the Reanalysis data set, bringing the total number of target sites to 15. These points were: northwestern Utah (Dugway), east-central Nevada (Caliente), southeastern Utah (Blanding), southwestern Colorado (Durango), northeastern Arizona (Winslow), northwestern New Mexico (Grants), northeastern New Mexico (Cline’s Corners), southeastern Arizona (Stafford) and southwestern New Mexico (Deming). Four of these sites were eventually recognized as infested (Table 4). Infected ribes were found in 2005 on the West Tavaputes Plateau in southern Duchesne County, Utah, (east of Dugway) but infected white pines have not been discovered (B.W. Geils, unpublished data). A small population of infected white pine was found in the Jemez Mountains of Sandoval County, New Mexico, (Grants) and dated to 1999 (D. Conklin, personal communication). In 1999, an infestation was discovered by Dave Conklin on Gallinas Peak, Lincoln County, New Mexico, (Cline’s Corners); the infestation was then several years old. Another small, localized but well-established population of infected white pines was discovered (also by Dave Conklin) in the New Mexico San Francisco Mountains of western Catron County (nominally Safford). The infestation was dated by canker age observations in 2007 to circa 1995.
The four infestations at the added sites provided additional data for testing minimum coupled conditions sufficient for establishment (Table 5). Although infrequent (0.001), very highly favorable observations had occurred at Cline’s Corners of the Gallinas Peak infestation. Highly favorable conditions were observed at Grants (0.007) and Safford (0.009) where the Jemez Mountains and San Francisco Mountains infestations were discovered. Within the period of study, 1975–1990, no very highly or highly favorable observation were recorded for Dugway where rust was found on ribes in 2005 (Tavaputes Plateau infestation). Moderately favorable observations had been recorded there, however, with a frequency of 0.292.
Synoptic climatology for LDD of blister rust in the greater southwest
In the greater Southwest, upper-level conditions favorable for rust transport occur more frequently than surface conditions favorable for establishment (Table 5). The least favorable site for transport is Deming and the most favorable is Elko (index values of 0.035 and 0.380, respectively). Infestations occur near both of these sites, suggesting that upper-level transport is not a limiting factor. The regional pattern of upper-level transport likelihood is shown in Fig. 2. Upper-level transport likelihood displays a peak in the northwest corner, high values in the extreme north and west, and lowest values at Deming and Flagstaff.
The least favorable site for infection is Durango, and the most favorable is Sierra Blanca (index values <0.001 and 0.076, respectively). The infestation known to be nearest to Durango is about 2.5° east (in the Sangre de Cristo Mountains, Table 4); the oldest and most extensive infestation of the region is in the Sacramento Mountains and adjacent ranges of the Sierra Blanca site. Figure 3 shows the regional pattern of favorability for surface conditions. This pattern displays a peak in the southeast corner, high values extending across the central New Mexico ranges into the ranges of south-central Colorado, and lowest values in the northwest corner.
Establishment of the rust requires the timely sequence of transport and infection modeled as a coupled index (Table 5, Fig. 4). The least favorable site is Flagstaff, and most favorable site is Elko (index values <0.001 and 0.066, respectively). An infestation is well established in the Jarbidge Mountains north of Elko (Table 4); no infestation has yet been found in the forests around Flagstaff. The coupled likelihood pattern has a peak in the northwest corner with high values extending across the western and northern portions (reflecting the influence of the upper level factor) and a second, lower peak in the southeastern corner (reflecting influence of the surface factor). Less favorable areas are central and southeastern Arizona (reflecting low coupling where the two factors are individually moderate). Overall, the map of the coupled, synoptic index accounts well for known infestations; the poorest fits occur where infestations appear restricted to local micro-sites (e.g., the New Mexico San Francisco Mountains). The model identifies vulnerable areas in eastern and southern Nevada and across Utah as favorable for spring transport and ribes infection by WPBR.
Discussion
Dispersal to the Sacramento mountains
From observations of heavy infection by a native rust on ribes and knowledge of the climate at Cloudcroft, New Mexico, an early report from the Bureau of Plant Industry (1921) predicted that “if blister rust ever reached this region [Sacramento Mountains], it would flourish...[there].” The rust did reach the Sacramento Mountains and it now seriously infests the adjacent White Mountains (Sierra Blanca) and nearby Capitan Mountains (Geils 2000).
Several lines of evidence support the hypothesis that the initial Sacramento infestation resulted from a single introduction in 1969 by aerial transport from California. The advanced age and remote location of cankers suggests importation and planting of infested pines or ribes is unlikely (known transplanting of seedlings from Idaho occurred after 1970). The genetic profile of the rust (Hamelin et al. 2000) is explained by a single introduction of a rust from California. By 1969, WPBR had been in California for several decades and had spread well south along the Sierra Nevada (Kliejunas 1985; Kliejunas 2002). Although a flight from California is more than 1,200 km, the rust also appears to have completed other, somewhat shorter dispersals (Geils et al. 2003), and longer distances for other rusts are reported (Nagarajan and Singh 1990). Source regions such as Oregon, Idaho and Wyoming cannot be ruled out; few, infrequent synoptic patterns (Frank et al. 2008, this issue) would transport rusts from these locations to south-central New Mexico.
In addition to transport of viable rust spores, deposition must occur on a susceptible telial host, followed by infection, development over the season by several other spore stages, infection of white pine and development of perennial cankers. The Sacramento Mountains are noteworthy in the Southwest United States for a large and extensive population of Ribes pinetorum, a susceptible ribes capable of supporting a large inoculum potential (Van Arsdel and Geils 2004). These mountains also have, in close proximity to the ribes, many very susceptible southwestern white pine (Pinus flexilis var. reflexa). Other mountainous areas of the region also have some ribes populations, white pine populations, exposure to western and northern rust sources and spring-summer moisture; the intensities of these factors vary geographically (e.g., few ribes in some ranges well populated with white pines).
Upper-level flow and spore transport
The patterns of upper level flow identified by the ULSC (Frank et al. 2008, this issue) describe the potential connections between source and target locations. Although host and rust populations have patchy distributions at various landscape scales, they can be mapped at a county level using data from published literature, herbarium records, and field reports (Fig. 1, review completed by B.W. Geils, June 2006). County distributions are sufficient to identify source and target areas for a synoptic analysis with a spatial resolution of a 2.5° grid. Given their proximity to western and northwestern sources, there is little surprise that sites in Nevada and Utah are frequently exposed to favorable conditions for spore transport (Fig. 2). The relatively frequent occurrence of flow across the entire region suggests transport alone may not be limiting, at least as far away from the source populations as central Colorado and eastern New Mexico. This agrees with the finding of Aylor (2003) that LDD may be more often limited by establishment at the deposition site than by conditions of transport.
Surface conditions and infection of ribes
In general, deposition of rust spores and the following stages in the rust life cycle are processes determined by a microenvironment of temperature and moisture that usually varies over very short distances (leaf boundary layer or in sun vs in shade). However, in this case we are interested in relatively long periods of very high humidity in a regionally dry climate zone. When synoptic data suggest conditions are favorable for infection, the conditions ought to occur widely across the area. Additional episodes favorable for infection are likely only in special habitats such as narrow, wet canyons. Although there is a low probability that a rust spore released at 1,000 km distance will arrive at a site, probability of infection is even lower if there are fewer ribes leaves. The synoptic model does not explicitly include assessment of the abundance of ribes, which varies greatly from location to location within a synoptic site (see Kearns 2005). Conditions favorable for infection are generally uncommon in the region, but there is significant geographic variation and micro-site exceptions exist.
Coupled conditions and initial establishment of blister rust
The time limitations on viability of rust spores require that, for a spore to establish an initial infestation on ribes, conditions suitable for germination and infection must follow soon after deposition. Coupling upper-level and surface conditions into a single metric identifies and ranks the favorability of this sequence of events. Comparing the relative frequency of these events across a region then provides a means of assessing the risk of LLD at a synoptic scale. There remains, however, uncertainty over the minimum conditions necessary for establishment. Two results support an assertion that a sequence of highly or very-highly rated observations satisfy these minimum conditions. First, in a comparison of infested and putatively non-infested sites, ratings of high or better occurred only at the infested sites (Table 2). Second, for the Sacramento and Jarbidge Mountains infestations, the coupled calendars’ high and very high observations agree with the estimates of origin year by canker age. Very highly and highly favorable observations are consistently recorded for the infested sites Elko, La Veta, Grants, Cline’s Corners, Safford, and Sierra Blanca, but these events are rare (<0.02). On the West Tavaputes Plateau in 2005, B.W. Geils found infected ribes in a wet cove of the Dugway site. This site had not previously (1975–1990) experienced periods identified as highly or very highly favorable for establishment, but moderate observations had been frequent (0.292). Although individual moderately favorable conditions may not identify specific times when establishment could occur, many moderately favorable periods could be an indication that suitable conditions are sometimes met in the best local habitats.
A coupled index based on the weighted frequencies of linked upper level and surface conditions provides a reference for assessing the LDD risk of blister rust. The coupled index and its component indices help to understand the transport and infection processes that result in either the introduction of the rust to a new area or the long distance movement of genes from a donor to a recipient population. A better understanding of the production of spores and their escape from the canopy would help to refine the model. Other factors, such as the abundance and proximity of susceptible hosts and the frequency of favorable meteorological conditions for continuing reproduction of the rust, are also important for determining the spread and intensification of an infestation at the stand and landscape levels.
Applications
The synoptic climatology model for LDD simplifies many biological requirements for successful infestation by WPBR but still provides a useful display of vulnerability at a regional scale. Because the Arizona San Francisco Peaks have susceptible populations of white pine and ribes in close proximity to each other (and to motivated forest pathologists), this area will receive continued monitoring for early detection. The synoptic model, however, suggests that a lack of rust in the Peaks is not sufficient reason to forgo continued reconnaissance in the “drier” Great Basin and “more distant” Southern Rockies, where sites are more, not less, vulnerable to infestation. Susceptible populations of white pines and ribes also occur in Mexico, but their vulnerability to WPBR is unknown. The synoptic model provides a tool for assessing the potential of LDD introducing blister rust in to Mexico. Over time, global climate changes and, consequently, the genetic connections between metapopulations are altered. The synoptic model provides a framework at a sufficiently large and long scale to develop a means to assess an important component of forest health in response to climate change.
Afterword
Since this paper was submitted, scouting for additional infestations in the southwestern United States has continued. By the end of 2007, several additional, very small, infested areas had been discovered, but some areas, such as Flagstaff, appear to remain uninfested. D. Conklin and B.W. Geils (unpublished data) located more trees, infected for a long period in the San Francisco Mountain infestation as well as other, small and recent infestations about 40 km and 100 km to the southeast. D. Conklin has also located several additional, infested locations near to the original Jemez Mountain discovery. These may indicate that secondary spread has already occurred. R.S. Danchock photographed a single canker in the Zuni Mountains about 30 km southwest of Grants, New Mexico; we have not yet determined if this single canker is part of an enduring infestation. The apparent lack of perennial cankers at West Tavaputes Plateau, Utah, suggests that this is another case whereby infection of ribes was not followed by infection of pine. None of these observations, however, refute the general conclusions of the synoptic model—that within this Southwestern, synoptic region, transport of rust spores is nowhere limiting but, where conditions for infection are suitable, varies from widespread in the southeast to eastern portions to only localized elsewhere.
References
Andow DA, Kareiva PM, Levin SA, Okubo A (1990) Spread of invading organisms. Landscape Ecol 4:177–188
Aylor DE (1986) A framework for examining inter-regional aerial transport of fungal spores. Agric For Meteorol 38:263–288
Aylor DE (2003) Spread of plant disease on a continental scale: role of aerial dispersal of pathogens. Ecology 84(8):1989–1997
Blodgett JT, Sullivan KF (2004) First report of white pine blister rust on Rocky Mountain bristlecone pine. Plant Dis 88:311
Bowden J, Gregory PH, Johnson CG (1971) Possible wind transport of coffee leaf rust across the Atlantic Ocean. Nature 229:500–501
Brown JKM, Hovmoller MS (2002) Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 297(5581):537–541
Bureau of Plant Industry (1921) Annual report for 1921 on the white pine blister rust work in the Far West. Unnumbered report. U.S. Department of Agriculture, Berkeley, CA, p 25
Caswell H, Lensink R, Neubert MG (2003) Demography and dispersal: life table response experiments for invasion speed. Ecology 84(8):1968–1978
Clark JS, Lewis M, McLachlan JS, HilleRisLambers J (2003) Estimating population spread: what can we forecast and how well? Ecology 84(8):1979–1988
Conklin DA (2004) Development of the white pine blister rust outbreak in New Mexico. Rep. R3-04-01. Albuquerque, NM: U.S. Department of Agriculture, Forest Service, Southwest Region. 11 p. Online:http://www.fs.fed.us/r3/publications/documents/wp_blister_rust_nm.pdf. [Accessed October 17, 2006]
Davis JM, Monahan JF (1991) Climatology of air parcel trajectories related to the atmospheric transport of Peronospora tabacina. Plant Dis 75:706–711
Draper MA, Walla JA (1993) First report of Cronartium ribicola in North Dakota. Plant Dis 77:952
Frank KL, Geils BW, Kalkstein LS, Thistle HW Jr (2008) Synoptic climatology of the long-distance dispersal of white pine blister rust. I. Development of an upper level synoptic classification. Int J Biometorol (this issue)
Fujioka FM (1996) Climatic factors that influence the spread of white pine blister rust. In: Kinloch BB Jr, Marosy M, Huddleston ME (eds) Sugar pine: status, values and roles in ecosystems, proceeding of a symposium presented to the California Sugar Pine Management Committee. Pub 3362. University of California, Division of Agriculture and Natural Resources, Davis, CA, pp 119–124
Geils BW (2000) Establishment of white pine blister rust in New Mexico. HortTechnology 10(3):528–529
Geils BW (2001) Impacts of white pine blister rust. In: Fosbroke SLC, Gottschalk KW (eds) Proceedings, U.S. Department of Agriculture interagency research forum on gypsy moth and other invasive species 2001; 2001 January 16–19; Annapolis MD. Gen Tech Rep NE-285. U.S. Department of Agriculture, Forest Service, Northeastern Research Station, Newton Square, Pennsylvania
Geils BW, Conklin D, Frank K, Guyon J, Harris JL, Hoffman J, Jacobi W, Kearns H, Newcomb M, Smith E, Van Arsdel E, Vogler D (2003) New information on the distribution of white pine blister rust for 2002. In: Stone J, Maffei H (eds). Proceedings of the 50th Western International Forest Disease Work Conference; 2002 October 7–11; Powell River, BC. U.S. Department of Agriculture, Forest Service, Central Oregon Service Center, Bend, OR
Hamelin RC, Hunt RS, Geils BW, Jensen GD, Jacobi V, Lecours N (2000) Barrier to gene flow between eastern and western populations of Cronartium ribicola in North America. Phytopathology 90:1073–1078
Hawksworth FG (1990) White pine blister rust in southern New Mexico. Plant Dis 74:938
Hunt RS (2003) White pine blister rust. Recent Res Dev Mycol 1(2003):73–85
Isard SA, Gage SH, Comtois P, Russo JM (2005) Principles of the atmospheric pathway for invasive species applied to soybean rust. BioScience 15(10):851–861
Kalnay E, Kanamitsu M, Kistler R, Collins W, Deaven D, Gandin L, Iredell M, Saha S, White G, Woollen J, Zhu Y, Chelliah M, Ebisuzaki W, Higgins W, Janowiak J, Mo KC, Ropelewski C, Wang J, Leetmaa A, Reynolds R, Jenne R, Joseph D (1996) The NCEP/NCAR reanalysis 40-year project. Bull Am Meteorol Soc 77:437–471
Kearns HSJ (2005) White pine blister rust in the central Rocky Mountains: Modeling current status and potential impacts. Dissertation. Colorado State University, Fort Collins, Colorado
Kliejunas J (1985) Spread and intensification of white pine blister rust in the southern Sierra Nevada. Phytopathology 75:1367
Kliejunas J (2002) An update on the southern extent of white pine blister rust in California. Rep R02-01. U.S. Department of Agriculture, Forest Service, Pacific Southwest Region, State and Private Forestry, Forest Health Protection, Vallejo, CA
Lundquist JE, Geils BW, Johnson DW (1992) White pine blister rust on limber pine in South Dakota. Plant Dis 76:538
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz F (2000) Biological invasions: causes, epidemiology, global consequences and control. Issues in Ecology 5, Ecological Society of America, Washington, D.C.
Mielke JL (1943) White pine blister rust in western North America. Bulletin 52. Yale University, School of Forestry, New Haven, CT
Mims SA, Mims FM (2004) Fungal spores are transported long distances in smoke from biomass fires. Atmos Environ 38(5):651–655 DOI 10.1016/j.atmosenv.2003.10.043
Munoz J, Felicisimo AM, Cabezas F, Burgaz AR, Martinez I (2004) Wind as a long-distance dispersal vehicle in the Southern Hemisphere. Science 304:1144–1147
Muller-Landau HC, Levin SA, Keymer JE (2003) Theoretical perspectives on evolution of long-distance dispersal and the examples of specialized pests. Ecology 84(8):157–167
Nagarajan S, Singh DV (1990) Long-distance dispersal of rust pathogens. Annu Rev Phytopathol 28:139–153
NESCIS (2003) TD-3200 Cooperative summary of the day. U.S. Department of Commerce, Washington, D.C.
NOAA-CIRES Climate Diagnosis Center (2002) http://www.cdc.noaa.gov/cdc/data.ncep.reanalysis.html. [Accessed 12 March 2002]
Purdy LH, Krupa SV, Dean JL (1985) Introduction of sugarcane rust into the Americas and its spread to Florida. Plant Dis 69:689–693
Samman S, Schwandt JW, Wilson JL (2003) Managing for healthy white pine ecosystems in the United States to reduce the impacts of white pine blister rust. Rep R1-03-118. U.S. Department of Agriculture, Forest Service, Missoula, Montana
Seem RC, Magarey RD, Zack JW, Russo JM (2000) Estimating disease risk at the whole plant level with General Circulation Models. Environ Pollut 108(3):389–395
Smith JP, Hoffman JT (2000) Status of white pine blister rust in the Intermountain West. West N Am Nat 60(2):165–179
UCAR (2003) NCEP/NCAR Reanalysis > Project Description. http://dss.ucar.edu/pub/reanalysis/rean_proj_des.html. [accessed August 4, 2003], April 1
Van Arsdel EP (1967) The nocturnal diffusion and transport of spores. Phytopathology 57:1221–1229
Van Arsdel EP (1973) Origin of south Texas peanut rust epidemics. In: Calpouzos L, Campos A (coords) Symposium on the aerobiology of diseases, pests, and allergens in the Western hemisphere. Science and Man in the Americas. June 20–July 4, 1973
Van Arsdel EP, Geils BW (2004) The Ribes of Colorado and New Mexico and their rust fungi. Rep. FHTET 04-13. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team.
Van Arsdel EP, Riker AJ, Kouba TF, Suomi VE, Bryson RA (1961) The climatic distribution of blister rust on white pine in Wisconsin. Station Paper 87. U.S. Department of Agriculture, Forest Service, Lake States Experiment Station, St. Paul, MN
Van Arsdel EP, Conklin DA, Popp JB, Geils BW (1998) The distribution of white pine blister rust in the Sacramento Mountains of New Mexico. In: Jalkanen R, Crane PE, Walla JA, Aalto T (eds) Proceedings of the 1st IUFRO rusts of forest trees working party conference; 1998 August 2–7; Saariselkä, Finland. Research Paper 712. Finnish Forest Research Institute, Rovaniemi, Finland
Van Arsdel EP, Geils BW, Zambino PJ (2006) Epidemiology for hazard rating of white pine blister rust. In: Guyon, J (comp) Proceedings of the 53rd Western International Forest Disease Work Conference; 2005 August 26–29; Jackson, Wyoming. U.S. Department of Agriculture, Forest Service, Intermountain Region, Ogden, UT
Vogler DR, Charlet DA (2004) First report of the white pine blister rust fungus (Cronartium ribicola) infecting whitebark pine (Pinus albicaulis) and Ribes spp. in the Jarbidge Mountains of northeastern Nevada. Plant Dis 88:772
Waggoner PE, Aylor DE (2000) Epidemiology: a science of patterns. Annu Rev Phytopathol 28:71–94
Yonow T, Kriticos DJ, Medd RW (2004) The potential geographic range of Pyrenophora semeniperda. Phytopathology 94:805–812
Acknowledgments
The authors thank Frank Hawksworth for finding the rust; Gene Van Arsdel for insisting it came from California; Dave Conklin and John Popp for showing us where it was; Mary Lou Fairweather for challenging us to explain why not the San Francisco Peaks; Detlev Vogler for encouraging us to see it as a dynamic, interacting, and evolving member of a complex pathosystem; Paul Zambino for comments on the manuscript; and Richard Sniezko for sharing Danchock’s find. This work was funded by the United States Department of Agriculture, Forest Service, Forest Health Protection, Forest Health Technology Enterprise Team Project# TD.01.M01 and Rocky Mountain Research Station (03-JV-11221605-297), and University of Delaware, Department of Geography, Center for Climatic Research. Development of the ULSC and its application to the Sacramento outbreak was performed by Katrina Frank in partial fulfillment of a Ph.D. in Climatology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Frank, K.L., Geils, B.W., Kalkstein, L.S. et al. Synoptic climatology of the long-distance dispersal of white pine blister rust II. Combination of surface and upper-level conditions. Int J Biometeorol 52, 653–666 (2008). https://doi.org/10.1007/s00484-008-0158-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-008-0158-3