
Abstract
Key message
Canopy and stem phenology of Qinghai spruce, central Qilian Mountains, respond to different environmental factors depending on season and elevation.
Abstract
To understand vegetation species response to climate change, much research has been devoted to changes in forest phenology. Results of such studies are not only of scientific interest; they are potentially of great use in forest management. This study focuses on variations in canopy and stem phenology as affected by climate and elevation. We collected data on canopy phenology (as recorded in the Normalized Differential Vegetation Index) and stem phenology [using the Vaganov–Shashkin (V–S) model] in Qinghai spruce (Picea crassifolia) growing at two sites in the central Qilian Mountains, Northeast Tibetan Plateau. One site was at a higher elevation, near the local alpine tree-line, and the other was near the local lower tree-line. At both sites, a significant correlation was found between canopy and stem spring phenology. This would seem to be mainly due to spring temperatures. No such correlation was found between canopy and stem autumn phenology. The study suggests that the main factors affecting stem growth after the beginning of growing season would be temperature and soil moisture, and that these have different effects depending on elevation. At the lower elevation, soil moisture seems to be the main factor limiting growth. At the higher elevation, temperature was the determining factor. Climate change will have different effects depending on elevation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Phenology is attracting much research interest of late. Temporal changes in phenology reflect on-going climate change (Parmesan and Yohe 2003; Peñuelas et al. 2013). Climate change is having, and will have, profound impacts on species ranges, primary productivity, and the vegetation carbon cycle (Settele et al. 2014). It is particularly important to understand the phenology of forests, as climate change has already caused extensive tree mortality and widespread forest die-back in many regions [e.g. (Allen et al. 2010; Carnicer and Mooney 2011; Anderegg et al. 2013; Liu et al. 2013)].
Many studies of forest phenology have focused on canopy phenology. Researchers have noted changes in bud burst, flowering, and leaf defoliation (Chmielewski and Rötzer 2001; Ahl et al. 2006). They have used canopy phenology to assess species ability to adapt to local environmental conditions (Du et al. 2014; Jones et al. 2014) and to predict growing-season changes under global climate change. Canopy phenology studies have been based to a large extent on data from deciduous broadleaf forests (Kross et al. 2011; Schwartz et al. 2002); much less interest has been shown in coniferous forests.
Tree stem phenology research has also gained attention in recent years, as it is central to dendroclimatology and dendroecology studies (Deslauriers et al. 2008; Rossi et al. 2008; Li et al. 2013; Yang et al. 2017). Plant stems constitute good records of apical and radial growth (which depend on the net photosynthetic product of leaves) (Deslauriers et al. 2017); long-lived species can record environmental variation over long time scales. Conifers have been a major source of data for dendroclimatology and dendroecology studies (George 2014).
Some researchers have looked at both stem and canopy phenology (Rossi et al. 2009; Cuny et al. 2012; Antonucci et al. 2015; Perrin et al. 2017). It is important to know how each type of phenology responds to environmental variation. Comparison of these responses can help to understand the relationship between apical and lateral meristems, and thus tree physiology in general. If stem and canopy studies are to be combined, then a good starting point would be conifers, and in particular, the Qinghai spruce (Picea crassifolia). This is one of the conifer species that has been the target of both canopy phenology and dendrochronological studies (Du et al. 2014; Liu et al. 2016).
The Qilian Mountains are located in the northeastern area of the Qinghai-Tibet Plateau. They are home to several long-lived tree species, which have been the subject of a large number of dendrochronology studies (Yang et al. 2014; Zhang et al. 2015; Liang et al. 2016). Qinghai spruce is one of those long-lived species; it is also one of the dominant species in the Qilian forests, accounting for more than half of the total forest area (Cheng et al. 2014; Zhu et al. 2017). These forests have been noticeably affected by global climate change. Starting in the 1960s, the average temperature has risen by 0.26 °C per decade in this region (Du et al. 2014). One would expect changes in Qinghai spruce phenology and physiology, and indeed, those changes have been observed and studied (Chen et al. 2011; Chang et al. 2014; Tian et al. 2017).
The current study builds on previous scientific work in the Qilian region. We have collected both tree-ring width records and canopy phenology data in an effort to elucidate the long-term relationship between the stem and canopy phenology of the Qinghai spruce. It should be noted that there are no long-term, large-scale field observations of bud and stem cambial activity in this area. For the stem phenology data, we used a Vaganov–Shashkin (V–S) model (Anchukaitis et al. 2006; Evans et al. 2006; Vaganov et al. 2006). We relied upon remote sensing data for a long-term record of canopy phenology (Du et al. 2014).
The specific objectives of this study were to detect the temporal relationship between the stem and canopy phenology in virgin Qinghai spruce stands in two elevations in the central Qilian Mountains and to determine how both have been affected by climate change. Phenology reflects the adaption of plants to their environment, hence it is of great importance to learn their response to climate in different elevations. The results are also of great potential assistance to scientific forest management.
Materials and methods
Study area and study species
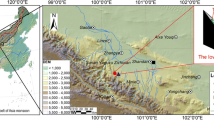
The study was conducted at two sites in the central Qilian Mountains (Fig. 1). One site, QLN, is located close to the Qilian meteorological station (elevation 2787 m). The other site, YNG, is close to the Yeniugou meteorological station (elevation 3320 m). Both the study sites lie in a forested region with a temperate continental climate. Annual rainfall is concentrated in a five-month period, from May to September. Over the period 1960–2014, the mean annual air temperature as measured at the Qilian meteorological station was 0.2 °C. During the same period, the Yeniugou meteorological station showed a mean annual temperature of − 0.9 °C. The maximum and minimum mean daily air temperatures were 22.2 °C max and − 25.5 °C min for Qilian, and 20.6 °C max and − 26.2 °C min for Yeniugou. Mean annual precipitation was 409 mm for Qilian and 419 mm for Yeniugou.

Tree ring sampling sites and meteorological stations
Qinghai spruce is found at elevations ranging from about 2500 to 3400 m a.s.l. The mean height of the mature spruce ranges from 5 to 11 m; the average stand density is about 2400 stems ha−1. The leaf area index (LAI) of canopies averages approximately 2.0 (Du et al. 2014).
Tree-ring chronology and modeling
Following standard dendrochronological methods, tree-ring samples were collected from site QLN (Fig. 1b) in May 2015 and from site YNG in 2007 and 2016. Increment cores (one or two cores) were extracted at breast height from presumably old and healthy trees, which were growing in virgin stands. Trees sampled grew from 2700 to 2900 m at the QLN site (near the local lower tree-line), and from 2900 to 3100 m at the YNG site (near the local alpine tree-line).
The cores were mounted, air-dried, and sanded to make the vessels and rings visible. Ring widths were measured using a Lintab tree-ring measuring system (http://www.rinntech.de) with a precision of 0.001 mm. The quality of the cross-dating was confirmed using COFECHA software (Grissino-Mayer 2001). No missing rings were found. The ring-width chronologies were constructed using RCSsigFree_v45 software (Melvin and Briffa 2008) (http://www.ldeo.columbia.edu/tree-ring-laboratory/resources/software). Age-related growth trends were removed for all samples, using a cubic spline curve. We used an expressed-population signal greater than 0.85 (Wigley et al. 1984; Briffa and Jones 1990) to identify the reliable parts of each ring-width chronology. We collected several descriptive statistics to characterize the chronology: the mean segment lengths of the cores, mean sensitivity, and mean correlation between series.
We used the Vaganov–Shashkin (V–S) model (Anchukaitis et al. 2006; Evans et al. 2006; Vaganov et al. 2006) to establish a model of stem phenology time-series for the two research sites. 1960–2014 data from the Qilian and Yeniugou meteorological stations (records of climate, daily temperature, precipitation) were used as inputs. Even though these stations were not located at the research sites, they were close enough that climate conditions were similar. We attempted to partially correct temperature inputs for the elevation difference between the YNG tree-ring site and the Yeniugou meteorological station with an adiabatic correction of 5.6 °C/km (Chen et al. 2014). This suggested a temperature correction of 1.96 °C. No temperature correction was applied to the data from site QLN; it is quite close to the same elevation as the Qilian meteorological station.
Modeled and actual tree-ring width chronologies were compared. The model was adjusted to attain the highest possible Pearson correlation coefficient. We also used data from a 1-year (2014) dendrometer-detected tree-ring growth study conducted by Tian et al. (2017) at the QLN site. Data for altitudes of 2700 and 2800 m a.s.l. were collected, which served as a useful reference when tuning model parameters. The stem phenology records thus obtained were used to determine seasonal transition dates: start and end of stem growth season (SOSs and EOSs). We could then investigate which major factors were influencing SOSs, EOSs, and the whole stem radial growth season.
Canopy phenology using remote sensing data
We used remote-sensing data (GIMMS and MODIS NDVI) collected from 1982 to 2014 over the Qilian Mountains. The Normalized Differential Vegetation Index (NDVI) is based on the differential reflectance of green vegetation in the infrared and near-infrared bands. It is used as an indicator of plant canopy greenness and productivity, as it has been shown to be strongly correlated with the strength of photosynthetic activity (Pettorelli et al. 2005; Jeong et al. 2011; Zhang et al. 2013). The time-series NDVI data did require some pre-processing before use. Snow coverage during the non-growing season and poor atmospheric conditions (e.g. clouds) add noise to the data and depress NDVI values. After noise elimination, the NDVI time series were fitted (using a double-logistic function based on non-linear least-squares method) to the upper envelope of NDVI data (Jönsson and Eklundh 2004). We extracted the pixels representing the locations of the meteorological stations from the satellite data for further analysis.
We used the inflection-point-based method to determine two transition dates: start of canopy growth season (SOSc) and the end of canopy growth season (EOSc). This method has been used to study a variety of biomes in different climatic zones and has been well-validated by many researchers [see, among others, (Schwartz et al. 2002; Yang et al. 2012; Du et al. 2014)]. It is based on the rate of change of the fitted NDVI curve, that is, the derivative of the curvature. We defined the dates of SOSc or EOSc as the dates on which RC values achieved local maxima on the left (SOSc) or the right (EOSc) edge of the annual curve.
Statistical analysis
We analyzed the relationship between climate and tree stem radial growth for the period 1960–2014 through correlation analysis of ring-width chronology and the climatic data. We used DendroClim2002 software (Biondi and Waikul 2004). The meteorological data used: mean monthly temperature and monthly total precipitation. Statistical significance levels were estimated with two-tailed significance tests.
We used Pearson’s correlation coefficient and multiple linear regression analysis to investigate the influence of climate factors that could be controlling SOSc and EOSc variation. As the V–S model works with three main factors, we investigated the effects of eighteen variables: six warmth-related (mean temperatures 10, 20, 30, 60, 90 and 120 days before mean SOSc and EOSc); six precipitation-related variables (total precipitation 10, 20, 30, 60, 90 and 120 days before mean SOSc and EOSc); and six sunlight-related variables (10, 20, 30, 60, 90 and 120 days of accumulated sunshine time before mean SOSc and EOSc).
A multiple linear regression model was used to identify the eighteen variables responsible for the shifts in SOSc and EOSc using R (R Development Core Team 2018). We used the R relaimpo package to evaluate the relative importance of the variables. To eliminate the effects of multicollinearity, only three variables (one warmth-related, one precipitation-related, and one sunlight-related) were used in each separate analysis. Each variable was standardized before performing linear regression analysis. 216 models were built for each SOSc and EOSc in each of the two sites, for a total of 864 models.
Results
Ring-width chronologies and response to climate
Table 1 summarizes the statistical characteristics of the ring width chronologies. The mean segment length of cores at site QLN was 84 years; for YNG it was 156 years. Mean sensitivity values were 0.30 and 0.16; inter-serial correlation values were 0.77 and 0.57. We have a reliable ring-width chronology for site QLN for the period 1894–2014. The chronology for site YNG runs from 1803 to 2016 (see Fig. 2 for both chronologies). Because we only had reliable instrumental climate records for the period 1960–2014, we have only used the ring-width records for the same period.

Observed (solid black line) and simulated (dotted gray line) chronologies of tree-ring width. r1 and r2 are the Pearson correlation coefficients for observed/simulated chronologies for both study sites. Correlation between observed and simulated chronologies was significant at a P < 0.01 level
Correlation analysis showed that at site QLN radial growth was positively and significantly correlated with precipitation in May, June, and the previous September (P < 0.05). No significant correlation with temperature was found (Fig. 3). At site YNG there was a significant correlation between radial growth and temperature in last autumn and from current May to July. There was also a significant correlation with precipitation in May (P < 0.05) (see Fig. 3).

Pearson correlation coefficients between observed tree-ring chronologies and observed monthly temperature (black bar) and total precipitation (gray bar) from previous September to current August. QLN is the study site close to the Qilian meteorological station; YNG is the study site close to Yeniugou station. Correlation significance levels (two-tailed test): *P < 0.05; **P < 0.01
Stem phenology and climate
Using the parameters listed in Table 2, we found highly significant (P < 0.01) positive relationships between the actual tree ring width chronologies and the simulated ones. Correlation coefficients between actual and simulated chronologies were 0.61 for both Qilian and Yeniugou meteorological stations, from 1960 to 2014 (Fig. 2), demonstrating that confidence in our V–S model simulation was not misplaced.
Stem radial growth in site QLN seems to be mainly sensitive to the availability of water, although during time periods at the beginning of growing season, temperature can become an important factor. Radial growth begins close to or in May for site QLN (Fig. 4). Growth remains high until the end of June, which we believe is due to net positive precipitation-evaporation in May and June. This is followed by a relatively dry period from June until the end of August, resulting in a drastic decrease in growth. Although there is again more available water in the fall, growth seems to be limited by temperature as the growing season draws to a close. 10 and 20 days before the EOSs in QLN, more than half of the days were temperature-limited. However, 30, 60, 90, and 120 days before the QNG EOSs, the majority of days were moisture-limited (Table 3). The SOSs in site QLN ranged from the middle April to early June; SOSs had a mean value of 142 ordinal days. The EOSs QLN ranged from 242 to 276 ordinal days. The mean value was 259 ordinal days.

The simulated daily mean radial growth response to: a sunlight (blue line), temperature (green line), soil moisture (red line), b these three factors combined (black line). Mean V–S model based SOSs and EOSs are indicated by arrows
At site YNG, tree-ring formation seems to be limited by temperature throughout the course of the growing season. However, water availability (from spring melt) seems to have some influence in early May (Fig. 4). 10, 20, 30, 60, 90, and 120 days before EOSs, more than 70% of the days were temperature-limited. Temperature seems to be the major factor influencing SOSs for both sites. 10, 20, 30, 60, 90, and 120 days before the date of SOSs, more than 50% of the days that were temperature-limited (Table 3). The average stem-growth season at site QLN is 117 days; at site YNG it is shorter, at 85 days. Average SOSs for YNG was 163 ordinal days. Average EOSs for YNG was 248 ordinal days.
Canopy phenology and climate
At site YNG, SOSc ranged from ordinal days 141 to 177. The earliest date was in 2004; the latest was in 1992. The earliest onset of canopy dormancy occurred in 1991 (day 258), and the latest in 1993 (day 287) (see Fig. 5). At site QLN, SOSc ranged from ordinal days 142–170. The EOSc at QLN ranged from ordinal days 263 to 289.

Characteristics of the SOSs and EOSs derived from the V–S model (solid black line) and the SOSc and EOSc derived from remote sensing data (dotted gray line). Thin black lines show linear trends, 1982–2014. r is the Pearson correlation coefficient between the SOSs and SOSc, and between EOSs and EOSc. P is the significance of their linear trend
Figure 6 shows correlations between SOSc, EOSc, and the eighteen explanatory factors used in our analyses. It is apparent that warmth factors have higher correlation coefficients with both QLN and YNG SOSc than do the precipitation and sunlight variables. Mean temperature at 90 days before the SOSc (T_90) seems to be the most influential factor at both sites (Table 4). Few of the eighteen variables showed any significant correlation with EOSc. The variance percentages correlated with those variables were low for both sites. Of the six sunlight variables, only two, when correlated with SOSc and EOSc, displayed P < 0.05 levels of significance.

Pearson correlation coefficients (r) between the remote sensing data based SOSc and EOSc and the eighteen variables. See Table 4 for an expansion of the abbreviations. See Fig. 1 for the locations of the two meteorological stations. Horizontal dotted lines indicate 95% confidence intervals of the correlation coefficients
Comparisons between stem and canopy phenology
Stem and canopy spring phenology showed similar patterns, as indicated by the significant correlation coefficients (0.46 and 0.56) between SOSs and SOSc at both QLN and YNG (Fig. 5). This may result from the same temperature impact factor (T_90) of SOSs and SOSc at both sites (Tables 3, 4). Significantly (P < 0.05) advancing trends were seen for the period 1982–2014 in SOSs at both sites. The significant earlier SOSc was seen at QLN. none of the linear trends in EOSc and EOSs at both sites were significant (P < 0.05).
Correlation coefficients between the EOSs and EOSc at both sites were low and did not reach a significant level (P < 0.05). This could be attributed to the long interval between EOSc and EOSs, and the different factors affecting EOSc and EOSs (Tables 3, 4). The interval EOSc-EOSs at QLN was 14.6 days on average. the interval EOSc-EOSs at YNG was 26.2 days on average. Temperature and moisture levels before EOSs had the greatest effect on the timing of EOSs at both sites. On the contrary, climate seems to play a little role in the variation of EOSc (Tables 3, 4). EOSc preceded EOSs at both sites.
Discussion
Impact of climate on canopy and stem phenology
SOS
Our research indicated that there is a significant correlation between the start of the start of stem radial growth season and canopy growth season at sites QLN and YNG during the period 1982–2014 (Fig. 5). Another study of cold-region black spruce also showed synchronism between xylem and bud formation phenology (Perrin et al. 2017). As the mean spring temperature in two sites was increasing (P < 0.01, ESM_1), SOSc and SOSs for two sites also increased (P < 0.05). Recent observational and experimental studies in cold regions suggest that global warming has led to earlier SOS in the Northern Hemisphere (Settele et al. 2014). A recent study (Tian et al. 2017) near site QLN has suggested that warmer soil temperatures at the start of the growing season could favor root activity. This would allow for the transport of more water and nutrients for xylem cell growth. Moisture-related variables were the second-most important driver of stem phenology in the spring (Table 3). Drought stress may delay the onset of cambial cell division at drought-prone forest sites (Ren et al. 2018). This relationship has also been observed (using remote-sensing) for the Tibetan Plateau as a whole. Variations in available moisture strongly affect the spring vegetation green-up date (Piao et al. 2011; Ding et al. 2016).
EOS
None of the three climatic variables (temperature, moisture, and sunlight) seem to have played important roles in variations in EOSc at either study site (Table 4). We infer that canopy growth may cease due to endogenous factors (Körner and Basler 2010; Cooke et al. 2012). Environmental influence on phenology may be marginal.
At both sites, the end of stem growth preceded the end of canopy growth. It is possible that, after the stem growth ended, the tree has to store sufficient photosynthetic products to sustain the plant over the winter and during the following spring. This hypothesis is supported by the fact that tree-ring width chronologies are significantly correlated with autumn climate conditions in the previous year (Fig. 3) (Liu et al. 2016).
Temperature was also a major factor in the timing of EOSs at site YNG (Table 3). This site is near the upper tree line. This result is in line with previous dendroclimatological studies of Qinghai spruce growing near the upper tree line (Liu et al. 2016). The major impact factors in the timing of EOSs in Qilian were alternatively temperature- or precipitation-related variables (Table 3). A similar result was found by Zhang et al. (2018), who studied Juniperus przewalskii growing on the southern slopes of the Qilian mountains: drought conditions seem to end xylem growth.
Implications for the V–S model used for stem phenology research
Traditional linear statistical analyses alone are not useful in suggesting a physical or biological mechanism for variability or change in the relationship between climate and tree growth. The radial-growth V–S model provides a new and more useful approach to explain stem phenology data. Still it must be admitted that there are some areas of uncertainty in the process-model simulations used in this study. The first problem is uncertainty as to the quality and accuracy of local precipitation data. YNG ring-width models were developed with input from the Yeniugou meteorological station, which is ~ 300 m lower in elevation. There may have been differences in precipitation between the site and the station. Second, the model itself is a very simple depiction of tree growth. It does not take into account other important inputs into tree growth, such as nutrient and CO2 availability. Tree-ring width is also influenced by tree age to some extent. These may be some of the reasons that variation in the V–S modeled SOSs and EOSs were much larger than those seen in remote sensing data for SOSc and EOSc (Fig. 5). It is to be hoped that there will be further studies of radial growth in Qinghai spruce over a longer observation period. Such studies will improve our understanding of the ecological and physiological characteristics of the spruce and allow us to predict how this species might cope with changing climate conditions.
Implications for forest growth
In recent decades the Chinese government has implemented several projects, such as the National Forest Conservation Program and the Grain for Green program, which have significantly increased vegetation coverage in the Qilian Mountains (Li 2004). Many semi-arid grasslands near the lower tree line have been converted to P. crassifolia plantation forests (Zhu et al. 2017). Our study has shown that rising spring temperatures (ESM_1) could start the growing season earlier, which might increase tree growth. However, since the 1980s there has been no significant increase in annual precipitation in Qilian station (ESM_1). Our V–S modeling results indicate that soil moisture is the most important factor limiting xylem cell differentiation during the June–August growing season at the lower-elevation QLN site (Fig. 4). (Note that according to Tian et al. (2017), major radial growth occurred in the June–July period.) Rising temperatures will likely increase water stress, which would complicate forest management (Yao et al. 2000; Lin et al. 2017).
Conclusion
This study compiled and analyzed the stem and canopy phenology of Qinghai spruce and their correlation with climate; the study was conducted at two sites, at different elevations, in the central Qilian Mountains, Northeast Tibetan Plateau. Both the NDVI-recorded canopy spring phenology and the V–S-modeled stem spring phenology are predominantly limited by temperature. These two events are linked; there were significant positive correlations between SOSc and SOSs at both sites. Canopy and stem autumn phenology are not correlated to the same degree. This may have been due to other factors that were not inputs to V–S model. The model did show its utility in its simulations of intra-seasonal radial-growth impact factors. Our results also showed that soil moisture is the major limiting factor for radial growth in the growing season at the lower elevation study site, QLN. The temperature is the major limiting factor for radial growth at the upper elevation study site, YNG. These results suggest that forest management must take elevation differences into account when facing the challenges of climate change.
Author contribution statement
XP participated in the experiment design, sample collection, data analysis, manuscript draft and revision. JD participated in the experiment design, data analysis, and manuscript draft. BY and SX participated in the design of the study and manuscript draft. GL participated in the experiment design and sample collection.
References
Ahl DE, Gower ST, Burrows SN et al (2006) Monitoring spring canopy phenology of a deciduous broadleaf forest using MODIS. Remote Sens Environ 104:88–95
Allen CD, Macalady AK, Chenchouni H et al (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684
Anchukaitis KJ, Evans MN, Kaplan A et al (2006) Forward modeling of regional scale tree-ring patterns in the southeastern United States and the recent influence of summer drought. Geophys Res Lett 33:347–360
Anderegg WRL, Kane JM, Anderegg LDL (2013) Consequences of widespread tree mortality triggered by drought and temperature stress. Nat Clim Change 3:30–36
Antonucci S, Rossi S, Deslauriers A et al (2015) Synchronisms and correlations of spring phenology between apical and lateral meristems in two boreal conifers. Tree Physiol 35:1086–1094
Biondi F, Waikul K (2004) DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput Geosci 30:303–311
Briffa KR, Jones PD (1990) Basic chronology statistics and assessment. In: Cook ER, Kairiukstis LA (eds) Methods of dendrochronology: applications in the environmental sciences. Kluwer Academic Publishers, Dordrecht, pp 137–152
Carnicer J, Mooney HA (2011) Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc Natl Acad Sci USA 108:1474–1478
Chang XX, Zhao WZ, He ZB (2014) Radial pattern of sap flow and response to microclimate and soil moisture in Qinghai spruce (Picea crassifolia) in the upper Heihe River Basin of arid northwestern China. Agric For Meteorol 187:14–21
Chen F, Yuan YJ, Wei WS (2011) Climatic response of Picea crassifolia tree-ring parameters and precipitation reconstruction in the western Qilian Mountains, China. J Arid Environ 75:1121–1128
Chen RS, Song YX, Kang ES et al (2014) A cryosphere-hydrology observation system in a small alpine watershed in the Qilian mountains of China and its meteorological gradient. Arct Antarct Alp Res 46:505–523
Cheng GD, Xiao HL, Fu BJ et al (2014) Advances in synthetic research on the eco-hydrological process of the Heihe River Basin. Adv Earth Sci 29:431–437 (in Chinese with English abstract)
Chmielewski FM, Rötzer T (2001) Response of tree phenology to climate change across Europe. Agric For Meteorol 108:101–112
Cooke JEK, Eriksson ME, Junttila O (2012) The dynamic nature of bud dormancy in trees: environmental control and molecular mechanisms. Plant Cell Environ 35:1707–1728
Cuny HE, Rathgeber CBK, Lebourgeois F et al (2012) Life strategies in intra-annual dynamics of wood formation: example of three conifer species in a temperate forest in north-east France. Tree Physiol 32:612–625
Deslauriers A, Rossi S, Anfodillo T et al (2008) Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol 28:863–871
Deslauriers A, Fonti P, Rossi S et al (2017) Ecophysiology and plasticity of wood and phloem formation. In: Amoroso MM, Daniels LD, Baker PJ et al (eds) Dendroecology tree-ring analyses applied to ecological studies. Springer International Publishing AG, Switzerland
Ding MJ, Li LH, Nie Y et al (2016) Spatio-temporal variation of spring phenology in Tibetan Plateau and its linkage to climate change from 1982 to 2012. J Mt Sci 13:83–94
Du J, He ZB, Yang JJ et al (2014) Detecting the effects of climate change on canopy phenology in coniferous forests in semi-arid mountain regions of China. Int J Remote Sens 35:6490–6507
Evans MN, Reichert BK, Kaplan A et al (2006) A forward modeling approach to paleoclimatic interpretation of tree-ring data. J Geophys Res Biogeosci 111:G03008
George SS (2014) An overview of tree-ring width records across the Northern Hemisphere. Quat Sci Rev 95:132–150
Grissino-Mayer HD (2001) Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree Ring Res 57:205–221
Jeong SJ, Chang-Hoi HO, Gim HJ et al (2011) Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob Change Biol 17:2385–2399
Jones OM, Kimball JS, Nemani RR (2014) Asynchronous Amazon forest canopy phenology indicates adaptation to both water and light availability. Environ Res Lett 9:124021
Jönsson P, Eklundh L (2004) TIMESAT—a program for analyzing time-series of satellite sensor data. Comput Geosci 30:833–845
Körner C, Basler D (2010) Phenology under global warming. Science 327:1461–1462
Kross A, Fernandes R, Seaquist J et al (2011) The effect of the temporal resolution of NDVI data on season onset dates and trends across Canadian broadleaf forests. Remote Sens Environ 115:1564–1575
Li WH (2004) Degradation and restoration of forest ecosystems in China. For Ecol Manag 201:33–41
Li XX, Liang EY, Gričar J et al (2013) Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau. Tree Physiol 33:48–56
Liang EY, Wang YF, Piao SL et al (2016) Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc Natl Acad Sci USA 113:4380–4385
Lin PF, He ZB, Du J et al (2017) Recent changes in daily climate extremes in an arid mountain region, a case study in northwestern China’s Qilian Mountains. Sci Rep 7:2245
Liu HY, Williams AP, Allen CD et al (2013) Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Glob Change Biol 19:2500–2510
Liu Y, Sun C, Li Q et al (2016) A Picea crassifolia tree-ring width-based temperature reconstruction for the Mt. Dongda region, Northwest China, and its relationship to large-scale climate forcing. PLoS One 11:e0160963
Melvin TM, Briffa KR (2008) A “signal-free” approach to dendroclimatic standardisation. Dendrochronologia 26:71–86
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42
Peñuelas J, Sardans J, Estiarte M et al (2013) Evidence of current impact of climate change on life: a walk from genes to the biosphere. Glob Change Biol 19:2303–2338
Perrin M, Rossi S, Isabel N (2017) Synchronisms between bud and cambium phenology in black spruce: early-flushing provenances exhibit early xylem formation. Tree Physiol 37:593–603
Pettorelli N, Vik JO, Mysterud A et al (2005) Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol Evol 20:503–510
Piao SL, Cui MD, Chen AP et al (2011) Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agric For Meteorol 151:1599–1608
Ren P, Rossi S, Camarero JJ et al (2018) Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann Bot 121:617–624
Rossi S, Deslauriers A, Griçar J et al (2008) Critical temperatures for xylogenesis in conifers of cold climates. Glob Ecol Biogeogr 17:696–707
Rossi S, Rathgeber CBK, Deslauriers A (2009) Comparing needle and shoot phenology with xylem development on three conifer species in Italy. Ann For Sci 66:206–206
Schwartz MD, Reed BC, White MA (2002) Assessing satellite-derived start-of-season measures in the conterminous USA. Int J Climatol 22:1793–1805
Settele J, Scholes R, Betts R et al (2014) Terrestrial and inland water systems. In: Field CB, Barros VR, Dokken DJ, Mach KJ, Mastrandrea MD, Bilir TE, Chatterjee M, Ebi KL, Estrada YO, Genova RC, Girma B, Kissel ES, Levy AN, MacCracken S, Mastrandrea PR, White LL (eds) Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Climate Change 2014: impacts, adaptation, and vulnerability. part a: global and sectoral aspects. Cambridge University Press, Cambridge, pp 271–359
Tian QY, He ZB, Xiao SC et al (2017) Response of stem radial growth of Qinghai spruce (Picea crassifolia) to environmental factors in the Qilian Mountains of China. Dendrochronologia 44:76–83
Vaganov EA, Hughes MK, Shashkin AV (2006) Growth dynamics of tree rings. Springer, Berlin
Wigley TML, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim 23:201–213
Yang X, Mustard JF, Tang J et al (2012) Regional-scale phenology modeling based on meteorological records and remote sensing observations. J Geophys Res Biogeosci 117:G03029
Yang B, Qin C, Wang JL et al (2014) A 3,500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau. Proc Natl Acad Sci USA 111:2903–2908
Yang B, He MH, Shishov V et al (2017) New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proc Natl Acad Sci USA 114:6966–6971
Yao TD, Liu XD, Wang NL et al (2000) Amplitude of climatic changes in Qinghai-Tibetan Plateau. Chin Sci Bull 45:1236–1243
Zhang GL, Zhang YJ, Dong JW et al (2013) Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proc Natl Acad Sci USA 110:4309–4314
Zhang QB, Evans MN, Lyu LX (2015) Moisture dipole over the Tibetan Plateau during the past five and a half centuries. Nat Commun 6:8062
Zhang JZ, Gou XH, Pederson N et al (2018) Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiol 38:840–852
Zhu X, He Z-B, Du J et al (2017) Temporal variability in soil moisture after thinning in semi-arid Picea crassifolia plantations in northwestern China. For Ecol Manag 401:273–285
Acknowledgements
The study was jointly funded by the National Natural Science Foundation of China (nos. 41701050, 41601051, 41520104005, 41325008), the CAS Light of West China Program, and the Foundation for Excellent Youth Scholars of the Northwest Institute of Eco-Environment and Resources, CAS.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Communicated by E. Liang.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Peng, X., Du, J., Yang, B. et al. Elevation-influenced variation in canopy and stem phenology of Qinghai spruce, central Qilian Mountains, northeastern Tibetan Plateau. Trees 33, 707–717 (2019). https://doi.org/10.1007/s00468-019-01810-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-019-01810-z