
Abstract
Key message
A global increase in temperature could potentially increase the trees’ growth at high altitude and decrease at low altitude. Another immediate consequence would be an increase of the altitude threshold where the inversion of tree growth response to temperature occurs.
Abstract
The first network of 18 tree-ring chronologies of Norway spruce (Picea abies) from the Eastern Carpathians (Romania) was studied in relation to the climatic factors and regional patterns in tree growth responses to climate. The sites are distributed along a latitudinal and altitudinal transect. The tree growth reaction to climate variability was analyzed by means of response functions. We used redundancy analysis (RDA) to identify regional patterns in the climatic response. The Norway spruce diameter growth patterns in Eastern Carpathians from Romania correspond to different climatic responses according to three elevation levels: low (≤1000 m a.s.l.); intermediate (1000–1300 m a.s.l.); high (≥1300 m a.s.l.). At high altitudinal level tree growth is strongly limited by summer temperatures. This climatic signal progressively decreases with decreasing altitude and increasing mean temperature. Tree growth at low elevation sites is controlled mainly by summer precipitations and in the intermediate elevation sites there is not any statistically significant correlation with climatic variables. At elevations of 1000–1100 m a.s.l., at a mean temperature of 13–13.5 °C in June and 15.5–16 °C in July, further increases in mean temperature result in an inversion of the relationship between tree-ring growth and temperature (i.e., the response becomes negative). A global increase in temperature could potentially increase the trees’ growth at high altitude and decrease at low altitude. Another immediate consequence would be an increase of the altitude threshold where the inversion of tree growth response to temperature occurs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Norway spruce is an important evergreen coniferous tree native to montane and boreal European forests, ranging from the European Alps to the Balkan and Carpathian Mountains, and extending north into Scandinavia and northern Russia (Barnes and Wagner 2004). It has shown good yield and quality performance under very different site conditions, producing high-quality timber. It is also ecologically valuable, providing protection and recreation forests in mountain regions in much of Europe (Skrøppa 2003). Norway spruce (Picea abies) is one of the most important conifer tree species not only in Europe, but also in Romania, where pure and mixed spruce stands cover over 28 % of the total forest area and over 55 % of the entire Eastern Carpathians forest area. It is one of the most important forest species in Romania as it covers many areas and is productive in wood of high quality (Ichim 1990).
Forests are a product of the geographic environment and crucially depend on climatic factors. Understanding the links between climate variations and forest ecosystems, as well as the impact of past climatic variations and current and predicted changes on forest vegetation, is essential to adapt management practices and strategies to ensure the sustainable use of forest ecosystems (Villalba et al. 1994; Tessier et al. 1997; Tardif et al. 2003; Andreassen et al. 2006). Sustainable management strategies for mountain forests must be based on the knowledge of the impact of environmental factors on forest ecosystems (Savva et al. 2006; Sidor 2011). Beside the uncertainties related to future climate change, a variety of biotic and abiotic factors influencing tree growth complicate our understanding of the interactions between forest ecosystems and climate variability. Within the geographical range of a species, environmental features vary in relation to elevation, creating spatial variability in the factors limiting growth, and consequently species sensitivity to climatic variability. However, the regional climate can be modified by site factors, such as slope, aspect and degree of exposure to wind, even at the same elevation (Barry 1992). A variety of climatic parameters must, therefore, be explored in more integrated approaches to understand the complex relations between climatic variability and tree physiological responses (Bűntgen et al. 2006).
Climatic conditions are likely to change, which will probably affect forest productivity. Increases in mean annual temperature of 0.5–1.5 °C by 2020–2029 compared with the years 1980–1990 and of 2.0–2.5 °C by 2090–2099 (IPCC 2007) on a global level may not seem relevant, although such high and rapid temperature rises are exceptional. At the local level, however, certain regions will be under significantly greater pressure than others.
The temporal assessment of growth–climate response patterns via the comparison of tree-ring chronologies and meteorological time series provides unique long-term understanding of the interplay between terrestrial ecosystems and external forcing agents (Carrer et al. 2007; Büntgen et al. 2008). The influence of future climatic changes on forest growth can only be assessed if species-specific responses are well understood with respect to changes in the ecological site conditions (Friedrichs et al. 2009).
More detailed studies at regional and sub-regional scales are needed to detect the local patterns of individual species responses and their main constraints (Peterson et al. 2002; Bunn et al. 2005). Norway spruce is one of the most significant European tree species and also a tree species with the highest number of various health and growth problems which have appeared in the last decades (Rybníček et al. 2010). The forest and timber industry in Romania is very important for the Romanian economy and one of the most important sources of income in rural areas (Abrudan et al. 2009), even more than in some Eastern European countries. The annual value of products and environmental services provided by Romanian forests are estimated to be around 3.1 billion US $ (Poynton et al. 2000). In many regions of Europe, it was reported that the relation climate–Norway spruce tree growth varies along an altitudinal range (Levanic et al. 2009—Southeastern European Alps; Savva et al. 2006—Tatra Mountains, Poland; Andreassen et al. 2006—Norway; Wilson and Hopfmueller 2001—Bavarian Forest, Germany; Cejkova and Kolar 2009—Sumava Mountains, Czech Republic). In Romania little is known about Norway spruces’ dendroclimatic regional variation and responses to climate change. Several dendroclimatological studies have investigated the relation between climate and radial trees’ growth (Popa 2003, 2005; Bouriaud and Popa 2007; Kern and Popa 2007; Sidor and Popa 2007; Sidor 2011), but only in forest stands located at the upper limit of current forest vegetation.
In this paper, we report a comparative analysis of the radial growth reactions to climatic variability of Norway spruce in a network of sites covering the natural altitudinal range of spruce in Romania’s Eastern Carpathians. The aims were to:
-
1.
investigate the radial growth responses of Norway spruce to climate variability;
-
2.
describe the regional patterns in these responses, focusing on the network of study sites;
-
3.
to highlight the main climatic factors that determine the regional variability in the growth of Norway spruce.
Materials and methods
Study area
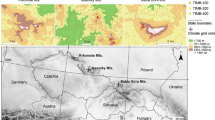
The study area is located in Romania, in the Eastern Carpathians (Fig. 1). The northern boundary is the border with Ukraine and the western the Transylvanian Depression. The eastern limit is one Suceava Plateau (between the Suceava and Moldova rivers), Moldova and the Curvature Subcarpathians, while the southern limit is the Prahova Valley (Roşu 1980).

Locations of the Norway spruce tree-ring chronologies network in the Eastern Carpathians, Romania (dark triangles indicate the eighteen study sites locations)
The vegetation differs according to altitudinal belt and latitudinal position. The vegetation belts decrease in elevation further north, where the conifers lower tree line are much lower (below 1000 m) than in the south (Doniţă et al. 1992). The Curvature Carpathians forest is dominated by beech stands (Fagus sylvatica) and mixed stands of Norway spruce with beech, the Moldova Carpathians by mixed stands of beech and conifers (mostly spruce and silver fir (Abies alba)) and the western (volcanic mountains) and northern regions by large tracts of coniferous forests (mostly Norway spruce and silver fir). Subalpine and alpine vegetation is found in Ciucas, Calimani and the Rodna Mountains (Doniţă et al. 1992).
The highest peaks in the Eastern Carpathians are located in the north (Rodna—2303 m a.s.l., Calimani—2100 m a.s.l.) while two peaks in central and southern areas are lower. The soil types are brown acid soils and andisols (on volcanic rocks), brown acid soils in the flysch area, spodosols in the crystalline-mesozoic area, brown soils in the depressions and low elevation mountains areas (Roşu 1980).
The Eastern Carpathians climate is influenced by latitude, elevation, and exposure and landscape fragmentation. In very high areas over 1800 m a.s.l. (Rodna, Maramures, Suhard, Calimani, Ciucas) the climate is mostly alpine and subalpine, but Baltic influences affect the northern mountain areas. Average annual temperatures differ significantly, decreasing at the same elevation from south to north and at the same location from the lowest to the highest elevation. For example, they decrease from 6 to 7 °C at 1000 m elevation to 0 °C at 1800 m elevation. Average annual rainfall increases with altitude, but is more pronounced on west exposed slopes, e.g., it is over 1200 mm in the Ceahlau mountains, because of the influence of the oceanic air masses carried by the western prevailing winds. In the extreme northern part of the Eastern Carpathians (Rodna and Maramures Mountains) due to the combined effects of the mountain position and elevation, and western prevailing winds, has the highest annual average precipitation (over 1300 mm) (Sandu et al. 2008).
Tree-ring sampling and chronology development
We sampled eighteen forest stands in a network consisting of both latitudinal and altitudinal transects, in the Eastern Carpathians. At each site we selected between 17 and 55 trees according to dendrochronological classical procedures. From each tree two cores were extracted at a height of 130 cm from the ground perpendicular to the slope to reduce the presence of compression wood (Stokes and Smiley 1968). The annual ring width was measured using Lintab equipment and TSAP software with a precision of 0.01 mm (Rinn 2005).
The ring width series were cross dated by means of TSAP, in which the series of individual radial growth were graphically compared with the series of average growth. The cross dating reliability was verified with COFECHA, through the correlation analysis for overlapping sub-periods of 50 years (Holmes 1983; Grissino-Mayer 2001).
All the individual growth series were standardized to eliminate the non-climate signals and maximize the climate information within the chronology. To remove age influence, a 67 % cubic smoothing spline function with a 50 % cutoff frequency was applied (Cook and Kairiukstis 1990). The chronology was obtained by means of the bi-weighted average. All tree-ring data were processed with ARSTAN for Windows (Cook and Krusic 2006). During the analysis, we used the residual dendrochronological series.
Descriptive statistics were calculated for each detrended chronology to allow for comparisons among the chronologies (Table 1). The mean sensitivity (MS) is an indicator of the relative changes in tree-ring width variance between consecutive years (Fritts 1976). The signal strength of each chronology was evaluated using the mean series intercorrelation (RBAR) (Cook and Kairiukstis 1990). RBAR is the mean correlation coefficient among tree-ring series (Briffa and Jones 1990). Expressed population signal (EPS) measures how well the finite-sample chronology compares to the theoretical population chronology based on an infinite number of trees. It varies from 0.0 to 1.0 and the rough cutoff point for accepting an EPS value considered to be 0.85 (Wigley et al. 1984). RBAR and EPS values were calculated using 30-year moving window with a 15-year overlap. A useful statistic parameter to emphasize the quality of the chronologies is the mean correlation coefficient between individual series and the mean series.
Climate data
The network of weather stations is small and climatic data are only available for a short period of time. Obtaining the data necessary for statistical analyses to support dendroclimatological investigations is difficult in the study area. The majority of weather stations have records only for the last 40–50 years. They are situated near towns and the elicitation of a complete data set is rather difficult. To produce a homogeneous set of climatic data for the entire area of study, with time series for the last century, we used the climatic database with a 0.5° × 0.5° resolution CRU TS 3 (Mitchell and Jones 2005). For each site, we extracted the set of climatic data (monthly temperatures and precipitations) from the nearest grid cell for the period 1901–2006, accessed via the KNMI climate explorer (http://www.climexp.knmi.nl).
Climate–growth relationship analysis
The influence of the climate on trees’ growth was assessed by means of the response functions over a 15-month period from June of the previous year (t-1) to August of the current year of growth (t) (Fritts 1976). The study period was set to 1901–2000 and was covered by all the series. The statistical calculations were carried out using DENDROCLIM2002 (Biondi and Waikul 2004).
Definition of the regional patterns
To identify regional patterns in the climatic response of Norway spruce, we used the redundancy analysis (RDA), analyzing the period 1901–2000 for which the statistical cover is guaranteed for all the series. The RDA allows the study of the relationship between two tables of variables X and Y. The RDA is a non-symmetric method, where the components extracted from X are such that they correlate as much as possible with the variables of Y. The components of Y are then extracted so that they correlate as much as possible with the components extracted from X. In RDA biplots, the correlation between biotic and abiotic variables is given by the cosine of the angle between two vectors. Vectors pointing in roughly the same direction indicate a high positive correlation, vectors crossing at right angles correspond to a near zero correlation, and vectors pointing in opposite directions show a high negative correlation (ter Braak and Prentice 1988). Environmental variables with long vectors are the most important in the analysis (Legendre and Legendre 1998). In this analysis, each chronology was regarded as a sample (site) and each monthly response function as a descriptor (Legendre and Legendre 1998). For the statistical quantification of the similitude of the dendrochronological series, we calculated the matrix correlation coefficients for the common period 1901–2000.
Maps showing the spatial correlation between tree-ring composite chronologies and climatic data were generated using the KNMI climate explorer (van Oldenborgh and Burgers 2005; http://www.climexp.knmi/nl). These maps permit evaluation of the representativeness of composite chronologies at a regional scale.
Results
Chronology statistics
We analyzed eighteen chronologies distributed, both latitudinal and altitudinal, in the Eastern Carpathians. Table 1 shows the geographical location and the statistical parameters of the eighteen dendrochronological series. The chronologies were elaborated by extracting 858 radial growth cores from 431 trees. The length of the radial growth series varies from 119 to 305 years. The value of the mean ring width ranges between 1.4 and 3.4 mm. Generally, the chronologies located at elevations lower than 1150 m have mean ring width values higher than 2 mm. The variation amplitude of the mean sensitivity values is low, with higher values for the low-elevation series, than for those at higher elevations. The average value of the residual dendrochronological series rbar is relatively high for all the series, with values ranging between 0.26 and 0.49. For the analysis period 1901–2000, all the chronologies have EPS values higher than 0.85. The mean correlation coefficient between the individual series and the mean series has values between 0.52 and 0.68.
General climate–growth response
The analysis of the tree-ring growth response of the Norway spruce to climatic variations (Fig. 2a) revealed which climatic parameters and months of the year had most influence. Temperature had more influence than precipitation at all elevations. For the stands at elevations higher than 900 m the July temperatures during the year preceding the formation of the tree ring were most significant in negatively affecting growth, whereas the sites at 800–1200 m a.s.l. the September temperatures of the previous year had most impact. For the stands at 1300–1650 m a.s.l. previous October temperatures positively influenced ring width growth, as did the previous December temperatures for trees at 600–1150 m a.s.l. Regarding the temperatures during the current growth year, the January ones are most significant for trees growing at 1200–1650 m a.s.l. (positive influence), and the April ones for trees at 1050–1150 m a.s.l. (positive influence). June temperatures had a negative correlation for the stands located at elevations lower than 900 m but both June and July temperatures had a positive correlation for stands at altitude higher than 1300 m a.s.l. The amount of precipitations in July of the previous year also had a significant positive influence on the growth of trees at 1000–1300 m a.s.l. whereas that in May of the current year positively influenced tree growth only at 1050–1060 m a.s.l.

General responses of the radial growth of trees to the variation in temperature and precipitation. For clarity we only show the values greater than ± 0.1: a response functions; b variation in the response functions in relation to elevation
The response of tree-ring growth to climatic differences varied with elevation (Fig. 2b). Temperatures during the previous October, current June and July correlated positively with tree growth in relation to elevation, while precipitation during previous August correlated negatively but not significantly. The strongest correlation of the response function coefficients to elevation (0,901) is for the temperatures in June during the current growth year (Fig. 2). At the same time, the response functions values in current June also present the highest amplitude. The negative values of the response functions in the case of previous October, current June and July temperatures change to positive at an elevation of approximately 1000–1100 m.
Spatial patterns of climate–growth responses
The first 2 RDA axes for the period 1901–2000 express 53.4 and 11.2 % of the total variation (Fig. 3), which suggest separating the dendrochronological series into two groups, in relation to their climatic response would make sense. The first group of series correlates negatively with axis 1. This group comprises the series located below 1300 m a.s.l., within which those below 1000 m a.s.l. have higher negative RDA values on axis 1, whereas the second group of series located higher than 1300 m a.s.l. has positive RDA values on axis 1. As for the climatic variables, the temperatures in the previous October and in current June and July are strongly positively correlated with axis 1, and clearly separate the series at high elevation from those at lower elevations. The August rainfall during the previous growth year has a higher positive RDA value on axis 2, the low-elevation series being separated in relation to the response to this climatic variable. The low-elevation series in the southern part of the Eastern Carpathians (The Curvature Carpathians) responded positively to the precipitation in previous August, while those in the northern part responded negatively.

Redundancy analysis for the chronologies network. For clarity, we present only the climatic variables with elevated values on the 2 RDA axes—over 0.9. T temperature; P precipitations; 1–12 = month of year (1 = January and so on); p the previous growth year
To verify whether the spatial patterns in the tree-ring response to climate vary with elevation, we calculated and analyzed a correlation coefficient matrix among the studied chronologies (Fig. 4). The highest correlation values are between the dendrochronological series in neighboring elevation areas with the highest and most significant correlations between ring width series at elevations higher than 1300 m a.s.l. At the same time, we found a relatively high correlation between the series from elevations lower than 1000 m a.s.l.. For correlation between all the dendrochronological series the most similar were the low and intermediate elevation series.

Matrix of coefficients of correlation among the residual dendrochronological series for the period 1901–2000
The RDA (Fig. 3) and correlation coefficient matrix (Fig. 4) show that the tree-ring width index chronologies follow 3 variation models corresponding to the three elevation levels (low: ≤1000 m a.s.l.; intermediate: 1000–1300 m a.s.l.; high: ≥1300 m a.s.l.). Three regional dendrochronological series were, therefore, developed, one for each elevation level (Fig. 5). The average correlation coefficient is lowest to the low-elevation series (r = 0.665), and highest for the high-elevation series (r = 0.810). The inter-annual variation of the regional chronologies also differs for the low-elevation series, particularly in extreme years (Fig. 5). The intermediate elevation series has the lowest variation amplitude. Even if the correlation of low- and high-elevation chronologies with climate series is inverse the chronologies show common negative pointer years (e.g., 1928, 1947, 1958, 1971, 1989).

Regional dendrochronological series. The gray line represents the site dendrochronological series, and the thick line the regional chronologies at each altitude
The spatial correlation among the regional series with average temperatures in June–July was positive for the high-elevation series, negative for the low-elevation series and not significant for the intermediate series.
The temperatures in June and July of the current year were the climatic variables which accounted for most differences in the climatic response between the tree-ring growth series (Figs. 3, 4), expressed graphically in relation to the response functions in Figs. 5, 6 and 7 show their relationship. Generally, the sites where the average temperatures in June and July were higher than the regional mean responded negatively to the variation in June and July temperatures, but positively to lower temperatures. The thresholds for the change from climatic positive to negative response were 13–13.5 °C in June and 15.5–16 °C in July, at an average elevation of 1000–1100 m a.s.l..

Spatial correlation of the series of regional growth indices with temperatures in the June–July period of the year corresponding to the formation of the annual ring. Elevation chronologies: a low, b intermediate and c high

Mean temperatures in June (a) and July (b) in relation to the response functions
Discussions
We applied commonly used dendroecological and dendroclimatological methods to analyze climate–growth relationships for Norway spruce stands in Eastern Carpathians. These methods can help to identify long-term natural responses of plants because the length of tree-ring series provides an extended context for assessing changes in tree growth (Knapp et al. 2001), in particular for defining the historical range of variability of tree growth–climate relationships at multiple temporal and spatial scales (Swetnam et al. 1999). Even if the before-1950s gridded dataset CRU is often lacking local points from the region, we preferred to use it to produce a homogeneous set of climatic data for the entire area of study, with time series for the last century. Another reason is the fact that the network of weather stations from the study area has climatic data available only for the last 40–50 years and they are situated near towns and the elicitation of a complete data set is rather difficult.
The Norway spruce diameter growth patterns in Eastern Carpathians from Romania correspond to different climatic responses according to three elevation levels: low (≤1000 m a.s.l.); intermediate (1000–1300 m a.s.l.); high (≥1300 m a.s.l.). Such different climatic responses corresponding to different elevation levels have also been found for other regions from Europe. In the Bavarian Forest, in Germany, the climatic response of the Norway spruce diameter growth differed as well according to three elevation levels: low (≤680 m a.s.l.), intermediate (780–970 m a.s.l.) and high (≥1070 m a.s.l.) (Wilson and Hopfmueller 2001). In Switzerland, Norway spruce tree-ring growth has been found to correlate strongly with climate at the lower and higher elevation boundaries of the forest, whereas at intermediate elevations diameter growth in stands is primarily influenced by local factors (Lingg 1986). In Austrian Alps, tree growth at low altitudes is controlled mainly by spring–summer moisture availability, at high altitudes the radial growth is strongly influenced by summer temperatures and in the intermediate altitudinal range the diameter growth does not have a direct relationship with climatic variables (Leal et al. 2007).
Temperature is one of the main factors governing radial growth processes in Norway spruce (Makinen et al. 2002). In the Eastern Carpathians from Romania at the high altitudinal level, tree growth is strongly limited by summer temperatures. This climatic signal progressively decreases with decreasing altitude and increasing mean temperature. Tree growth at low-elevation sites is controlled mainly by summer precipitations and in the intermediate elevation sites there is not any statistically significant correlation with climatic variables. The temperature in June during the growing season had most impact on the radial growth of the Norway spruce. The temperatures in current July and the previous October also influence tree growth, but less. Physiological, earlywood formation depends partially on stored photosynthates from the previous year(s), whereas latewood formation relies essentially on current year photosynthates (Helle and Schleser 2004). In the studied area, the influence of June and July temperatures from growing seasons have a positive influence at elevations higher than 1300 m a.s.l. and a negative influence at elevations lower than 1000 m a.s.l.. At higher elevations the warm temperatures from June and July have a positive influence on latewood formation, whereas at lower elevations the warm temperatures from June and July determine a reduced latewood width. Similar results at elevations >1300 m a.s.l. and between 1000 and 1300 m a.s.l. have also been obtained in the Tatra Mountains in Poland (the highest mountains in the whole Carpathian Arc). There, however, chronologies of trees at low elevations between 800 and 1000 m a.s.l. responded positively to June temperatures (Savva et al., 2006). In the Bavarian Forest, the temperatures in June and July during the current growing season are positively correlated to the growth of the Norway spruce trees at elevations higher than 1070 m a.s.l., and negatively to the stands lower than 680 m a.s.l.. The temperature in June during the growing season is also the period of maximum growth of Norway spruce in the Alps (Tranquillini 1979; Deslauriers et al. 2003; Makinen et al. 2003). Thus, tree-ring growth in Norway spruce seems to depend on the temperature in the early part of growing season as Rossi et al. (2008) claim. In Norway, the temperature regime in June of the current year influenced the diameter growth of Norway spruce stands located at high elevations in most regions, except for the south-eastern part, where the radial growth of the Norway spruce is mainly influenced by June precipitation (Andreassen et al. 2006). In the European Alps, the radial growth of Norway spruce at high elevations responds positively to the higher temperatures during the period June–August of the current year (Frank and Esper 2005; Levanic et al. 2009). Also, we found persistent correlations between the tree-ring width chronologies and winter temperatures (December at low elevations, January at high elevations). Even if in that period the trees are definitely in dormancy, the repeated correlations found provide evidence that the winter temperatures could be a driving factor. Such correlations with winter temperatures have also been found for other tree-ring chronologies from the region (Koprowski 2013; Churakova et al. 2014). A possible explanation of these influences of winter temperatures could be due to the winter frost impacts on rooting system and water and mineral nutrient uptake of trees. Soil-frost events may influence the dynamics of fine roots and, therefore, affect root-derived C fluxes to the soil (Gaul et al. 2008). Jackson et al. (1997) estimated that fine roots consume approx. 30 % of the global net primary production. Also, Häussling et al. (1988) highlighted the important role of root growth in water and mineral nutrient uptake. In Eastern Carpathians from Romania the precipitation signals on tree-ring growth of the Norway spruce are weak and inconsistent across altitudes. Comparatively, in Bavaria below 700 m a.s.l., the radial growth of Norway spruce positively responded to precipitation during the growing season, while higher temperatures during spring and summer had a negative influence (Dittmar and Elling 1999; Wilson and Hopfmueller 2001).
In the Eastern Carpathians at elevations of 1000–1100 m a.s.l., at a mean temperature of 13–13.5 °C in June and 15.5–16 °C in July, further increases in mean temperature result in an inversion of the relationship between tree-ring growth and temperature (i.e., the response becomes negative). Radial growth in Norway spruce responds positively to the mean 1901–2006 June temperatures for the sites with temperature below 13–13.5 °C, and negatively if the mean June temperatures are higher. Negative responses at low elevations to June temperature may also be induced by water stress (Jolly et al. 2005). Barber et al. (2000), Wilmking et al. (2004) and Lloyd and Fastie (2002) show that in the northern latitudes Norway spruce radial growth in the twentieth century has decreased with increasing temperature. In July, the radial growth response to climate changes from negative to positive with a mean temperature of 15.5–16 °C. This temperature threshold corresponds in the study area to an average elevation of 1000–1100 m a.s.l. for both the June and July temperatures. This threshold at elevation of 1000–1100 m is probably related with the soil moisture. At elevations lower than 1000–1100 m, warm temperatures have a negative effect on ring width in absence of sufficient soil moisture. In Norway, the diameter growth response to climatic variation changes from negative to positive with a mean June temperature of 12–13 °C, at an elevation of 400–500 m a.s.l. in south-east Norway (Andreassen et al. 2006). In northwestern part of Canada, a July–August average temperature of 11.3 °C was identified to be the threshold for decline in a tree-ring record along an elevational tree line (D’Arrigo et al. 2004).
Our results confirm the ones obtained also in other studies (Woodward 1987; Bonan and Shugart 1989; Villalba et al. 1994; Makinen et al. 2002), that the radial growth of Norway spruce is generally limited by low summer precipitation at low elevation, and by low summer temperature at high latitudes and elevations. In Eastern Carpathians from Romania, a global increase in temperature could potentially increase the growth of trees at high altitude and decrease at low altitude. Another immediate consequence would be an increase of the altitude threshold where the inversion of tree growth response to temperature occurs.
Conclusions
The results obtained in the present study seem to be similar to those found in similar studies in Europe. This will contribute to substantiate findings regarding the main climate responses in diameter growth of the Norway spruce in the entire elevation gradient across its current distribution in the Eastern Carpathians. These findings help to understand the radial growth responses of Norway spruce to climate variability in the twentieth century and to identify the mechanistic drivers in climate change which will impact tree growth processes in the region. Identifying the spatial and ecological patterns is important for management decisions on species distribution, as well as for productivity projections, and understanding regional variation. The increase in mean temperature during the growing season forecasted in most climate change scenarios will induce significant changes in tree growth, with stronger growth in trees growing higher than 1100 m a.s.l. and less growth in trees growing below 1000 m a.s.l.
The forest sector in Romania contributes to between 3.5 and 4.5 % of the gross national product (GNP), and is thus economically very important. Norway spruce is the main species used in the timber industry in Romania. Thus, future changes in its productivity due to increases in air temperature will have significant impact on Romania’s forest economy. The differences in the impact of climate change on the growth of Norway spruce at high and low elevations need to be taken into account in deciding on Romania’s forest management policy.
References
Abrundan IV, Marinescu V, Ionescu O, Ioras F, Horodnic SA, Sestras R (2009) Developments in the Romanian forestry and its linkages with other sectors. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 37(2):14–21
Andreassen K, Solberg S, Tveito OE, Lystad SL (2006) Regional differences in climatic responses of Norway spruce growth in Norway. For Ecol Manage 222:211–221
Barber VA, Juday GP, Finney BP (2000) Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 405:668–673
Barnes BV, Wagner WH (2004) Michigan Trees: a guide to the trees of the Great Lakes region. University of Michigan Press, Ann Arbor, p 439
Barry RG (1992) Mountain climatology and past and potential future climatic changes in mountain regions: a review. Mt Res Dev 12:71–86
Biondi F, Waikul K (2004) DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput Geosci 30:303–311
Bonan GB, Shugart HH (1989) Environmental factors and ecological processes in boreal forests. Annu Rev Ecol Syst 20:1–28
Bouriaud O, Popa I (2007) Site and species influence on tree growth response to climate in Vrancea Mountains. Proc Rom Acad 9(1):63–72
Briffa KR, Jones PD (1990) Basic chronology statistics and assessment. In: Cook ER, Kairiukstis LA (eds) Methods of Dendrochronology: applications in the environmental sciences. International Institute for Applied Systems Analysis (IIASA). Kluwer Academic Publishers, Dordrecht, pp 137–152
Bunn AG, Waggoner LA, Graumlich LJ (2005) Topographic mediation of growth in high elevation foxtail pine (Pinus balfouriana Grev. et Balf.) forests on the Sierra Nevada. USA Glob Ecol Biogeogr 14:103–114
Buntgen U, Frank D, Schmidhalter M, Neuwirth B, Seifert M, Esper J (2006) Growth/climate response shift in a long subalpine spruce chronology. Trees 20:99–110
Büntgen U, Frank D, Wilson R, Carrer M, Urbinati C, Esper J (2008) Testing for tree-ring divergence in the European Alps. Glob Change Biol 14:2443–2453
Carrer M, Nola P, Eduard JL, Motta R, Urbinati C (2007) Regional variability of climate–growth relationships in Pinus cembra high elevation forests in the Alps. J Ecol 95:1072–1083
Cejkova A, Kolar T (2009) Extreme radial growth reaction of Norway spruce along an altitudinal gradient in the Sumava Mountains. Geochronometria 33:41–47
Churakova Sidorova OV, Eugster W, Zielis S, Cherubini P, Etzold S, Saurer M, Siegwolf R, Buchmann N (2014) Increasing relevance of spring temperatures for Norway spruce trees in Davos, Switzerland, after the 1950s. Trees 28:183–191
Cook ER, Kairiukstis LA (1990) Methods of dendrochronology. Applications in the environmental sciences. Kluwer Academic Publishers, Dordrecht, p 394
Cook ER, Krusic PJ (2006) ArstanWin 4.1.b_XP. http://www.ideo.columbia.edu
D’Arrigo RD, Kaufmann RK, Davi N, Jacoby GC, Laskowski C, Myneni RB, Cherubini P (2004) Thresholds for warming-induced growth decline at elevational tree line in the Yukon Territory Canada. Glob Biogeochem Cycle 18:GB3021. doi:10.1029/2004GB002249
Deslauriers A, Morin H, Urbinati C, Carrer M (2003) Daily weather responses of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Quebec (Canada). Trees 17:477–484
Dittmar C, Elling W (1999) Jahrringbreite von Fichte und Buche in Abhaengigkeit von Witterung und Hoehenlage. Forstwissenschaftliches Centralblatt 118:251–270
Donită I, Ivan D, Sanda V, Popescu A (1992) Romanian vegetation. Agricultural Technical Publishing, Bucharest 407 pp
Frank D, Esper J (2005) Temperature reconstructions and comparisons with instrumental data from a tree-ring network for the European Alps. Int J Climatol 25(11):1437–1454
Friedrichs DA, Buntgen U, Frank DC, Esper J, Neuwirth B, Loffler J (2009) Complex climate controls on 20th century oak growth in central-west Germany. Tree Physiol 29:39–51
Fritts HC (1976) Tree Rings and Climate. Academic Press, pp 567
Gaul D, Hertel D, Leuschner C (2008) Effects of experimental soil frost on the fine-root system of mature Norway spruce. J Plant Nutr Soil Sci 171:690–698
Grissino-Mayer HD (2001) Assessing crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree Ring Res 57:205–221
Häussling M, Jorns CA, Lehmbecker G, Hecht-Buchholz C, Marschner H (1988) Ion and water uptake in relation to root development in norway spruce (Picea abies (L.) Karst.). J Plant Physiol 133(4):486–491
Helle G, Schleser H (2004) Beyond CO2-fixation by Rubisco - an interpretation of 13C/12C variations in tree rings from novel intra-seasonal studies on broad-leaf trees. Plant Cell Environ 27:367–380
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bulletin 43:69–75
Ichim R (1990) Ecological management of the Norway spruce forests. Ceres Publishing Bucharest, Bucharest, p 186
IPCC (2007) Climate change 2007: the physical science basis. In: Solomon S et al (eds) Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Jackson RB, Mooney HA, Schulze ED (1997) A global budget for fine root biomass, surface area, and nutrient contents. Proc Nat Acad Sci 94:7362–7366
Jolly WM, Dobbertin M, Zimmermann NE, Reichstein M (2005) Divergent vegetation growth response to the 2003 heat wave in the Swiss Alps. Geophys res Lett 32(18):L18409
Kern Z, Popa I (2007) Climate–growth relationship of tree species from a mixed stand of Apuseni Mts., Romania. Dendrochronologia 24(2–3):109–115
Knapp PA, Soule PT, Grissino-Mayer HD (2001) Detecting potential regional effects of increased atmospheric CO2 on growth rates of western juniper. Glob Change Biol 7:903–917
Koprowski M (2013) Spatial distribution of introduced Norway spruce growth in lowland Poland: the influence of changing climate and extreme weather events. Quatern Int 283:139–146
Leal S, Melvin TM, Grabner M, Wimmer R, Briffa KR (2007) Tree ring-growth variability in the Austrian Alps: the influence of site altitude, tree species and climate. Boreas 36:426–440
Legendre P, Legendre L (1998) Numerical ecology, 2 English edn. Elsevier Science BV, Amsterdam, p 853
Levanic T, Gricar J, Gagen M, Jalkanen R, Loader NJ, McCarrol D, Oven P, Robertson I (2009) The climate sensitivity of Norway spruce in the southeastern European Alps. Trees 23:169–180
Lingg W (1986) Dendrookologische Studien an Nadelbaumen im alpinen Trockental Wallis (Schweiz). Eidg Anst forstl VersWes 287:1–81
Lloyd AH, Fastie CL (2002) Spatial and temporal variability in the growth and climate response of treeline trees in Alaska. Clim Change 52:481–509
Makinen H, Nojd P, Kahle HP, Neumann U, Tveite B, Mielikainen K, Rohle H, Spiecker H (2002) Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. For Ecol Manage 171:243–259
Makinen H, Nojd P, Kahle HP, Neumann U, Tveite B, Mielikainen K, Rohle H, Spiecker H (2003) Large-scale climatic variation and radial increment variation of Picea abies (L.) Karst. in central and northern Europe. Trees 17:173–184
Mitchell TD, Jones PD (2005) An improved method of constructing a database of monthly climate observations and associated high-resolution grids. Int J Climatol 25:693–712
Peterson DW, Peterson DL, Ettl GJ (2002) Growth responses of subalpine fir to climatic variability in the Pacific Northwest. Can J For Res 32:1503–1517
Popa I (2003) Comparative analysis of Norway spruce (Picea abies (L.) Karst.) and silver fir (Abies alba Mill.) dendroclimatological response in the northern Eastern Carpathians. Bucovina. Forestry 11(2):3–14
Popa I (2005) Historical reconstruction of the thermal dynamics regime of June in Rodna Mountains. For J 4:21–28
Poynton S, Mitchell A, Ionascu G, Mc Kinnenn F, Elliott J, Abrudan IV (2000) Economic evaluation and reform of the Romanian forestry sector. Editura Pentru viata, Brasov
Rinntech (2005) TSAP User reference, pp 110
Rossi S, Deslauriers A, Gričar J, Seo JW, Rathgeber C, Anfodillo T, Morin H, Levanic T, Oven P, Jalkanen R (2008) Critical temperatures for xylogenesis in conifers of cold climates. Glob Ecol Biogeogr 17:699–707
Rosu A (1980) Romanian Geography. Didactic and Pedagogical Publishing, Bucharest 484 pp
Rybníček M, Čermák P, Žid T, Kolář T (2010) Radial growth and health condition of Norway spruce (Picea abies (l.) Karst.) stands in relation to climate (Silesian Beskids, Czech Republic). Geochronometria 36:9–16
Sandu I, Poiană I, Pescaru V (eds) (2008) Romanian climate. Romanian Academy Publishing, Bucharest, p 365
Savva Y, Oleksyn J, Reich P, Tjoelker MG, Vaganov EA, Modrzynski J (2006) Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland. Trees 20:735–746
Sidor C (2011) The relationship between climate and radial growth of trees from upper mountain area. Editura Silvică, pp 196
Sidor C, Popa I (2007) Comparative analysis of Norway spruce, silver fir and Scots pine dendroclimatological response in the Curvature Carpathians. For J 3:3–8
Skrøppa T (2003) EUFORGEN Technical Guidelines for genetic conservation and use for Norway spruce (Picea abies). International Plant Genetic Resources Institute, Roma 6 pp
Stokes MA, Smiley TL (1968) An introduction to tree-ring dating. University of Chicago Press, Chicago, p 110
Swetnam TW, Allen CD, Betancourt JL (1999) Applied historical ecology: using the past to manage for the future. Ecol Appl 9:1189–1206
Tardif J, Camarero J, Ribas M, Gutierrez E (2003) Spatiotemporal variability in tree growth in the central Pyrenees: climatic and site influences. Ecol Monogr 73(2):241–257
Ter Braak CJF, Prentice IC (1988) A theory on gradient analysis. Adv Ecol Res 18:271–317
Tessier L, Guibal F, Schweingruber FH (1997) Research strategies in dendroecology and dendroclimatology in mountain environments. Clim Change 36(3–4):499–517
Tranquillini W (1979) Physiological ecology of the alpine timberline: tree existence at high altitudes with special reference to the European Alps. Springer-Verlag, Berlin, p 137
Van Oldenborgh G, Burgers G (2005) Searching for decadal variations in ENSO precipitation teleconnections. Geophys Res Lett 32(15). doi: 10.1029/2005GL023110
Villalba R, Veblen TT, Ogden J (1994) Climatic influence on the growth of subalpine trees in the Colorado front range. Ecology 75:1450–1462
Wigley TML, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim Appl Meteorol 23:201–213
Wilmking M, Juday GP, Barber VA, Zald HJ (2004) Recent climate warming forces contrasting growth responses of white spruce at treeline in Alaska through temperature thresholds. Glob Change Biol 10:1724–1736
Wilson RJ, Hopfmueller M (2001) Dendrochronological investigations of Norway spruce along an elevational transect in the Bavarian Forest. Ger Dendrochronol 19(1):67–79
Woodward FI (1987) Climate and Plant Distribution. Cambridge University Press, Cambridge
Author contribution statement
Cristian Gheorghe SIDOR and Ionel POPA conceived the ideas, collected the data, analyzed the data and wrote the paper. Radu VLAD and Paolo CHERUBINI contributed to the data analyses and to the writing of the paper.
Acknowledgments
The work was supported by CNCS-UEFISCDI project number PN-II-RU-TE-2011-3-0040. We are grateful to Silvia Dingwall for editing and revising the usage of English language.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by E. Liang.
Rights and permissions
About this article

Cite this article
Sidor, C.G., Popa, I., Vlad, R. et al. Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees 29, 985–997 (2015). https://doi.org/10.1007/s00468-015-1178-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-015-1178-3