
Abstract
Key message
Across sites in Brazil and Hawaii, LMA and N mass were strongly correlated with height and shade index, respectively, which may help simplify canopy function modeling of Eucalyptus plantations.
Abstract
Within tree canopies, leaf mass per area (LMA) and leaf nitrogen per unit area (N area) commonly increase with height. Previous research has suggested that these patterns occur as a strategy to optimize carbon gain by allocating available resources to upper canopy leaves that are exposed to greater light availability. We tested three hypotheses about the influences of height, shade index (a proxy for light), and stand age on LMA and leaf nitrogen for even-aged Eucalyptus saligna and Eucalyptus grandis × urophylla plantations in Brazil and Hawaii, USA, spanning most of the environmental conditions found across 19.6 million ha of Eucalyptus spp. plantations around the world. Shade index was developed by incorporating canopy depth (inner-crown shading) and a tree height ratio relative to neighbor trees (shading from other trees). Across all sites and ages, leaf height accounted for 45 % of the variation in LMA, whereas shade index accounted for only 6 %. A combination of both factors was slightly better in accounting for LMA variation than height alone. LMA–height relationships among sites were strongest under greater light availability and in older stands. Leaf nitrogen per unit mass (N mass) consistently decreased with shade index, whereas N area showed no consistent pattern with height or shade index. These relationships indicate that N mass is primarily driven by light, while height is the primary driver for LMA. The general relationships between LMA and leaf N mass across all sites may simplify canopy function modeling of E. saligna and E. grandis × urophylla plantations.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Two fundamental leaf traits, leaf mass per unit area (LMA, g m−2) and leaf nitrogen per unit area (N area, g m−2), correlate strongly with photosynthetic capacity and dark respiration rate and arguably represent the most useful plant traits for predicting physiological function (Ellsworth and Reich 1993; Cavaleri et al. 2008). Taken together, both LMA and N area may allow for simplification of modeling used to scale processes such as canopy gas exchange and vegetation productivity across large spatial extents (Ellsworth and Reich 1993; Reich et al. 1999; Hanson et al. 2004; Cavaleri et al. 2010). Tree species across a broad range of forest types display a consistent pattern of increasing LMA from the bottom to the top of the canopy (Hutchison et al. 1986; Marshall and Monserud 2003; Koch et al. 2004; Woodruff et al. 2004; England and Attiwill 2006; Ishii et al. 2008; Mullin et al. 2009; Cavaleri et al. 2010), and N area scales with LMA in relation to height (Ellsworth and Reich 1993). Researchers have theorized that foliar responses to light gradients involve allocation of carbon and nitrogen toward the tops of canopies to optimize use of solar energy (Field 1983; Bond et al. 1999). This theory is primarily based on structural and biochemical changes in leaf traits observed along a vertical canopy gradient.
These patterns of LMA relate to gradients in light through canopies (Ellsworth and Reich 1993; Kenzo et al. 2006), but other factors may actually drive the pattern, such as the gravity potential gradient (Koch et al. 2004; Woodruff et al. 2004; Cavaleri et al. 2010), exposure (Sack et al. 2006), or a combination of these factors (Burgess and Dawson 2007). The light hypothesis assumes that light interception in the upper portions of the canopy is the primary driver of leaf functional and morphological gradients (Ellsworth and Reich 1993). Support for this hypothesis stems from early experiments showing leaves that developed in high light availability in greenhouses and growth chambers (‘sun leaves’) tended to have higher LMA as a result of stacked and/or longer palisade cells when compared with the LMA of leaves in lower light (‘shade leaves’) (Nobel et al. 1975; Nobel 1977; Smith and Nobel 1978).
In some experiments, LMA correlated more strongly with leaf height than to light environment (Ishii et al. 2008; Cavaleri et al. 2010), perhaps as a direct result of changes in water relations within the canopy (Koch et al. 2004; Woodruff et al. 2004; Cavaleri et al. 2010). Xylem water potential decreases with height as a result of increasing path length and the gradient of decreasing water potential with height due to gravity (Scholander et al. 1965; Hellkvist et al. 1974; Koch et al. 2004; Woodruff et al. 2004; Ishii et al. 2008). Decreasing xylem water potential lowers leaf turgor pressure high in the canopy (unless countered by osmotic adjustment; Woodruff et al. 2004). Greater hydrostatic tension, or more negative water potential, reduces turgor pressure, which in turn limits cell expansion and growth during leaf development in upper canopies (Hsiao 1973; Woodruff et al. 2004; Meinzer et al. 2008). One consequence of limited cell expansion could be more densely packed mesophyll cells, causing greater leaf density and thus higher LMA. This theory is generally supported among tall conifers (Koch et al. 2004; Ishii et al. 2008; Mullin et al. 2009) and within a tropical forest stand containing high biodiversity (Cavaleri et al. 2010).
Nitrogen per unit area (N area) follows a similar trend to LMA with canopy height, while nitrogen per unit mass (N mass) often remains constant with height and light gradients (Hollinger 1989; Ellsworth and Reich 1993; Anten et al. 1998). This pattern may result from increasing LMA with light (Ellsworth and Reich 1993; Anten et al. 1998). Nitrogen is an essential component of many molecules involved in photosynthesis, including protein–pigment complexes in the thylakoid membrane, the product of the electron transport chain (NADPH), and soluble proteins involved in the Calvin–Benson cycle (esp. RUBISCO) (Evans 1989). Leaves with higher LMA and greater photosynthetic capacity per unit leaf area may benefit from higher nitrogen (Evans 1989). Vertical patterns in nitrogen allocation are not constrained by water relations and should respond to light availability gradient within canopies to optimize carbon gain (Field 1983; Hirose and Werger 1987).
As trees grow taller and older, structure and function change markedly, including increases in LMA (Yoder et al. 1994; Thomas and Winner 2002), decreases in height growth (Ishii et al. 2000; Bond et al. 2007), hydraulic conductance (McDowell et al. 2002b), total crown photosynthesis, midday leaf water potential (Barnard and Ryan 2003), transpiration (England and Attiwill 2007), and the ratio of leaf area to sapwood (McDowell et al. 2002a). For Eucalyptus regnans, older, taller trees had leaves that were more xeromorphic (thicker leaves and cuticle, smaller leaf area, and greater LMA) and may contribute to declining transpiration with age (England and Attiwill 2006, 2007). In extensive reviews, Thomas and Winner (2002) and Ryan et al. (2006) concluded that an increase in LMA with tree age was the most consistent change among species. The mechanisms behind the increase in LMA with tree age may include hydraulic limitations and subsequent reductions in leaf expansion, and altered gene expression caused by selection pressures such as exposure to greater irradiance, wind, herbivores, and pathogens (Thomas and Winner 2002).
We combined data from many earlier analyses of Eucalyptus saligna and Eucalyptus grandis × urophylla plantations from study sites in Brazil and Hawaii to test for within-crown patterns of LMA and N area. By investigating very uniform stands with little structural or genetic diversity (including single-age clonal plantations), we were able to avoid many confounding factors. E. saligna is native to Australia and has been found to attain heights of 42 m after 10 years and 82 m after 50 years at sites in Hawaii (Walters 1973; Burns and Honkala 1990). Eucalyptus grandis × urophylla was developed by the Aracruz pulp company for the purposes of combining high wood quality of Eucalyptus grandis and wood density and canker resistance of Eucalyptus urophylla (Gominho et al. 2001). Due to its fast growth and yield of valuable forest product, Eucalyptus spp. are one of the most widely planted hardwoods in the world (Turnbull 1999; Thomas et al. 2009). Eucalyptus spp. plantations cover more than 19.6 million hectares globally with about 20 % found in Brazil (Iglesias-Trabedo and Wilstermann 2008). The objectives of the study were to investigate the influence of leaf height, shade index (a proxy for light environment), and tree age on LMA, N area, and N mass for young E. saligna and E. grandis × urophylla stands within and among six study sites in Brazil and one in Hawaii, testing three hypotheses:
-
1.
LMA relates more strongly with height than with shade index (a proxy for light environment) because of the dominance of height relationships in other studies.
-
2.
LMA increases with tree age (beyond the effect of height alone) because stands experience dramatic changes in structure and function as trees age.
-
3.
Light is the primary driver of N area as a result of greater allocation of nitrogen under high light conditions to optimize carbon gain.
Materials and methods
Study sites
A study from Hawaii (HAW) with Eucalyptus saligna Sm. seed-origin trees was joined with six clonal-tree Eucalyptus grandis (W. Hill ex Maid) × urophylla (S.T. Blake) experiments in Brazil. Six of the study sites in Brazil (ARA, CEN, IP, SUZ, VCP, VER) were part of the Brazil Eucalyptus Potential Productivity (BEPP) Project (Stape et al. 2010). Results are also given for site–year combinations; for example, VER-3 refers to site VER measured in year 3.
The study site in Hawaii (HAW) was upslope from Peepekeo (19o50′N, 155o7′W) at an elevation of 350 m on a deep thixotropic, isothermic Typic Hydrudands in the Kaiwiki series. The mean annual temperature during the 6-year rotation (1994–2000) was 21 °C, with 3,400 mm year−1 of precipitation ensuring no water limitation on tree growth. This site contained stands of two age classes: 1.5 and 5.5 years. This first stand was planted in 1994 with 3 × 3 m spacing. An additional stand was planted 4 years later with 3 × 3 m spacing.
The ARA site was located near Aracruz in Espirito Santo, Brazil at 19°49′S, 40°05′W. Mean annual temperature (MAT) during the rotation at ARA was 24 °C and mean annual precipitation (MAP) was 1,360 mm year−1. Trees were planted in March 2001 with 3 × 3 m spacing. The CEN site was located near Guanhaes in Minas Gerais, Brazil (18°35′S, 42°59′W) on an Oxisol, where MAT was 19.4 °C and MAP was 1,108 mm year−1. Trees were planted in April 2004 with 3 × 3 m spacing. The IP site was located near Mogi Guacu in São Paulo, Brazil (22°21′S, 46°58′W) on an Oxisol, where MAT was 21.6 °C and MAP was 1,317 mm year−1. Trees were planted in October 2000 with 3 × 2.8 m spacing. The SUZ site was near Teixeiras de Freitas in Bahia (18o 20′S, 39 52′W) on an Ultisol with about 25 % clay. MAT during the experiment averaged 25 °C, 1,350 mm year−1 precipitation. Trees were planted in December 2001 with 3 × 3 m spacing. The VCP site was located near Luis Antonia in São Paulo, Brazil at 21°32′S, 48°22′W, where MAT was 22 °C and MAP was 1,307 mm year−1. Trees were planted in March 2000 with 3 × 3 m spacing. The Veracel site (VER) was located near Eunapolis in Bahia, Brazil at 16°21′S, 39°34′W, where MAT was 23.0 °C and MAP was 1,433 mm year−1. Trees were planted in March 2001 with 3 × 3 m spacing. For this study, trees were sampled during one (ARA, IP, VCP, SUZ), two (VER and HAW), or three (CEN) years at each site (Table 1) on different plots within the overall experimental plantation.
Experimental design and measurements
Six plots each were studied at sites ARA, IP, VER, and SUZ. At ARA and SUZ, three of the six plots were heavily fertilized (three times annually), irrigated, and uniform with all trees planted on the same day; and the other three plots were heavily fertilized, irrigated, and heterogeneous in structure with 1/3 of trees planted on day 0, 1/3 planted 40 days later, and the remaining 1/3 planted on day 80. At IP, two plots were heavily fertilized, irrigated, and uniform; two plots were heavily fertilized, irrigated, and heterogeneously planted. The remaining two plots were moderately fertilized, not irrigated, and uniformly or heterogeneously planted. At VER, three plots were moderately fertilized (operational levels of fertilization, see Stape et al. 2010), not irrigated, and uniform, and the three other plots were moderately fertilized, not irrigated, and heterogeneous. For sites listed above, each plot had 12 rows and 12 columns of trees, and every fourth tree of the central 36 trees in each plot was selected for destructive sampling (9 trees per plot × 6 plots per site = 54 trees per site, Table 1). At VCP, 24 trees from 11 plots were chosen for sampling, spread across all treatments to provide a wide variety of trees. Four plots were heavily fertilized, irrigated, and uniformly planted. Four plots were moderately fertilized, not irrigated, and uniformly planted. The remaining three plots were not fertilized, received no irrigation, and uniformly planted. Stape et al. (2010) provide additional information for BEPP sites including details about standard company fertilization and quantities of supplemental irrigation. At site HAW, two plots (one 1.5-year-old, and one 5.5-year-old) were sampled, with 13 destructively sampled trees in the younger plot and 24 trees in the older plot. Both plots were heavily fertilized (4 times annually).
Measurements for trees at all sites included total tree height and height to the base of the live crown. Destructively sampled canopies were divided into thirds based on crown length. Actual leaf height was estimated as the center of each relative canopy position, calculated using total tree height and height to base of live crown (height to lowest live foliage). All leaves were stripped from each tree and weighed in the field by canopy third. One bulked sample (of more than 100 leaves) was taken from each canopy position and returned to the laboratory for LMA and leaf N analyses. LMA was measured in all sites, while foliar N was only measured in IP-3, VCP-3, and VER-3. Leaf area was measured using a bench-top optical area meter (LI-3100, Licor, Inc. Lincoln, NE). Leaves were dried at 70 °C for 24 h and weighed to determine dry weight and subsequent estimates of LMA. Dried leaves were ground and analyzed for nitrogen (N) concentration by combustion furnace (NA1500 CHN Analyzer, Carlo Erba Strumentazione, Milan, Italy). Leaf nitrogen was either expressed as N mass (g kg−1) or calculated as N area (g m−2) using LMA measurements. Hawaii plot procedures were handled similarly to those of the Brazil plots (Barnard and Ryan 2003).
Statistical analysis and shade index
We developed a multiple regression model selection procedure for each site for predicting LMA, N mass, and N area based on the lowest AIC (Akaike Information Criterion) value (Anderson 2008). Independent variables included leaf sample height, shade index, age, and interactions among these variables. Analysis of variance (ANOVA) was used to determine significant main and interaction effects, with α = 0.05. Statistical analyses were conducted using R statistical software (R Core Team 2012). We also pooled all data across age and site to discern any emergent relationships, and ANOVA was used to determine significant main effects, with α = 0.05. For analysis where all data were pooled together, the interaction term (height × shade index) was ignored for the purposes of identifying the relative contribution of shade index and height toward a more parsimonious model of predicting LMA, N mass, and N area across all sites.
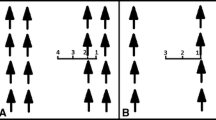
The actual light environment of each leaf sample location was not measured directly, so we developed a shade index (SI) that incorporates estimates of within-canopy shading (canopy depth) and shading of neighboring trees (weighting factor based on tree height relative to mean tree height; Fig. 1). Canopy depth in deciduous and conifer plantations has been found to be strongly and negatively related to light availability (Leverenz et al. 1982; Gower and Norman 1991; Rodrigo et al. 2001) and was used as an indicator of light availability (Iversen and Norby 2008). Shade index was calculated as:

For each tree, the canopy was divided into thirds, and both leaf height and canopy depth were estimated using the center of each third based on tree height and base to live crown measurements. Shade index was developed by calculating canopy depth (tree height–leaf height), and multiplying by a weighting factor (ratio of mean tree height:tree height) to account for shading from neighbor trees. In this example, leaf height was at the lower canopy, but we calculated leaf height for leaf samples collected at three canopy positions (upper, middle, and lower) for each tree
Where CD is canopy depth (m) and is calculated as [tree height–leaf height]; HTmean (m) is mean tree height at each site; and HTtree (m) is total tree height (Fig. 1). A higher ratio of mean tree height to total tree height indicates that a tree is shorter relative to mean tree height and would experience lower light availability due to shading from neighboring trees.
To identify any issues with collinearity, we plotted shade index versus leaf sample height for all sites and calculated variance inflation factor (VIF) for each relationship (Table 2). The VIF quantifies the influence of multicollinearity among independent variables on the standard errors. A VIF value >10 indicates serious collinearity, while values near one indicate no collinearity. Shade index was negatively correlated with sample leaf height at all sites (Table 2). Most sites contained a low VIF, but four sites (VER-3, CEN-2, CEN-10.5, HAW-1.5) had issues of collinearity.
To further separate out the effects of height and shade index, we grouped data into three bands of height (low, mid and high) and plotted LMA versus shade index and vice versa for three bands of shade index (low, mid, and high). Group categories were selected by separating height and shade index into three groups of equal size based on rank (smallest to largest). For the pooled data (across sites and ages), we compared the contribution of shade index and height to the full model using partial r 2 values. To further tease apart the effects of height and shade index on N area for pooled data, we plotted the residuals of shade index vs. N area against height and the residuals of height vs. N area against shade index.
Results
Leaf mass per area
Sample leaf height (hereafter referred to as height) was highly significant in best-fit models for LMA at all sites (Table 3). Shade index was included in best-fit models for LMA at all sites and was significant at four of six sites (Table 3). The shade index × height interaction was significant in one of the models (ARA) (Table 3). Age effects were tested at three sites that contained two or three age classes (VER, CEN, and HAW; Table 3). At VER, all possible parameters and interactions were included in the best-fit model. Main effects, shade index, height, and age, and the interaction effects, height × age and shade index × height × age, were significant (Table 3). At CEN and HAW, height, age, and height × age interaction were included in the model. Shade index was also included in the HAW model. The height and height × age interaction was significant at HAW (Table 3).
LMA increased linearly with height at all sites within single-age classes (IP, VCP, SUZ, and ARA), whereas LMA increased (IP), decreased (SUZ), or did not change (VCP) with shade index (Fig. 2). At ARA, LMA consistently increased with height within all shade index classes (Fig. 2g), and LMA increased with shade index at the mid height class but did not change with shade index at the low or high height class (Fig. 2h).

Relationship between LMA (g m−2) and height (m) (a, c, e) and shade index (b, d, f) for sites with non-significant height × shade index interaction. Relationship between LMA (g m−2) and height by levels of shade index (g) and shade index by levels of height (h) for the ARA site that had a significant height × shade index interaction. See Table 3 for significance of main effects and interactions. See Fig. 4 for R 2 values at ARA
For sites that contained more than one age class of stands (CEN, VER, and HAW), relationships between LMA and height form non-overlapping point clouds for each age (Fig. 3). LMA increased with height at the low and mid shade index class at VER-3 and VER-6 (Figs. 3a, 4a). LMA did not increase with height at the younger HAW (HAW-1.5) and CEN (CEN-2) sites, whereas LMA increased with height for the older HAW (HAW-5.5) and CEN (CEN-6.4, -10) sites (Fig. 3b, c).


R 2 values for relationships between LMA, N mass, and N area and height within classes of low, mid, and high shade index classes (a, c, e) and for relationships between LMA, N mass, and N area and shade index within classes of low, mid, and high height classes (b, d, f). R 2 values for non-significant relationships were not included in the figure. Negative signs above each bar indicate negative linear relationships between LMA, N mass, or N area and height or shade index. Bars without a negative sign indicate positive relationships
Height explained more variation in LMA within classes of shade index compared to shade index within classes of height, as indicated by the consistently higher R 2 values (Fig. 4). With a few exceptions, relationships between LMA and height were generally stronger at low and mid shade index classes (greater light availability) compared to the high shade index class (lower light availability; Fig. 4a). Furthermore, relationships between LMA and height were much stronger at older sites compared to younger sites at VER and HAW (Fig. 4a). Relationships between LMA and shade index tended be stronger at low to mid height classes compared to the high height class (Fig. 4b). We observed one significant relationship between LMA and shade index at the high height class (Fig. 4b).
Leaf nitrogen
Shade index was selected for leaf N mass best-fit models for all sites (Table 3). Height was selected in models at IP and VCP (Table 3). A significant interaction between shade index and height was also included in the model at VCP (Table 3). At VER, shade index was the only variable selected in the model (Table 3). N mass and height were positively correlated at IP and VER-3 (Fig. 5a, c), but negatively correlated within low and mid shade index classes at VCP (Figs. 4c, 5e). N mass and shade index were negatively correlated at IP and VER-3 (Fig. 5b, d) and negatively correlated within low and mid height classes at VCP (Figs. 4d, 5f).

Relationship between N mass (mg g−1) and height (m) (a, c) and shade index (b, d) for sites with non-significant height × shade index interaction. Relationship between N mass (mg g−1) and height by levels of shade index (e) and shade index by levels of height (f) for the VCP site that had a significant height × shade index interaction. See Fig. 4 for R 2 values at VCP
N mass was more strongly correlated with shade index within height classes compared to height within classes of shade index (Fig. 4c,d). This was most apparent in the low to mid height classes, where relationships between N mass and shade index were stronger compared to high height classes (Fig. 4d). Relationships between N mass and height were generally stronger within low and mid shade index classes compared to the high shade index class (Fig. 4c).
For leaf N area, shade index was selected and significant in models at all sites (Table 3). Height was included in all models and significant in the models for IP and VER (Table 3). N area increased with height at IP (Fig. 6a), but decreased with shade index (Fig. 6b). At VER-3, N area decreased with height within the high shade index level (Figs. 6c, 4e), and no changes with shade index within height classes were observed (Figs. 6d, 4f). At VCP, N area decreased with height within the low shade index class (Figs. 6e, 4e), and N area decreased with shade index within the low and mid height class (Figs. 6f, 4f). We did not observe any general trends between N area and height within shade index classes and between N area and shade index within height classes.

Relationship between height and leaf N area (mg m−2) and height (m) (a) and shade index (b) for the IP site with non-significant height × shade index interaction. Relationship between N area (mg m−2) and height by levels of shade index (c, e) and shade index by levels of height (d, f) for the VER-3 and VCP sites that had a significant height × shade index interaction. See Fig. 4 for R 2 values at VER-3 and VCP
Overall patterns across sites
Despite differences of LMA–height profiles among sites, we observed a significant relationship between LMA and height when data from all sites were pooled (Fig. 7a). Further analysis revealed that the partial r 2 for height was much greater compared to the partial r 2 for shade index (Table 4). Once height was taken into account, adding light to the model only explained 0.01 % additional variability in LMA. We observed a strong and negative relationship between N mass and shade index (Fig. 7b). Also, the partial r 2 for adding shade index into the full model was much greater compared to the addition of height (Table 4). The partial r 2 value for adding height into the full model for predicting N area was slightly greater compared to partial r 2 for shade index (Table 4). The relationship between height and residuals (shade index vs. N area) was positively significant, whereas the relationship between shade index and residuals (Height vs. N area) was not significant (Fig. 7c, d).

Relationship between LMA and height (a), leaf N mass and shade index (b), residuals of N area vs. shade index and height (c), and residuals of N area vs. height and shade index (d) for all sites pooled. Height and shade index were strongly correlated with LMA and N mass, respectively, across all sites and age classes in Brazil and Hawaii
Discussion
Leaf mass per area
These experiments supported our first hypothesis that leaf morphology of E. saligna and E. grandis × urophylla clonal plantations responds more strongly to leaf height than shade index (light environment) within and across all sites, consistent with previous findings in temperate conifers and tropical trees (Koch et al. 2004; Ishii et al. 2008; Mullin et al. 2009; Cavaleri et al. 2010). LMA showed a positive linear relationship with height within and across all sites. Xylem water potential routinely decreases linearly with height due to gravity (−0.01 MPa m−1) and path length (Scholander et al. 1965; Koch et al. 2004). Increasing tension with height due to gravity and path length has been linked to reduced leaf turgor pressure (Woodruff et al. 2004; Meinzer et al. 2008), and reduced cell expansion limits leaf development and may produce leaves with greater LMA. For E. saligna and E. grandis × urophylla, these mechanisms likely explain the linear increase in LMA with height.
While shade index was less influential than height, it did improve the fit of models within each site. We provide evidence that variation in LMA is primarily accounted for by height within broadly defined light conditions. Recent investigations of the effects of light and height on leaf morphology have concluded that leaf morphology tends to be more responsive to light availability below a threshold in the lower portions of forest canopies where light is limited, while hydraulic limitations are most important in the upper canopy of trees (Ishii et al. 2008; Mullin et al. 2009; Cavaleri et al. 2010). Light appeared to have slight, but statistically significant effects on the relationship between LMA and height for E. saligna and E. grandis × urophylla. Relationships between LMA and shade index did not reveal a threshold, but further analysis of LMA–height relationships within shade index classes show that these relationships tend to be stronger at higher light availability. Mullin et al. (2009) reached a similar conclusion where shoot mass per area of Sequoia sempervirens correlated strongly with height for foliage near the tops of trees, but not at mid-crown and base-of-crown canopy positions. The effects of hydraulic limitations may only be discernible under fully saturated light availability where leaves may be predisposed to additional factors such as lower turgor pressure.
Surprisingly, we found that LMA frequently decreased with increasing light availability in low to mid height classes. Consistent with our results, Anten et al. (1998) found that LMA of plants decreased with increasing light availability for low to mid height classes. Anten et al. (1998) attributed the decrease in LMA with increasing light availability to changing light availability during the course of the growing season. Leaves that developed later in the growing season under lower light availability had a lower LMA than leaves growing below them that developed early in the growing season under high light availability. For fast-growing evergreen, E. grandis, the mean leaf life span is 140–150 days, and tree height growth is 5.8–6.2 m year−1 (Laclau et al. 2009). Thus, it is plausible that the observed decrease of LMA with increasing light availability in low to mid height classes may be attributed to increasing light attenuation by dominant trees and subsequent declines in light availability for subordinate trees during the course of the study period. In other words, E. saligna and E. grandis × urophylla grow so fast that leaves lower in the canopy may experienced very different growth conditions during development months prior to current growing conditions. This also may contribute to inconsistent relationships observed for LMA and shade index. Nouvellon et al. (2010) reached a similar conclusion for 2- to 4-year-old E. grandis × urophylla that displayed little variation in LMA among three crown sections (upper, middle, and lower), and suggested that leaf acclimation of lower canopy leaves to increasingly shadier conditions following leaf development cannot occur.
These experiments provide partial support for our second hypothesis: increasing tree age leads to greater LMA. At VER, LMA at the younger site rarely exceeded 120 g m−2, whereas LMA at the older site displayed many values above 120 g m−2. A similar theme was observed at CEN and HAW. Our results support previous findings of increasing LMA with tree age, including E. regnans and E. grandis (Yoder et al. 1994; King 1999; Thomas and Winner 2002; England and Attiwill 2006). However, we also observed many LMA values at the older sites within or below the range of values at the younger sites. An important finding from this study is that the height–LMA relationships were much stronger at older sites compared to the younger sites where tree crowns rapidly increase in height. At the VER site, this observation was apparent for LMA–height relationships within greater light conditions (low to mid shade index), which provides further support of our conclusion that hydraulic limitations on leaf development are more important when leaves are light saturated. More importantly, our results provide evidence that as trees grow taller, hydraulic limitations within high light conditions may be accentuated, which is likely attributed to greater hydrostatic tensions at greater heights and longer path length from roots to leaves. England and Attiwill (2006) reached a similar conclusion for E. regnans, which displayed decreases in leaf area and increases in LMA and thickness with tree age and total tree height.
As E. saligna and E. grandis × urophylla stands age and develop, stronger height–LMA relationships for the older stands may also emerge as a result of positive feedback of whole-tree light absorption of dominant versus suppressed trees on leaf development. At the HAW site, Ryan et al. (2004) found that canopies progress from a homogenous canopy stratum (1.5 year) to a more complex vertical arrangement (5.5 year) in combination with leaf area distribution skewed toward the top canopy position for older stands. The progression from a homogeneous stratum toward a more complex vertical arrangement at the HAW site may contribute toward stronger height–LMA relationships observed at the older site (HAW-5.5). Dominant and taller Eucalyptus trees that outcompete neighbor trees within plantations are able to absorb more light and use light more efficiently (Binkley et al. 2010), which may be a consequence of and also facilitate the production and maintenance of costly tissues such as leaves with a greater LMA. This alternative conclusion is further supported by King (1999) who found that Eucalyptus grandis leaves with greater LMA maximize biomass production in larger trees but not for smaller trees, which tend to maximize biomass production through leaves with lower LMA. Whole-tree light absorption rather than light availability at the leaf level may also contribute to variation in LMA within a stand. We do not dispute the evidence of hydraulic limitations on leaf development, but concede to the possibility that multiple mechanisms may be working simultaneously during leaf development.
Across all sites and age classes, LMA and height maintained a strong positive correlation, whereas shade index appeared to contribute little to variation in LMA. The height–LMA relationship observed in tree canopies may simplify scaling of leaf-level photosynthesis and respiration to whole-canopy photosynthesis and respiration (Ellsworth and Reich 1993; Cavaleri et al. 2010). Despite the broad range of environmental conditions (irrigated and not irrigated, heavy to moderate fertilization) spanning the study sites, we provide evidence of a general pattern of increasing LMA with height for E. saligna and E. grandis × urophylla plantations. The strong relationship between LMA and height across the Brazil and Hawaii sites could greatly improve modeling efforts of canopy function over larger spatial scales, perhaps with the help of remote sensing (i.e., LiDAR, MODIS land information). This may be particularly important for Eucalyptus spp. because these plantations cover approximately 20 million ha of land globally. The global height–LMA relationship that we provide may simplify future research that aims to estimate carbon cycling in these plantations.
Leaf nitrogen
Previous research has suggested that nitrogen is allocated within light gradients as a strategy to maximize carbon gain (Field 1983; Hirose and Werger 1987). In support of our third hypothesis, we observed a general pattern where leaf nitrogen on a mass basis (N mass) consistently decreased with shade index, whereas leaf nitrogen on an area basis (N area) decreased, increased, or did not change with shade index. Barnard and Ryan (2003) also observed greater nitrogen concentration in leaves at the top canopy position of Eucalyptus saligna trees in Hawaii. While the current study did not directly measure light conditions, shade index can be considered a proxy for light conditions in the plantations as it incorporates both within-crown and inter-crown shading.
Canopy light gradients have been implicated as a strong driving force for N area in tree and plant canopies, likely due to the strong influence of light on LMA (used to convert N mass to N area, N mass * LMA = N area) (Ellsworth and Reich 1993; Bond et al. 1999). Previously reported strong relationships between area-based leaf nitrogen, photosynthesis, RUBISCO nitrogen, chlorophyll content, and electron transport capacity within species suggests that leaf nitrogen plays a critical role in the capacity of a leaf to assimilate carbon (Evans 1983; 1986; 1989). Consistent with other studies, we found that N area generally increased with height or decreased with shade index (Ellsworth and Reich 1993; Bond et al. 1999), but our results do not show that N area always increased with height within levels of shade index. In another study that compared leaf N area and N mass distribution within plant canopies, N area was primarily determined by light availability, while N mass appeared to be determined by leaf age (Anten et al. 1998).
Across all sites (pooled data), height appeared to be slightly more important than shade index as indicated by the greater partial r 2 of N area vs. height. This conclusion was further supported by relationships (or lack of) between residuals (N area vs. shade index) and height and residuals (N area vs. height) against shade index. Height was a better predictor of the departure from the trend of N area vs. shade index compared to shade index of the departure from the trend of N area vs. height. Thus, the increasing trend of LMA with height across all sites likely explains the positive relationship between N area and height.
In contrast, N mass consistently decreased with shade index (increased with light) which contrasts many studies that have found no changes in N mass along a light gradient. Our study provides strong evidence that leaf N mass is primarily driven by light. An important finding for this study was that N mass was more strongly correlated with shade index compared to N area and that shade index was more important than height when data were pooled across all sites. Each site also had different levels of fertilization treatment (zero, moderate, or heavy fertilization), yet there was an apparent general trend of N mass with shade index. Most sites either received heavy fertilization or standard operational fertilization, and previous work at these sites showed that tree growth did not respond to more intense fertilization (Stape et al. 2010). Our research suggests that scaling canopy processes from stand level to a larger spatial scale for Eucalyptus spp. may benefit from the use of mass-based measures of photosynthesis and nitrogen.
Conclusions and implications
Leaf height strongly affected LMA in E. saligna and E. grandis × urophylla plantations located in Brazil and Hawaii, consistently increasing with height across and within levels of shade index. Increasing hydrostatic tension with height due to gravity and path length is a likely mechanism that restricts leaf development and subsequent increases in LMA with height. In contrast, N mass was primarily affected by shade index (a proxy for light availability). N area did not consistently increase with height, but was consistently greater when shade index was lowest (i.e., exposure to light was highest). We conclude that the primary driver for LMA is height, while the primary driver for leaf nitrogen is light. Our results provide evidence that LMA and N area are not distributed optimally with respect to light and carbon gain, but the distribution of N mass appears to be distributed based on light availability. LMA and N area appear to be distributed with respect to height and subsequent water potential gradients, and our conclusion supports recent findings in temperate conifers and tropical trees where restrictions on water transport dictates leaf morphology. As a result of these restrictions, leaf morphological characteristics that determine LMA (thicker cuticle and leaves, smaller leaf area) appear to be distributed to reduce water loss rather than to optimize carbon gain.
These results also show that accounting for broadly defined light conditions and age can further improve predictions of LMA gradients in even-aged E. saligna and E. grandis × urophylla plantations at a regional scale. Furthermore, we provide evidence of a general relationship between LMA and height for E. saligna and E. grandis × urophylla. This relationship could simplify modeling of forest productivity of Eucalyptus spp. plantations which currently provide an increasing, worldwide supply of wood products.
References
Anderson DR (2008) Model based inference in the life sciences: a primer on evidence. Springer, New York, London
Anten NPR, Miyazawa K, Hikosaka K, Nagashima H, Hirose T (1998) Leaf nitrogen distribution in relation to leaf age and photon flux density in dominant and subordinate plants in dense stands of a dicotyledonous herb. Oecologia 113:314–324
Barnard HR, Ryan MG (2003) A test of the hydraulic limitation hypothesis in fast-growing Eucalyptus saligna. Plant Cell Environ 26:1235–1245
Binkley D, Stape JL, Bauerle WL, Ryan MG (2010) Explaining growth of individual trees: light interception and efficiency of light use by Eucalyptus at four sites in Brazil. Forest Ecol Manag 259:1704–1713
Bond BJ, Farnsworth BT, Coulombe RA, Winner WE (1999) Foliage physiology and biochemistry in response to light gradients in conifers with varying shade tolerance. Oecologia 120:183–192
Bond BJ, Czarnomski NM, Cooper C, Day ME, Greenwood MS (2007) Developmental decline in height growth in Douglas-fir. Tree Physiol 27:441–453
Burgess SSO, Dawson TE (2007) Predicting the limits to tree height using statistical regressions of leaf traits. New Phytol 174:626–636
Burns RM, Honkala BH (1990) Silvics of North America. U.S Department of Agriculture, Forest Service, Washington
Cavaleri MA, Oberbauer SF, Ryan MG (2008) Foliar and ecosystem respiration in an old-growth tropical rain forest. Plant Cell Environ 31:473–483
Cavaleri MA, Oberbauer SF, Clark DB, Clark DA, Ryan MG (2010) Height is more important than light in determining leaf morphology in a tropical forest. Ecology 91:1730–1739
Ellsworth DS, Reich PB (1993) Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia 96:169–178
England JR, Attiwill PM (2006) Changes in leaf morphology and anatomy with tree age and height in the broadleaved evergreen species, Eucalyptus regnans F. Muell. Trees-Struct Funct 20:79–90
England JR, Attiwill PM (2007) Changes in sapwood permeability and anatomy with tree age and height in the broad-leaved evergreen species Eucalyptus regnans. Tree Physiol 27:1113–1124
Evans JR (1983) Nitrogen and photosynthesis in the flag leaf of wheat (Triticum aestivum L.). Plant Physiol 72:297–302
Evans JR (1986) The Relationship between carbon-dioxide-limited photosynthetic rate and ribulose-1,5-bisphosphate-carboxylase content in two nuclear-cytoplasm substitution lines of wheat, and the coordination of ribulose-bisphosphate-carboxylation and electron-transport capacities. Planta 167:351–358
Evans JR (1989) Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 78:9–19
Field C (1983) Allocating leaf nitrogen for the maximization of carbon gain: leaf age as a control on the allocation program. Oecologia 56:341–347
Gominho J, Figueira J, Rodrigues JC, Pereira H (2001) Within-tree variation of heartwood, extractives and wood density in the Eucalypt hybrid Urograndis (Eucalyptus grandis × E. urophylla). Wood Fiber Sci 33:3–8
Gower ST, Norman JM (1991) Rapid estimation of leaf-area index in conifer and broad-leaf plantations. Ecology 72:1896–1900
Hanson PJ, Amthor JS, Wullschleger SD, Wilson KB, Grant RF, Hartley A, Hui D, Hunt ER, Johnson DW, Kimball JS, King AW, Luo Y, McNulty SG, Sun G, Thornton PE, Wang S, Williams M, Baldocchi DD, Cushman RM (2004) Oak forest carbon and water simulations: model intercomparisons and evaluations against independent data. Ecol Monogr 74:443–489
Hellkvist J, Richards GP, Jarvis PG (1974) Vertical gradients of water potential and tissue water relations in Sitka spruce trees measured with pressure chamber. J Appl Ecol 11:637–667
Hirose T, Werger MJA (1987) Maximizing daily canopy photosynthesis with respect to the leaf nitrogen allocation pattern in the canopy. Oecologia 72:520–526
Hollinger DY (1989) Canopy organization and foliage photosynthetic capacity in a broad-leaved evergreen montane forest. Funct Ecol 3:53–62
Hsiao TC (1973) Plant responses to water stress. Annu Rev Plant Phys 24:519–570
Hutchison BA, Matt DR, Mcmillen RT, Gross LJ, Tajchman SJ, Norman JM (1986) The architecture of a deciduous forest canopy in eastern Tennessee, USA. J Ecol 74:635–646
Iglesias-Trabedo G, Wilstermann D (2008) Eucalyptus universalis. Global cultivated eucalypt forests map 2008. Version 1.0.1. In: GIT Forestry Consulting’s EUCALYPTOLOGICS. Retrieved from www.git-forestry.com [March 29th 2009]
Ishii H, Reynolds JH, Ford ED, Shaw DC (2000) Height growth and vertical development of an old-growth Pseudotsuga-Tsuga forest in southwestern Washington State, USA. Can J Forest Res 30:17–24
Ishii HT, Jennings GM, Sillett SC, Koch GW (2008) Hydrostatic constraints on morphological exploitation of light in tall Sequoia sempervirens trees. Oecologia 156:751–763
Iversen CM, Norby RJ (2008) Nitrogen limitation in a sweetgum plantation: implications for carbon allocation and storage. Can J Forest Res 38:1021–1032
Kenzo T, Ichie T, Watanabe Y, Yoneda R, Ninomiya I, Koike T (2006) Changes in photosynthesis and leaf characteristics with tree height in five dipterocarp species in a tropical rain forest. Tree Physiol 26:865–873
King DA (1999) Juvenile foliage and the scaling of tree proportions, with emphasis on Eucalyptus. Ecology 80:1944–1954
Koch GW, Sillett SC, Jennings GM, Davis SD (2004) The limits to tree height. Nature 428:851–854
Laclau JP, Almeida JCR, Goncalves JLM, Saint-Andre L, Ventura M, Ranger J, Moreira RM, Nouvellon Y (2009) Influence of nitrogen and potassium fertilization on leaf lifespan and allocation of above-ground growth in Eucalyptus plantations. Tree Physiol 29:111–124
Leverenz J, Deans JD, Ford ED, Jarvis PG, Milne R, Whitehead D (1982) Systematic spatial variation of stomatal conductance in a Sitka spruce plantation. J Appl Ecol 19:835–851
Marshall JD, Monserud RA (2003) Foliage height influences specific leaf area of three conifer species. Can J Forest Res 33:164–170
McDowell N, Barnard H, Bond BJ, Hinckley T, Hubbard RM, Ishii H, Kostner B, Magnani F, Marshall JD, Meinzer FC, Phillips N, Ryan MG, Whitehead D (2002a) The relationship between tree height and leaf area:sapwood area ratio. Oecologia 132:12–20
McDowell NG, Phillips N, Lunch C, Bond BJ, Ryan MG (2002b) An investigation of hydraulic limitation and compensation in large, old Douglas-fir trees. Tree Physiol 22:763–774
Meinzer FC, Bond BJ, Karanian JA (2008) Biophysical constraints on leaf expansion in a tall conifer. Tree Physiol 28:197–206
Mullin LP, Sillett SC, Koch GW, Tu KP, Antoine ME (2009) Physiological consequences of height-related morphological variation in Sequoia sempervirens foliage. Tree Physiol 29:999–1010
Nobel PS (1977) Internal leaf area and cellular CO2 resistance—photosynthetic implications of variations with growth-conditions and plant species. Physiol Plant 40:137–144
Nobel PS, Zaragoza LJ, Smith WK (1975) Relation between mesophyll surface-area, photosynthetic rate, and illumination level during development for leaves of Plectranthus parviflorus Henckel. Plant Physiol 55:1067–1070
Nouvellon Y, Laclau JP, Epron D, Kinana A, Mabiala A, Roupsard O, Bonnefond JM, le Maire G, Marsden C, Bontemps JD, Saint-Andre L (2010) Within-stand and seasonal variations of specific leaf area in a clonal Eucalyptus plantation in the Republic of Congo. Forest Ecol Manag 259:1796–1807
Reich PB, Ellsworth DS, Walters MB, Vose JM, Gresham C, Volin JC, Bowman WD (1999) Generality of leaf trait relationships: a test across six biomes. Ecology 80:1955–1969
Rodrigo VHL, Stirling CM, Teklehaimanot Z, Nugawela A (2001) Intercropping with banana to improve fractional interception and radiation-use efficiency of immature rubber plantations. Field Crop Res 69:237–249
Ryan MG, Binkley D, Fownes JH, Giardina CP, Senock RS (2004) An experimental test of the causes of forest growth decline with stand age. Ecol Monogr 74:393–414
Ryan MG, Phillips N, Bond BJ (2006) The hydraulic limitation hypothesis revisited. Plant Cell Environ 29:367–381
Sack L, Melcher PJ, Liu WH, Middleton E, Pardee T (2006) How strong is intracanopy leaf plasticity in temperate deciduous trees? Am J Bot 93:829–839
Scholander PF, Hammel HT, Bradstreet D, Hemmingsen EA (1965) Sap pressure in vascular plants. Science 148:339–346
Smith WK, Nobel PS (1978) Influence of irradiation, soil-water potential, and leaf temperature on leaf morphology of a desert broadleaf, Encelia farinosa Gray (Compositae). Am J Bot 65:429–432
Stape JL, Binkley D, Ryan MG, Fonseca S, Loos RA, Takahashi EN, Silva CR, Silva SR, Hakamada RE, Ferreira JMA, Lima AMN, Gava JL, Leite FP, Andrade HB, Alves JM, Silva GGC, Azevedo MR (2010) The Brazil Eucalyptus Potential Productivity Project: influence of water, nutrients and stand uniformity on wood production. Forest Ecol Manag 259:1684–1694
Thomas SC, Winner WE (2002) Photosynthetic differences between saplings and adult trees: an integration of field results by meta-analysis (vol 22, p 117, 2002). Tree Physiol 22:817–817
Thomas D, Henson M, Joe B, Boyton S, Dickson R (2009) Review of growth and wood quality of plantation-grown Eucalyptus dunnii Maiden. Austral For 72:3–11
Turnbull JW (1999) Eucalypt plantations. New Forest 17:37–52
Walters GA (1973) Growth of Saligna eucalyptus: a spacing study after 10 years. J Forest 71:346–348
Woodruff DR, Bond BJ, Meinzer FC (2004) Does turgor limit growth in tall trees? Plant Cell Environ 27:229–236
Yoder BJ, Ryan MG, Waring RH, Schoettle AW, Kaufmann MR (1994) Evidence of reduced photosynthetic rates in old trees. Forest Sci 40:513–527
Acknowledgments
We thank the more than 100 people involved in Brazil Eucalyptus Potential Productivity Study (especially the project leader Jose Luiz Stape) and the companies that funded the work. Randy Senock, Christian Giardina, Holly Barnard, and James Fownes were particularly important in the Hawaii project, which was supported by National Science Foundation grants DEB93-06356 and DEB97-0852. MG Ryan was supported by a CSIRO McMaster’s Fellowship during the preparation of this manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Penuelas.
Rights and permissions
About this article
Cite this article
Coble, A.P., Autio, A., Cavaleri, M.A. et al. Converging patterns of vertical variability in leaf morphology and nitrogen across seven Eucalyptus plantations in Brazil and Hawaii, USA. Trees 28, 1–15 (2014). https://doi.org/10.1007/s00468-013-0925-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-013-0925-6