
Abstract
A study was conducted on the variation in growth, biomass, juvenile wood anatomy, and needle morphology of Pinus halepensis Mill. from three Spanish regions of provenance characterized by environmental differences, without the influence of the site factor. Seeds collected from two progenies in each region were planted in a single plot, and the trees were felled at the age of 7 years. The results showed significant differences between provenances, as well as the genetic or environmental influence on the traits analyzed. Trees adapted to moderate summer drought conditions (Inland Catalonia region) are primarily characterized by higher average values for height, diameter, biomass, cell wall thickness, inter-tracheid wall strength, pit membrane diameter, torus diameter, bordered pit aperture diameter, and ray tracheid abundance in comparison with trees adapted to severe summer drought conditions (Southern region and Balearic Islands region). The greater structural requirements of trees from the Inland Catalonia region, subjected to higher weight and wind loads, resulted in thick cell walls. Moreover, the large pits and more abundant ray tracheids in trees from this provenance would allow more efficient water transport and greater water storage capacity, respectively. The differences found between provenances suggest the adaptive nature of the anatomy of this species, which demonstrates the importance of the region of provenance when choosing reproduction material for reforestation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Within the genus Pinus in the Mediterranean region, the species Pinus halepensis Mill. occupies one of the largest areas, at around 3.5 million ha (Le Houerou 1980), particularly in the west. As it extends toward the east, the species is replaced by P. brutia in the region of the Aegean Sea. It is a typical Mediterranean species, capable of surviving severe summer drought and growing in poor, barren soil.
The Spanish P. halepensis forests occupy 1,770,530 ha (Maldonado et al. 1998), of which around 800,000 ha are natural forests, constituting almost 7% of the total forested area in Spain. The species is mainly found in the coastal regions of the Mediterranean (Catalonia, Autonomous Region of Valencia, Murcia, eastern Andalusia, and the Balearic Islands) and in the basin of the River Ebro as far as Álava, extending beyond the Mediterranean basin in the high valleys of the Guadalquivir, Guadiana, and Tajo rivers (Ortuño and Ceballos 1977). Although it is accepted that site water conditions are responsible for defining morphological and physiological attributes in P. halepensis seedlings (Royo et al. 2001), the seed source origin constitutes a determining factor in the end results. For instance, populations from dry areas of the western Mediterranean exhibited a conservative water-use, as opposed to fast growing sources from the northernmost distribution range (Voltas et al. 2008). Moreover, populations from resource-poor habitats had earlier reproduction but lower growth than populations from fertile environments (Climent et al. 2008).
In Spain, P. halepensis has a total of 18 regions of provenance (Gil et al. 1996), defined as territories subjected to practically uniform ecological conditions and consisting of populations which present analogous phenotypic or genotypic features. In order to ascertain the adaptive differentiation and evolutionary potential of the species, studies must be conducted in a single environment, given that there is little association between neutral genetic diversity and diversity assessed for quantitative or adaptive features in tree species, as seen in Pinus pinaster Ait. (González-Martínez et al. 2002).
In general, use of isozymes and terpenes has shown that P. halepensis is a species of very little genetic variability within the genus. Schiller et al. (1986) recognized two large groups: the eastern Mediterranean group, which implies introgression with P. brutia, and the western Mediterranean group. It has also been found that intrapopulation variability is much higher than interpopulation variability in nine populations throughout the Mediterranean area (Gómez et al. 2001).
Earlier provenance tests showing the response of different origins to the same environment revealed the existence of significant differences between P. halepensis provenances. For example, those from Morocco, Spain, and Algeria present lower growth and a poorer trunk form than the eastern provenances close to the distribution of P. brutia (Weinstein 1989). Significant genotype–environment interaction has also been noted in traits associated with adaptation to cold and drought, which suggests differential response by provenances to experimental sites (Gil et al. 1996). It is likely that this variation between provenances also affects the wood anatomical features, as seen in studies by other authors on different species (Aguilar-Rodríguez et al. 2006; Esteban et al. 2007).
The wood structure provides the tree with a number of functions essential for survival, including water, carbon and nutrient transport, mechanical support, and water and photosynthate storage (Ezquerra and Gil 2001; Rosner et al. 2007). First, adaptation of the wood to the environment must establish a balance between optimizing water transport and reducing the risk of embolism and hydraulic conductivity loss. Second, investment in aerial biomass requires mechanical support of the crown (compression), and growth in height increases the bending moment of the trunk, which must be resisted under dynamic wind load (Spatz and Bruechert 2000). Therefore, the wood structure will be determined by compromises between hydraulic and mechanical functions, which may complicate the search for structure–function relations (Chave et al. 2009).
Although the wood anatomy of P. halepensis from the Spanish forests has been widely studied (Castellarnau 1883; Peraza 1964; Esteban and Guindeo 1988; de Palacios 1997; Esteban et al. 2002), such works have nonetheless been limited to anatomical descriptions, with no analysis of variation according to regions of provenance. It is therefore hypothesized that the populations of P. halepensis growing in regions with different environmental conditions have developed specific morphological and anatomical adaptations which explain the differentiation of the species in its regions of provenance. The objective of this study is to check this hypothesis by studying variation and relations between tree growth and biomass, juvenile wood anatomy, and needle morphology of P. halepensis trees from three Spanish regions of provenance grown in a single plot in order to avoid the effect of the environment.
Materials and methods
Plant material
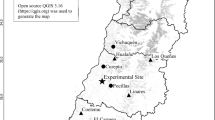
The material used for this study was collected in three natural forest of P. halepensis in the Spanish Mediterranean. These forests, part of the following regions of provenance: 3 (Inland Catalonia), 17 (Southern Spain), and 18 (Balearic Islands) (Table 1), were chosen because of their contrasting climates. The locations of these three regions are shown in Fig. 1, in addition to the distribution of average temperatures and rainfall throughout the year. The Inland Catalonia region, characterized by two maximum rainfalls (in spring and autumn) separated by 75 days of summer drought and with an average annual rainfall of 539 mm, is the most favorable region for plant activity. Winters in this region are cool (4–7°C) and summers are warm (<24°C). The Southern region has a maximum rainfall in December followed by abundant rain from January to March, and a marked summer drought lasting 130 days, from May to September. Annual rainfall is 584 mm. Winters are mild (>10°C) and summers are warm (>25°C). The Balearic Islands region is the driest of the three areas due to limited spring rainfall. It has maximum rainfall in October and a summer drought lasting 120 days, from May to August. Annual rainfall is 451 mm. Winters are mild (>10°C) and summers are warm (24–25°C) (Gil et al. 1996). The three populations are characterized by their occupation of predominantly limestone soil with mid-high permeability.

Location and climograms of the three Pinus halepensis regions of provenance included in this study (3. Inland Catalonia; 17. Southern Spain; 18. Balearic Islands)
Seeds were collected from two different families or progenies for each region of provenance, i.e., from two different trees. In 1998, after a year in a nursery, the plants were transferred to a single experimental plot at the National Agricultural Institute (INIA), in Madrid, as part of a provenance–progeny test consisting of 22 Spanish provenances. The plantation stake measured 1 × 0.6 m and the soil was of a sandy loam texture. The plot was not artificially irrigated during the test. The average annual rainfall for the duration of the test was 502 mm and the average annual temperature was 14.2°C, with a 4-month summer drought from June to September. At the age of 7 years, 17 trees from the Inner Catalonia region were felled, 11 from the Southern region and 15 from the Balearic Islands region, making a total of 43 trees (Table 1). From these trees a basal disk taken at a height of 3–5 cm from soil level was used to study the xylem anatomy and radial increment.
As the seeds were sown in a single plot, the variability of the species could be studied without the influence of other ecological factors (soil, temperature, humidity, exposure, etc.).
Field performance
Survival percentage, height, diameter, aerial biomass (stems, branches, and needles), and number of growth cycles (number and frequency of a second or third flush in 1 year) of the trees were measured in the last year of the trial.
Anatomical and radial growth measurements
Microscopic slides were prepared following the usual methods of softening, cutting, staining, and mounting. The wood anatomy was described in accordance with the recommendations of the IAWA Committee (2004). The samples were observed by means of light microscopy, both without staining and after staining with safranine and Sudan 4 in order to make the resin red (Jane 1970). With the light microscopy slides and the WinCell image analysis program, the following measurements were taken: axial tracheids [length, tangential lumen diameter (D) and cell wall thickness]; bordered pits [membrane diameter (D m), torus diameter (D t) and aperture diameter (D a)]; axial and radial resin canals (diameter); medullary rays (number per mm2, number of ray parenchyma cells per mm2 and number of ray tracheid cells per mm2); and cross-field pits (number per cross-field). The axial and radial resin canal diameters were measured in accordance with Method B in IAWA (2004), which measures the tangential diameter of the entire resin canal complex at the widest point. The axial tracheid measurements were taken on the earlywood. Tracheid length was measured in accordance with Ladell’s (1959) indirect method, by which two parallel lines are drawn on the tangential section at a known distance, perpendicular to the longitudinal axis of the tracheids. This is used to determine the number of cell tips contained between the lines in a predetermined number of adjacent cells, establishing the following relation: average tracheid length = (no. of cells × distance between lines)/no. of cell tips. Following Hacke et al. (2001), the tangential lumen (b) and thickness (t) of the double wall of two adjacent tracheids were measured and the inter-tracheid wall strength (t 2/b 2) was calculated. Because t 2/b 2 varies considerably depending on the value of b, this value was calculated for b = D h, where D h = ∑D 5/∑D 4 (tracheid hydraulic diameter). The torus overlap [(D t − D a)/(D m − D a)] was also calculated, as well as the pit aperture percentage (100 × D a/D m).
The biometry was carried out on five preparations from each tree taken from a basal disk, from rings 5 and 6. On each preparation 35 random measurements were taken for each of the variables studied. The number of pits per cross-field was measured on the five preparations from each tree, on ten different rays. The ray frequency and ray cell abundance were measured on all the rays contained in 1 mm2 on the tangential section.
The radial tree growth was studied on the same basal disk by measuring the width of each growth ring along the northern radius of the disk.
Foliar measurements
In the last year of the trial, six needles per tree were harvested and the leaf mass per area was calculated as a sclerophyll index (Gratani and Bombelli 2000). The projected needle area was obtained with a scanner. Images were analyzed using Scion image for Windows (Scion Corporation, Frederick, MD, USA). The needle samples were subsequently dried at 40°C for 5 days to determine leaf dry mass per unit area (g/cm2).
Needle length was measured with a digital caliper. Cross-sections of the central portion of three needles per tree were obtained with a sliding microtome, stained with safranine-fast green and mounted following a standard method (Johansen 1940). Sections were observed under light microscope and digitalized. Using Scion Image software (Scion Corporation, Frederick, MD, USA), measurements were taken from the following tissues: epidermis plus hypodermis, mesophyll, sum of resin canal lumens, and endodermis plus stele.
Statistical analysis
The variables were analyzed using a generalized linear model (GLM) (McCullagh and Nelder 1989) approach to ANOVA with type III sum of squares, using the following model:
with Y being the response variable, μ general mean, R effect of the provenance region, P(R) effect of the progeny nested within each region, T(F) effect of the tree nested within each progeny, and ε the error term. For field performance traits the T(F) factor was not considered. Regions were compared through multiple range tests with Fisher’s least significant difference (LSD) intervals.
Results
Plant survival and growth
The three provenances showed a high survival percentage (Table 2), with the maximum value in the Inland Catalonia provenance, whose trees also showed higher values for height, trunk diameter, aerial biomass, branch weight, and number of cycles than those from the Southern and Balearic Islands provenances (Table 2). Plant survival stabilized after the age of 2 years and the height of the plants was greater in the Inland Catalonia provenance than in the Southern and Balearic Islands region throughout the experiment.
The effect of the provenance was significant (P < 0.01) for the variables of height, base diameter, aerial biomass, branch weight, and number of cycles (Table 3), while the progeny effect was only significant (P < 0.05) for the number of cycles.
Anatomical wood description
Cross-section
Yellowish wood, characteristic of sapwood given the age of the trees. Abrupt transition from earlywood to latewood (Fig. 2a.1). Tracheids with rectangular outline in latewood and hexagonal in earlywood. Axial parenchyma absent. Axial resin canals present, with thin-walled epithelial cells (Fig. 2a.2). In wounded wood traumatic resin canals appear in tangential rows on the growth ring boundary (Fig. 2b.1), in addition to organic deposits in tracheids (Fig. 2b.2).

Pinushalepensis wood. a, bCross-section. a.1 Abrupt transition from earlywood to latewood. a.2 Axial resin canals present, with thin-walled epithelial cells. Scale bar 250 μm. b.1 Traumatic resin canals in tangential rows on the growth ring boundary. b.2 Organic deposits in tracheids. Scale bar 250 μm. cTangentialsection. Fusiform rays with resin canals. Scale bar 150 μm. d–fRadialsectiond.1 Uniseriate bordered pitting in radial walls of earlywood. d.2 Organic deposits in tracheids in wounded wood. Scale bar 100 μm. e End walls of ray parenchyma cells nodular. Scale bar 50 μm. f.1 Horizontal walls of ray parenchyma cells pitted. f.2 Cross-field pitting pinoid. f.3 Ray tracheids present, with dentate walls. Scale bar 50 μm
Tangential section
Uniseriate rays. Fusiform rays are due to the presence of radial resin canals with thin-walled epithelial cells (Fig. 2c).
Radial section
Tracheid pitting in radial walls of earlywood predominantly uniseriate (Fig. 2d.1). Organic deposits in tracheids in wounded wood (Fig. 2d.2). End walls of ray parenchyma cells nodular (Fig. 2e). Horizontal walls of ray parenchyma cells pitted (Fig. 2f.1). Cross-field pitting pinoid (Fig. 2f.2). Ray tracheids present, with dentate walls (Fig. 2f.3).
Xylem biometry
The average values of the measurements taken for each region of provenance are shown in Table 2. Trees from the Inland Catalonia region showed higher values for the variables of tracheid diameter (including the wall), wall thickness, inter-tracheid wall strength, pit membrane diameter, torus diameter, aperture diameter, ray frequency, number of ray tracheids per mm2, earlywood radial increment, and total radial increment than the trees from the Southern region and the Balearic Islands region. In contrast, tracheids were shorter and the torus overlap was lower in the trees from the Inland Catalonia region than in Southern and Balearic Islands region trees. Southern region trees showed a larger tracheid lumen and resin canal diameter than trees from the other two regions. Trees from the Balearic Islands region showed a lower pit aperture percentage than Inland Catalonia and Southern region trees.
The effect of the provenance was significant (P < 0.05) for all the xylem variables except radial resin canal diameter and number of ray parenchyma cells per mm2 (Table 3). The effect of the progeny nested within the provenance was significant in 10 of the 19 variables studied. The effect of the tree factor nested within the progeny was significant in all the variables except earlywood radial increment and total radial increment.
The average values for tree height, tracheid length, cell wall thickness, tracheid lumen diameter, and pit aperture diameter for each of the progenies studied are shown in Fig. 3. These parameters were chosen because of their value in explaining the structural variability between provenances and progenies. A high correspondence was observed between tree height (Fig. 3a) and tracheid wall thickness (Fig. 3c) but not between tree height and tracheid length (Fig. 3b). Tree height and tracheid wall thickness showed a highly significant correlation (P < 0.0001; Fig. 4). Tracheid length (Fig. 3b) showed high values in progenies 1723 (Southern region) and 1825 (Balearic Islands region). Tracheid lumen diameter showed differences between progenies in the Inland Catalonia and the Southern region, with a maximum value in progeny 1723 from the Southern region (Fig. 3d). This parameter showed a significant correlation (r = 0.36; P < 0.01) with the axial resin canal diameter (data not included). A notable aspect of the pit aperture diameter (Fig. 3e) is seen in the higher values for the two progenies from the Inland Catalonia region in comparison with the other two regions.

Average values for each of the progenies studied: a tree height, b tracheid length, c tracheid wall thickness, d tracheid lumen diameter, e pit aperture diameter. Different letters indicate significant differences between progenies (P < 0.05)

Relation between tree height and average cell wall thickness in earlywood tracheids. Each point corresponds to one tree. The line is a linear regression and the error bars represent one standard error
Needle morphology and anatomy
Needle length, dry mass per area, and epidermis plus hypodermis area did not show differences between regions (Table 2). The endodermis plus stele area and the mesophyll area were significantly higher in the Southern region.
The effect of the provenance was significant (P < 0.05) for the variables of endodermis plus stele area, sum of resin canal lumens, and mesophyll area (Table 3). The progeny factor was significant for all the needle variables except length and dry mass per area. Finally, the tree factor was significant for the variables of sum of resin canal lumens and mesophyll area.
Discussion
This study reveals intraspecific genetic differences in biomass and morphological and anatomical features of P. halepensis, which suggests the adaptive nature of some of the variables studied.
The larger size and aerial biomass attained by the trees from the Inland Catalonia region correspond to the more favorable climate conditions in their region of origin, as they are considerably less affected by summer drought. The greater availability of water and nutrients for the Inland Catalonia region trees, which allows greater growth, must correspond to specific adaptations in the structure of the conducting tissue. The larger pit dimensions in the trees from this region might result in higher hydraulic conductivity in the xylem. In Norway spruce, Rosner et al. (2007) found that high-specific hydraulic conductivity (or water permeability) in the xylem was more closely related to large pit apertures than to large earlywood tracheid lumens (e.g., Tyree et al. 1994; Kolb and Sperry 1999) or long earlywood tracheids (e.g., Mencuccini et al. 1997; Spicer and Gartner 1998). In fact, it is estimated that the pits are responsible for 50% or more of the total xylem hydraulic resistance (Choat et al. 2007). The higher average value for torus overlap found in the Southern and Balearic Islands trees in comparison with those from Inland Catalonia is noteworthy. This circumstance would make the pits in the drier regions more resistant to cavitation caused by the movement of the margo from its sealing position. Moreover, a negative relation between yield and xylem cavitation resistance has been observed in poplar and willow clones (Cochard et al. 2007). It would be interesting to check this relation in P. halepensis, as the data of the present study suggest that trees with a lower yield are more adapted to situations of water stress.
The high value of the axial tracheid wall thickness in the trees from the Inland Catalonia region in comparison with those from the other two regions is a notable finding. The high explained variance percentage for the provenance factor for this parameter, in addition to the inter-tracheid wall strength (Table 3), also suggests that these features are adaptive. In conifer wood, the tracheids need to be strong enough to simultaneously support the negative pressure of the water column and hold up the tree. The strong correlation found between cell wall thickness and tree height (Fig. 4) shows the importance of cell wall thickness as an element of structural strength, as the taller the tree is the more it is subject to wind load bending (Ezquerra and Gil 2001). These results concur with earlier studies in which high values for the double cell wall (t) in relation to the tangential width (b) are associated with resistance to collapse due to bending stress (e.g., Young 1989) and implosion stress in the conduit (e.g., Sperry et al. 2006). However, the greater mechanical requirements of the trees from the Inland Catalonia region did not result in longer tracheids, in contrast to what has been suggested in other studies (Mencuccini et al. 1997; Ezquerra and Gil 2001; Jagels et al. 2003; Jagels and Visscher 2006).
The higher values for wall thickness and inter-tracheid wall strength in the trees from the Balearic Islands provenance in comparison with the values in the Southern region trees are more difficult to explain in terms of mechanical strength, given that the two provenances present comparable heights and aerial biomass. Taking into account the fact that the Balearic Islands region is dryer than the Southern region, the differences in cell wall thickness could be associated with greater resistance to conduit implosion due to major negative pressure from the sap flow in situations of water stress (Hacke et al. 2001). The more drought-tolerant a plant is, the more it is able to tolerate negative pressure without cavitating and the greater the internal load on the tracheid walls is.
The differences between provenances in terms of lumen and earlywood tracheid length could have been influenced by a number of elements. Given that cell expansion is caused by turgor pressure on the wall as a result of water entering the vacuole, earlywood tracheid lumen could be affected by plant adaptation to the precipitation regime in its region of provenance at the start of the growing season. In this way, the Southern region receives most rainfall at the end of winter and the beginning of spring (278 mm from January to April, in comparison with 170 mm in the Inland Catalonia region and 137 mm in the Balearic Islands region), and the trees from this region had larger earlywood lumens.
The longer tracheids of progenies 1723 and 1825 (Fig. 3b) would allow water to cross fewer end-wall crossings per unit of length, which could mean lower total hydraulic resistance. Furthermore, reference has been made to trees growing at high altitudes having shorter tracheids than trees at lower altitudes (Zobel and van Buijtenen 1989). However, in the present study this relation was not observed, as the tracheids in the highest altitude provenance (Southern region) showed similar lengths to those in the lowest altitude provenance (Balearic Islands region). Small tracheids have also been associated with resistance to cavitation caused by frost (Pittermann et al. 2006; Sperry et al. 2006), but in the regions of provenance of this study, frost is not a significant stress factor compared to marked summer drought.
In the case of Pinus pinaster Ait., provenances with lower rainfall show the lowest values for osmotic potential at the turgor loss point and full turgor, as well as the lowest modulus of elasticity (Fernández et al. 1999). In situations of water stress, the low negative pressure of the sap flow can cause major hydraulic conductivity loss due to rupture of the water column. Rupture is caused by air seeding in the xylem conduits through the pits (Sperry and Tyree 1990; Pittermann et al. 2006). When embolism occurs, the capillary suction of the sap flow aspirates the membrane and the torus seals the aperture, creating a barrier to air movement (Hacke et al. 2004). The resulting valve is effective up to a certain difference in pressure, above which air filters through and causes tracheid cavitation.
Because of their pit structure, trees from the Inland Catalonia provenance would probably be quicker to cavitate as a result of water stress. It has been hypothesized that membranes of thick-walled earlywood cells could be less flexible (Rosner et al. 2007), which means it would not be easy for them to move in order to seal the aperture completely, as proposed for compression wood and latewood (Domec and Gartner 2002; Mayr and Cochard 2003). The differences found in the pit aperture percentage suggest that the pits in the Balearic Islands provenance, the driest of the three regions studied, would seal the pit more easily, and therefore these pits would be the most resistant to cavitation caused by water stress. According to Mayr et al. (2002), hydraulic vulnerability is positively related to pit aperture percentage, as pits with a higher aperture percentage appear to be more susceptible to air seeding due to the movement of the margo from its sealing position.
In addition to the functions of mechanical support and water conduction, tracheids have the ability to store water, which is important in the case of P. halepensis as this species normally grows in dry regions where the soil has low water retention capacity (Gil et al. 1996). One particularly important aspect is the different ray structure between provenances. The higher proportion of ray tracheids in trees from the Inland Catalonia region would indicate greater radial water movement capacity and water storage in the sapwood. This capacity in the provenance with least water stress could indicate an advantage over other, competing species, because the pines would take water from the soil in order to store it, as seen in the case of Pinus canariensis Chr. Sm. ex DC. (Climent et al. 2002). In dryer environments where less water is available, trees would not be able to accumulate the same amount of water and would therefore have a lower proportion of ray tracheids. In contrast, the abundance of ray parenchyma was similar in the three provenances, which suggests that this is a genetic factor which does not admit environmental variation. This parameter is probably determined by the minimal energy cost of maintaining the plant during times of water stress.
Provenance was highly significant (P < 0.001) in both earlywood increment and total increment, while neither the progeny nested within the provenance nor the tree nested within the progeny were significant (Table 3). This indicates uniformity in the radial growth pattern among trees from a single region and suggests that these parameters are adaptive. An interesting feature is the greater latewood increment seen in the trees from the Balearic Islands region in comparison with the Southern region trees, which is possibly associated with the maximum rainfall in the Balearic Islands region in September and October (Fig. 1).
A larger resin canal diameter would indicate greater resin flow through the canals. In P. sylvestris a strong correlation has been described between the resin productivity of the tree and resin canal diameter (Chudnyi 1974). Although it is widely accepted that the primary functions of resin canals are to prevent insect infestation (Pickard 2008) and protect the tree from fungal infections (Woodward 1992), resin canal formation has been associated with several types of stress, such as high summer temperatures (Rigling et al. 2003) or damage from wounding or mechanical pressure (Fahn and Zamski 1970). A notable aspect of the present study is the presence of organic deposits in the tracheids as a response to wounding, although no effects of temperature or drought on the axial resin canal diameter were observed, as the trees from the Inland Catalonia region and the Southern region showed similar values (Table 2). Furthermore, the correlation observed between axial resin canal diameter and tracheid lumen diameter does not lend itself to a clear functional interpretation, which means that further study on this aspect is required.
As regards the needles, no significant differences were found between progenies in terms of length or dry mass per area (Table 3). The latter parameter has been widely used as a sclerophyll index (Paula and Pausas 2006), and the fact that the three provenances showed similar values could indicate either the conservative nature of the species or similar phenotypic plasticity. The endodermis plus stele area was the variable most affected by provenance. The transfusion tissue, located in the stele, functions as storage for water and reserve metabolites (Canny 1993) and the larger area of this tissue in the Southern region, in conjunction with the larger mesophyll area, could help to maintain high photosynthetic levels in trees capable of sustaining reduced aerial biomass, which must make use of limited periods favorable for growth in environments subjected to extended periods of water stress (López et al. 2008).
The adaptive traits of P. halepensis seen in this study show the importance of the region of provenance when choosing the seed source for repopulation in Mediterranean environments, as seen in the provenances of P. pinaster used for this purpose after fire (Gil et al. 2009). The genetic characteristics of reproduction material will determine, to a certain extent, the tree structure and physiology and consequently the survival of trees and their ability to compete in a given environment. The high survival rate of the three provenances demonstrates the great value of this species for reforestation in poor soils and dry Mediterranean environments. The findings suggest that certain parameters are adaptive, particularly tree growth, cell wall thickness, bordered pit diameter, and ray tracheid abundance. However, the parameters of torus overlap, radial resin canal diameter, and ray parenchyma abundance were not affected by the provenance and also showed high intrapopulation variability, suggesting that these are genetic factors which allow low environmental variation. Fewer genetic differentiations were found in foliar parameters than in wood anatomy, which indicates major environmental influence and could indicate similar phenotypic plasticity in the three provenances.
References
Aguilar-Rodríguez S, Terrazas T, López-Mata L (2006) Anatomical wood variation of Buddleja cordata (Buddlejaceae) along its natural range in Mexico. Trees Struct Funct 20:253–261
Canny MJ (1993) Transfusion tissue of pine needles as a site of retrieval of solutes from the transpiration stream. New Phytol 123:227–232
Castellarnau JM (1883) Estudio micrográfico del sistema leñoso de las coníferas y en general del Género Pinus. Anales de la Sociedad Española de Historia Natural XII:136–219
Chave J, Coomes D, Jansen S, Lewis SL, Swenson NG, Zanne AE (2009) Towards a worldwide wood economics spectrum. Ecol Lett 12:351–366
Choat B, Cobb AR, Jansen S (2007) Structure and function of bordered pits: new discoveries and impacts on whole-plant hydraulic function. New Phytol 177:608–626
Chudnyi AV (1974) The importance of investigating the resin-duct system in Scots pine wood in selection for resin productivity. Genetika selektsiya, semenovodstvo i introduktsiya lesnykh porod, Moscow
Climent J, Chambel MR, Perez E, Gil L, Pardos J (2002) Relationship between heartwood radius and early radial growth, tree age, and climate in Pinus canariensis. Can J For Res 32:103–111
Climent J, Prada MA, Calama R, Chambel MR, De Ron DS, Alia R (2008) To grow or to seed: Ecotypic variation in reproductive allocation and cone production by young female Aleppo pine (Pinus halepensis, Pinaceae). Am J Bot 95:833–842
Cochard H, Casèlla E, Mencuccini M (2007) Xylem vulnerability to cavitation varies among poplar and willow clones and correlates with yield. Tree Physiol 27:1761–1767
De Palacios P (1997) Anatomía de maderas de coníferas a nivel de especie. Región norteamericana y europea, PhD thesis. Universidad Politécnica de Madrid, Escuela Técnica Superior de Ingenieros de Montes, Madrid
Domec JC, Gartner BL (2002) How do water transport and water storage differ in coniferous earlywood and latewood? J Exp Bot 53:2369–2379
Esteban LG, Guindeo A (1988) Anatomía e identificación de maderas de coníferas españolas. AITIM, Madrid
Esteban LG, de Palacios P, Guindeo A, García L, Lázaro I, González L, Rodríguez Y, García F, Bobadilla I, Camacho A (2002) Anatomy and identification of conifers wood as a species. Fundación Conde del Valle de Salazar-Mundi-Prensa, Madrid
Esteban LG, de Palacios P, Guindeo A, García Fernández F (2007) Comparative anatomy of the wood of Abies pinsapo and its two Moroccan varieties. IAWA J 28(3):285–299
Ezquerra FJ, Gil L (2001) Wood anatomy and stress distribution in the stem of Pinus pinaster Ait. Invest Agrar Sist Recur For 10:165–177
Fahn A, Zamski E (1970) The influence of pressure, wind, wounding and growth substances on the rate of resin duct formation in Pinus halepensis wood. Israel J Bot 19:429–446
Fernández M, Gil L, Pardos JA (1999) Response of Pinus pinaster Ait. provenances at early age to water supply. I. Water relation parameters. Ann For Sci 56:179–187
Gil L, Díaz-Fernández PM, Jiménez MP, Roldán M, Alia R, Agúndez D, de Miguel J, de Tuero M (1996) Las Regiones de Procedencia de Pinus halepensis Mill. en España. Organismo Autónomo Parques Nacionales, Madrid
Gil L, López R, García-Mateos A, González-Doncel I (2009) Seed provenance and fire-related reproductive traits of Pinus pinaster Ait. in Central Spain. Int J Wildland Fire (in press)
Gómez A, Alia R, Bueno MA (2001) Genetic diversity of Pinus halepensis Mill. populations detected by RAPD loci. Ann For Sci 58:869–875
González-Martínez SC, Gerber S, Cervera MT, Martínez-Zapater JM, Gil L, Alía R (2002) Seed gene flow and fine-scale structure in a Mediterranean pine (Pinus pinaster Ait.) using nuclear microsatellite markers. Theor Appl Genet 104:1290–1297
Gratani L, Bombelli A (2000) Correlation between leaf age and other leaf traits in three Mediterranean maquis shrub species: Quercus ilex, Phillyrea latifolia and Cistus incanus. Environ Exp Bot 43:141–153
Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloh K (2001) Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126:457–461
Hacke UG, Sperry JS, Pittermann J (2004) Analysis of circular bordered pit function. II. Gymnosperm tracheids with torus-margo pit membranes. Am J Bot 9:386–400
IAWA Committee (2004) IAWA list of microscopic features for softwood identification. IAWA J 25(1):1–70
Jagels R, Visscher GE (2006) A synchronous increase in hydraulic conductive capacity and mechanical support in conifers with relatively uniform xylem structure. Am J Bot 93:179–187
Jagels R, Visscher GE, Lucas J, Goodell B (2003) Paleo-adaptive properties of the xylem of Metasequoia: mechanical/hydraulic compromises. Ann Bot Lond 92:79–88
Jane FW (1970) The structure of wood, 2nd edn. A & C Black, London
Johansen DA (1940) Plant microtechnique. McGraw-Hill, New York
Kolb KJ, Sperry JS (1999) Differences in drought adaptation between subspecies of sagebrush (Artemisia tridentata). Ecology 80:2373–2384
Ladell JT (1959) A new method of measuring tracheid length. Forestry 32:124–125
Le Houerou N (1980) L’impact de l’homme et de ses animaux sur la forêt méditerranéenne. Forêts méditerranéennes II(I):31–34
López R, Climent J, Gil L (2008) From desert to cloud forest: the non-trivial phenotypic variation of Canary Island pine needles. Trees Struct Funct 22:843–849
Maldonado F, Sainz Ollero H, Sánchez de Dios R (1998) Distribución y estado de los bosques en España. UAM-WWF, Madrid
Mayr S, Cochard H (2003) A new method for vulnerability analysis of small xylem areas reveals that compression wood of Norway spruce has lower hydraulic safety than opposite wood. Plant Cell Environ 26:1365–1371
Mayr S, Wolfschwenger M, Bauer H (2002) Winter-drought induced embolism in Norway spruce (Picea abies) at the Alpine timberline. Physiol Plantarum 115:74–80
McCullagh P, Nelder JA (1989) Generalized linear models. Chapmann and Hall, London
Mencuccini M, Grace J, Fioravanti M (1997) Biomechanical and hydraulic determinants of tree structure in Scots pine: anatomical characteristics. Tree Physiol 17:105–113
Ortuño F, Ceballos A (1977) Los Bosques Españoles. INCAFO, Madrid
Paula S, Pausas JG (2006) Leaf traits and resprouting ability in the Mediterranean basin. Funct Ecol 20:941–947
Peraza C (1964) Estudio de las Maderas de Coníferas Españolas y de la Zona Norte de Marruecos. IFIE, Madrid
Pickard W (2008) Laticifers and secretory ducts: two other tube systems in plants. New Phytol 177:877–888
Pittermann J, Sperry JS, Hacke UG, Wheeler JK, Sikkema EH (2006) Inter-tracheid pitting and the hydraulic efficiency of conifer wood: the role of tracheid allometry and cavitation protection. Am J Bot 93:1265–1273
Rigling A, Brühlhart H, Bräker OU, Foster T, Schweingruber FH (2003) Effects of irrigation on diameter growth and vertical resin duct production in Pinus sylvestris L. on dry sites in the central Alps, Switzerland. For Ecol Manage 175:285–296
Rosner S, Klein A, Müller U, Karlsson B (2007) Hydraulic and mechanical properties of young Norway spruce clones related to growth and wood structure. Tree Physiol 27:1165–1178
Royo A, Gil L, Pardos JA (2001) Effect of water stress conditioning on morphology, physiology and field performance of Pinus halepensis Mill. seedlings. New For 21:127–140
Schiller G, Gonkle MT, Grunwald C (1986) Local differentiation among Mediterranean populations of Aleppo pine in their isoenzymes. Silvae Genet 35(1):11–19
Spatz HC, Bruechert F (2000) Basic biomechanics of self-supporting plants: wind loads and gravitational loads on a Norway spruce tree. For Ecol Manage 135:33–44
Sperry JS, Tyree MT (1990) Water-stress-induced xylem embolism in three species of conifers. Plant Cell Environ 13:427–436
Sperry JS, Hacke UG, Pittermann J (2006) Size and function in conifer tracheids and angiosperm vessels. Am J Bot 93:1490–1500
Spicer R, Gartner BL (1998) Hydraulic properties of Douglas-fir (Pseudotsuga menziesii) branches and branch halves with references to compression wood. Tree Physiol 18:777–784
Tyree MT, Davis SD, Cochard H (1994) Biophysical perspectives of xylem evolution: is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA J 15:335–360
Voltas J, Chambel M, Prada M, Ferrio J (2008) Climate-related variability in carbon and oxygen stable isotopes among populations of Aleppo pine grown in common-garden tests. Trees Struct Funct 22:759–769
Weinstein A (1989) Geographic variation and phenology of Pinus halepensis, P. brutia and P. eldarica. Israel. For Ecol Manage 26:215–225
Woodward S (1992) Responses of gymnosperm bark tissues to fungal infections. In: Blanchette A, Biggs R (eds) Defense mechanisms of woody plants against fungi. Springer, Berlin, pp 62–75
Young WC (1989) Roark’s formulas for stress and strain. McGraw Hill, New York
Zobel BJ, van Buijtenen JP (1989) The effect of provenance variation and exotic plantations on wood properties. In: Timell TE (ed) Wood variation. Springer, Berlin, pp 33–71
Acknowledgments
The authors are grateful to the following for their assistance: Prof. Luis Gil (Universidad Politécnica de Madrid), for reviewing the manuscript, making suggestions and providing a critique; Beatriz Becerril (Universidad Politécnica de Madrid), for assistance in preparing the microscopic slides; and Dr. José María Climent (CIFOR-INIA, Madrid), for providing the samples which made this study possible.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H. Cochard.
L. G. Esteban and J. A. Martín contributed equally to this work.
Rights and permissions
About this article
Cite this article
Esteban, L.G., Martín, J.A., de Palacios, P. et al. Adaptive anatomy of Pinus halepensis trees from different Mediterranean environments in Spain. Trees 24, 19–30 (2010). https://doi.org/10.1007/s00468-009-0375-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-009-0375-3