
Abstract
Carbon allocation within a plant depends on complex rules linking source organs (mainly shoots) and sink organs (mainly roots and fruits). The complexity of these rules comes from both regulations and interactions between various plant processes involving carbon. This paper presents these regulations and interactions, and analyses how agricultural management can influence them. Ecophysiological models of carbon production and allocation are good tools for such analyses. The fundamental bases of these models are first presented, focusing on their underlying processes and concepts. Different approaches are used for modelling carbon economy. They are classified as empirical, teleonomic, driven by source–sink relationships, or based on transport and chemical/biochemical conversion concepts. These four approaches are presented with a particular emphasis on the regulations and interactions between organs and between processes. The role of plant architecture in carbon partitioning is also discussed and the interest of coupling plant architecture models with carbon allocation models is highlighted. As an illustration of carbon allocation models, a model developed for peach trees, describing carbon transfer within the plant, and based on source–sink and Münch transport theory is presented and used for analyzing the link between roots, shoots and reproductive compartments. On this basis, the consequences of fruit load or plant pruning on fruit and vegetative growth can be evaluated.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plants invest in competing activities for carbon such as growth, reproduction or maintenance costs. In wild plants, allocation to reproduction and above and below ground vegetative parts is particularly important because it is closely related to plant dissemination and survival and hence fitness. Several theories have been proposed to explain the relative investment in the reproductive compartment, and between above and below ground vegetative parts. An important framework for explaining patterns of plant carbon allocation is the theory of allometry (West et al. 1997) which predicts intraspecific and interspecific scaling relationships among leaves, roots and reproductive biomass (Enquist 2002; Niklas and Enquist 2003). This theory, validated for wild plants, clearly shows that allometric rules reflect how metabolic production and biomass are partitioned between different plant parts. In agricultural crops, plants have been manipulated by humans for a long time through plant breeding and agricultural practices. Although it may at first appear that the great increase in productivity in the last century has been achieved through an enhanced efficiency of the photosynthetic apparatus, this does not seem to be the case (Ericsson et al. 1996). Indeed, the large gain in productivity among agricultural species is basically a result of a change in the carbon allocation pattern within the plant. In the case of fruit trees, increasing the number and size of fruits has been one of the main objectives of plant breeding. Consequently, modern cultivars have to invest an increased amount of carbohydrates produced via photosynthesis into fruit production. Fruit competition for carbohydrates can subsequently lead to a reduced vegetative growth of shoots and roots. This is typically the case of coffee where branch growth and development are strongly reduced on heavy fruit-bearing trees often leading to branch dieback and resulting in a strong alternate bearing pattern (Cannell 1971; Vaast et al. 2005). In order to cope with this kind of imbalance, agronomic practices seek to balance fruit and vegetative growth, either by manual or chemical fruit thinning or by increasing the vegetative growth through nitrogen supply, irrigation or pruning. Additionally, irrigation and fertilization may compensate the otherwise reduced growth of the root system. Plant architecture is also an important feature for management as it influences fruit size heterogeneity within the plant. Indeed, plant architecture is a network that connects the organs to each other and provides support for carbon transfers within the plant. Architecture also determines the spatial arrangement of organs and hence their activity (photosynthesis, respiration) mediated by energy exchange with their surrounding environment.
Carbon allocation within a plant depends on complex rules linking carbon source organs (mainly leaves) and carbon sink organs (mainly the sapwood of stems, branches and roots, and fruits). The complexity comes essentially from regulations due to feedback mechanisms, interactions between different functions and the spatial distribution of the different plant compartments. In order to analyse this complexity, carbon models of plant growth have been developed during the last 30 years (Le Roux et al. 2001). These models are powerful tools for analyzing how source and sink number, size and position within the plant affect carbon partitioning and, hence, vegetative and reproductive growth.
The first objective of this review is to address the conceptual framework of carbon allocation in plants, based on the current theories. The second objective is to exemplify the control of carbon partitioning through regulatory mechanisms and the interactions between processes in the plant system. The third objective is to analyse the role of plant architecture on carbon allocation. Finally, to illustrate the interest of carbon partitioning modelling, plant models are used to assess the effect of management practices such as fruit thinning and plant pruning on vegetative growth and fruit production.
Theories of carbon allocation
As first stated by Thornley and Johnson (1990) and subsequently confirmed by Lacointe (2000) and Le Roux et al. (2001), four main approaches have been used in models of carbon economy. These approaches are either empirical and allometric, teleonomic, driven by source–sink relationships, or based on transport and chemical/biochemical conversion concepts.
The models of carbon balance based on empirical allocation coefficients give usually reasonable predictions in the range of conditions for which these coefficients have been measured. These coefficients can be either constant or variable during the season as well as altered by external conditions by means of empirical relationships. An alternative to these allocation coefficients is to derive the biomass allocation between plant parts from the assumption that allometric relationships exist between different parts of the plant [Y = α X β where X and Y are masses (g) of two plant parts]. Causton and Venus (1981) presented the mathematical nature of allometry. The most interesting feature is that β is equal to the ratio of the relative growth rate of X and Y. Thus β can be regarded as the ratio of sink activities and can be very useful in models of carbon allocation. West et al. (1997, 1999) proposed a general model for the origin of allometric laws in biology whereby the evolution through natural selection resulted in optimal fractal-like vascular networks. These networks minimize the total hydrodynamic resistance and yet maximize the whole organism resource use by maximizing the scaling of surfaces where resources are exchanged with the environment. It has been shown, using this theory, that the mass and growth rate of one plant part can be expressed in terms of mass and growth rate of another part. One example is given for mango fruit for which a strong allometric relationship between stone dry weight and fruit dry weight has been established (Léchaudel et al. 2002). Derivation of such an allometric equation has been used by Lescourret and Génard (2005) to partition carbon between flesh and stone in a model of virtual peach fruit. At the level of the whole tree, King (2005) used an allometrically explicit model, which partitioned the biomass among the tree parts so as to maintain the specified allometry. He explored the effect of stem allometry on tree growth and found a strong influence of β. Allometric relationships, especially those based on trunk cross sectional areas, have been shown to be useful tools for optimizing the timing and intensity of management practices such as pruning (Droppelmann and Berliner 2000) and fruit thinning (Reginato et al. 2007). However, such models cannot represent either the regulations between processes, or the effect of management practices on carbon allocation coefficients. Moreover, the theory of allometric scaling is now a matter of debate (Muller-Landau et al. 2006).
Many models of plant carbon allocation are based on teleonomic approaches where some intrinsic goal is assumed, e.g., a functional balance between shoot and root (Davidson 1969) or a specific relationship between foliage and conducting tissue as in the pipe-model (Shinozaki et al. 1964). The pipe model theory states that the sapwood area (A, m2) at a given height and the foliage biomass above (W, g) are related through a constant ratio termed the proportionality constant (k, g m−2) (Shinozaki et al. 1964):
This can be interpreted, in terms of the functional balance theory, as a condition for an efficient water supply to foliage biomass. The proportionality constant is often assigned different values at different plant heights (Lacointe 2000). The pipe model is one of the most commonly used models to distribute resources between foliage and woody structure in process-based models (Sievänen et al. 2000). However, this model is probably inadequate to predict plant response to disturbances such as pruning or thinning (Le Roux et al. 2001).
Another important teleonomic approach concerns the root:shoot functional balance theory. Models of carbon allocation between shoots and roots have been based largely on carbon supplied by shoots and nitrogen supplied by roots (Charles-Edwards 1976; Reynolds and Thornley 1982). This approach has been extended to water supplied by roots (Chen and Reynolds 1997). According to the root:shoot functional balance theory, total nitrogen (or water) acquisition by the root system is proportional to total carbon assimilation by shoots. This functional balance theory was described by Davidson (1969) as:
where W s and W r are shoot and root biomasses (g), respectively, S s and S r shoot and root specific activities (day−1, h−1 or s−1, according to the time step), respectively, and k constant. The specific activities are usually allowed to vary with soil N or water availability (for S r) and with above-ground environmental resources such as light intensity and atmospheric CO2 concentration (for S s). In agreement with the root:shoot functional balance theory, Grechi et al. (2007) recently found a strong allometric relationship between grapevine roots and shoots. This relationship was highly dependent on nitrogen and light supply, so that a strong relationship between the root:shoot ratio and the C:N ratio could be established.
Many models utilize optimization principles whereby the relative root:shoot allocation is considered in the context of maximizing the relative growth rate of the plant or by constraining the C:N ratio to a target value (Thornley and Johnson 1990; Lacointe 2000; Le Roux et al. 2001). However such optimization must rely on major simplifications in order to facilitate analytical solutions toward the optimal control strategy and assumes that plants “anticipate” the environmental conditions (Reynolds and Chen 1996). As an alternative to optimisation, Reynolds and Chen (1996) and Chen and Reynolds (1997) proposed a “coordination theory” where the allocation coefficients are driven by the imbalance between either root N or water supply and shoot carbon supply. Such a coordination has been also proposed for young peach trees by Génard et al. (1998). In this case, a variable RS reflecting the balance between the mass of roots younger than 1 year (W r, g) and the leafy shoot mass (W s, g) is defined as:
The parameter RS e is equal to the ratio of the weight of young roots and the weight of shoots when the tree is at equilibrium. When RS is greater than one, there is an imbalance in favour of roots and assimilates are preferentially allocated to shoots, whereas when RS is less than one, assimilates are preferentially allocated to roots. The consequence is a fluctuation of the root:shoot ratio along the season. Equilibrium is reached when RS is equal to one.
In the models based on source–sink relationships, carbon allocation is assumed to depend on the respective ability of the different sinks to import available assimilates from the same sources (Lacointe 2000). According to Grossman and DeJong (1994), this ability, “sink strength” or “sink demand”, is based on the genetically determined potential growth respiration rate, maintenance respiration rate and net sink strength (g day−1). The potential net sink strength is the maximum rate at which the organ can accumulate dry matter per unit time. It is the product of sink size (g) and potential net sink activity (day−1) expressed as the relative growth rate. The potential net sink strength of an organ can be decreased by sub-optimal environmental conditions. That new sink strength is termed conditional net sink strength by Grossman and DeJong (1994). The sink growth rate, also called apparent net sink strength, is calculated from the conditional net sink strength, considering the resource availability.
Experimentally determined seasonal patterns of organ growth potential are frequently used in the literature to represent potential net sink strengths. Regarding fruits, Marcelis (1996) argued that series of growth rates, measured under conditions of non-limiting assimilate supply, provide a correct estimation of potential net sink strength, based on the idea that potential growth only changes with time. However, this idea implies that constrained fruit growth is able to reach the potential growth rate after removal of competing sinks. This capability has been observed for cucumbers (Marcelis 1993) or tomatoes, but not for a fruit tree species such as peach (Grossman and DeJong 1995). For such species, the conditional net sink strength at a given time depends on its state at this time, i.e., on the past growth. Moreover, the dynamics of fruit growth and particularly the time at which fruit growth decreases or stops, depends on the accumulation of matter in the fruit (Havis 1962; Proebsting and Mills 1981; Johnson 1995).
Lescourret et al. (1998) proposed the following equation for the conditional net sink strength (CSS, g day−1). This equation emphasizes the role of fruit “history” by means of the accumulated growth (W, g), both in terms of sink size and sink activity. It also emphasizes the role of time and temperature by means of accumulated degree-days (dd; Fig. 1).
where RGR ini (day−1) is initial relative growth rate, W max (g) refers to the limiting final potential weight and dd min and dd max (degree-days) are parameters.

Conditional net sink strength (CSS) of peach fruit for different initial masses. The fruit history (here the initial fruit mass) has strong effect on CSS. The parameters of the CSS equation were RGR ini = 0.011 day−1, W max = 37 g, dd min = 463 degree-days, dd max = 987 degree-days
Carbon supply, on the other hand, consists in the carbon produced by source organs with photosynthetic capacity, mainly leaves, and the carbon stored as reserves. Hence, source–sink models incorporate leaf photosynthesis models which differ in their complexity level in order to estimate carbon produced by leaves. Some models also incorporate fruit photosynthesis as a carbon source (Lescourret et al. 1998). In most source–sink models, carbon reserves accumulate when carbon supply exceeds total carbon demand. Alternatively, reserves are treated as competing sinks as is the case in the model for peach developed by Allen et al. (2005).
An important step after the quantification of organ demands and carbon availability in source organs is the carbon allocation from sources to sinks. Le Roux et al. (2001) summarized the allocation rules that are applied in most models. When demand is less than supply, each sink gets its own demand and excess supply goes to reserves. On the other hand, there are two approaches dealing with the case of supply shortage. In the “proportional” approach, the carbon supply by sources is shared by the sink organs that all get the same proportion of their demand. Alternatively, in the “hierarchical-priority” approach, the sink with the highest priority is served first, then the component with the next priority level is considered, and so on. The maintenance respiration requirements are assigned the highest priority because they are vital for organ survival. Most current models use a combination of these rules.
Models based on transport and chemical/biochemical conversion concepts open the way for a more mechanistic description of the carbon partitioning. They permit the avoidance of the use of empirical allocation coefficients, functional balance rules, or fixed allometric relationships. Thornley (1972, 1998) first proposed a transport-resistance model for shoot:root partitioning in relation to C and N availability. In this approach, C and N enter the plant through uptake processes, plant compartments are connected via transport pathways, and substrates are used for growth. Transport rate is proportional to concentration differences. This approach can account for the effect of N on carbon partitioning and shoot:root ratio. It is generally assumed that the mechanism governing the transport of assimilates is the Münch pressure-driven flow (Dale and Sutcliffe 1986). Using the simplified formulation of the Münch hypothesis described by Thornley and Johnson (1990), Minchin et al. (1993) presented a simple mechanistic model of phloem transport that explains sink priority as a function of sugar gradient. In the same theoretical framework, Bassow and Ford (1990) proposed a space–time model of carbon translocation in forest trees. More recently, Bruchou and Génard (1999) proposed a space–time model of carbon translocation along a shoot bearing fruits. The novelty of this model comes from the aggregation of physiological processes taking into account spatial aspects. The stem is represented as a set of compartments connected to source (leafy shoots) and sink (fruits) compartments. The physiological processes considered are photosynthesis, respiration of fruits and leaves, translocation of assimilates and fruit growth. Assimilate production is regulated by sink strength and light availability. Carbon translocation between two compartments depends on the gradient of assimilate concentration using the simplified formulation of the Münch hypothesis (Thornley and Johnson 1990). A similar approach has been used by Bidel et al. (2000) for modelling the root growth. These authors used their model to analyze how differences in meristem sink strength and/or phloem conductivity can control the morphology of the root system. Münch pressure-driven flow has been mathematically validated by Henton et al. (2002) by modelling the dynamics of long-distance solute transport inside a semi-permeable tube. Nevertheless, Bancal and Soltani (2002) state that, for a plant system, the use of resistances in the Münch modelling approach becomes a mathematical burden, not even easily quantifiable since they are related to anatomical traits that are difficult to assess. In the opinion of these authors, by leaving out resistances, it becomes easy to calculate sink activities directly from source activities, using an intuitive, accessible parameterization. Recently, Allen et al. (2005) presented a carbon allocation model within the whole tree architecture based on an electric analogy. Daudet et al. (2002) proposed a more complete theoretical model considering phloem and xylem flows in order to take into account carbon–water interactions.
An important step, concerning fruit trees, is the sugar unloading to fruits and the biochemical transformations of sugars within these fruits. The sugars can be transported from the phloem to the fruit by active transport, mass flow or diffusion. Fishman and Génard (1998) proposed the following equation to represent the total fruit uptake of carbohydrates (U, g):
where U a (g) is uptake rate due to active transport, U p (g) phloem flow of liquid entering the fruit, C p and C f sugar concentrations (g g FW−1) in the phloem and fruit, respectively, σ reflexion coefficient which is a measure of impermeability of the cell membrane to solutes, A membrane area (cm2) through which solutes diffuse and p s (g cm2 h−1) solute permeability coefficient. If σ = 1, the membranes are impermeable and there is no sugar uptake through mass flow. As p s is usually small, the diffusion component can often be neglected. The uptake rate (U a) obeys the Michaelis–Menten equation:
where v m (g sucrose g DW−1 h−1) is maximum uptake rate and KM (g g FW−1) Michaelis–Menten constant.
The transformation of phloem sugars (sucrose, sorbitol, ...) into sink soluble carbohydrates (sucrose, glucose, fructose, ...), starch or cell walls is one aspect of carbon partitioning that is usually ignored. It is a major process of growth because sink soluble carbohydrates drive the sink osmotic potential which in turn drives the sink water uptake. In the case of fruits, the transformation of phloem sugars into other sugars, starch and cell wall components determines their quality which is an important component of their market value. This is important not only for fruits produced for their flesh such as mango or peach but also for fruits produced for their seeds or beans such as coffee.
Génard and Souty (1996) and Génard et al. (2003) designed a mechanistic model, called SUGAR (Génard and Lescourret 2004) to predict the changes in sugar composition during fruit development (Fig. 2). The model was designed for peach, but the main principles can be used for other fruits. In this model, the unloaded sugars are either directly stored in the tissues or transformed into CO2 through the respiratory process or transformed into other sugars or used to synthesize other compounds (structural carbohydrates, etc.). The enzymatic reactions were described according to the “rate law” of chemical kinetics (Chang 2000), which states that the reaction rate is proportional to the reactant concentration. Thus, the rates of change in the carbon amounts of sugar compounds depend on sugars already accumulated in the fruit flesh. They are described through a set of differential equations, each equation being of the following form:
where C j (g) is carbon amount in sugar j, E j (g day−1) and R j (g day−1) which can be equal to zero, depending on the compartment, are, respectively, the carbon flow from the phloem and the carbon loss by respiration, k ij (day−1) is a function of parameters (θ) and variables (x) describing the relative rate of sugar transformation of sugar i into sugar j.

Diagram illustrating the carbon partitioning by the SUGAR model to predict the changes in sugar composition along the peach fruit development. Arrows and boxes represent carbon fluxes and carbon components, respectively. The two ellipses represent carbon supply and losses by respiration. The proportion of sucrose in the phloem-sourced sugar pool (F1) and the relative rates of sugar transformation F2–F6 for each arrow are indicated (from Génard and Lescourret 2004). The sucrose is splitted into glucose and fructose with same rate F2
Regulations and interactions
The carbon allocation within a plant and between different biochemical compounds results from strong regulation of source and sink strength. These regulations are at the basis of teleonomic approaches such as the functional equilibrium in which the sink strength of the shoot and root systems is regulated by the state of the equilibrium between them. These regulations are also considered in the source–sink models where the conditional net sink strength, and thus the carbon partitioning, is regulated by the history of the sink itself. Moreover, in some source–sink models, the source strength is regulated by the sink strength (Léchaudel et al. 2005) or by the amount of reserves in leaves (Lescourret et al. 1998). It is interesting to notice that the high genotypic differences observed in peach photosynthesis by Quilot et al. (2002) is not related to the variation of the potential photosynthesis that is very similar between genotypes, but to differences in fruit sink strength. Indeed, genotypes with low fruit sink strength accumulate reserves in leaves, which depresses the actual photosynthesis through a feedback mechanism. Such a feedback mechanism has also been documented in coffee plants with low fruit loads (Vaast et al. 2005; Franck et al. 2006).
In models based on the transport and chemical/biochemical conversion concepts, regulations are often emerging properties of the system. The model of Bruchou and Génard (1999) is used hereafter to exemplify this fact.
When the model was run for leaves:fruit ratios of 5 and 30, simulated photosynthesis was independent of the leaf:fruit ratio early in the morning, but after midday it decreased concomitantly with the ratio value (Fig. 3). Similarly, a decrease in photosynthesis with low fruit loads has been observed for different tree species (Gucci et al. 1995; Chalmers et al. 1975; BenMimoun et al. 1996; Franck et al. 2006; Iglesias et al. 2002; Syvertsen et al. 2003). The diurnal variation of leaf carbohydrate content (Fig. 3) followed a classical pattern, increasing during the day and decreasing at night (Upmeyer and Koller 1973; Sharkey and Pate 1976; Franck et al. 2006). The leaf carbohydrates content increased with the leaf:fruit ratio (LF) whereas the export of carbohydrates from leaves decreased from LF = 5 to LF = 30 (Fig. 3). The concentration of carbohydrates simulated in the woody shoot exhibited a diurnal periodicity and increased with increasing leaf:fruit ratio. The simulated unloading rate from phloem to fruit increased strongly with increasing leaf:fruit ratio. The diurnal variations in unloading rate closely followed the variations of the loading rate (Figs. 3, 4) showing that the system rapidly reacted to changes in assimilate supply.

Simulated diurnal variation of peach leaf photosynthesis and carbohydrate content, and phloem loading rate for leaves:fruit ratios equal to 5 and 30 (from Bruchou and Génard 1999)

Simulated diurnal variation of unloading rate from phloem to fruit according to leaf:fruit ratio in peach (from Bruchou and Génard 1999)
The consequence of these different unloading rates is an increase of fruit mass with increasing leaf:fruit ratio. These simulations showed that a change in source:sink ratio had a strong influence on the whole system. The sugars can be considered as a signal that operates as a regulator, i.e., an increase of leaf sugar content regulates the photosynthesis by feedback inhibition, or an increase in phloem sugar concentration induces an increase of sugar unloading in the fruit.
Carbon partitioning also results from the interactions between different processes. The transport resistance model of Thornley (1972, 1998), which considers the transport and chemical conversion of substrate C and N to structures, clearly shows that carbon allocation is highly dependent on N supply and allocation within the plant. Another example can be taken from the theoretical work of Daudet et al. (2002). These authors showed how carbon allocation can be affected by the water status of the plant. They showed that the fruit growth rate in terms of dry matter can be decreased by plant water stress despite the fact that the sucrose source remained unchanged. Interestingly, using the model of Fishman and Génard (1998), Lescourret et al. (2001) found that increasing skin surface conductance for water increased the fruit dry weight and decreased the fruit fresh weight. This feature is explained by the fact that when fruit surface conductance is increased, fruit transpiration also increases leading to a decreased fruit fresh weight. Concomitantly, this higher fruit transpiration increases the mass flow of sugar due to a higher water inflow to the fruit increasing its dry weight.
From these examples about regulation and interaction between carbon, nitrogen and water, it can be seen that mechanistic models are important tools for analyzing and understanding the carbon allocation within the plant.
Partitioning and architecture
During the last decade, much research has been initiated in order to understand the complex interactions between plant architecture and the physical and biological processes that drive plant growth and development. The impulse for this research was given by a new generation of so-called Functional Structural Plants Models (FSPM) allowing the mimicking of ecophysiological processes on virtual plants (Ford 1992; Bosc 2000; King 2005; Vos et al. 2007). Costes (2004) reviewed such models, and Sievänen et al. (2000) and Godin and Sinoquet (2005) discussed the different questions raised around these functional–structural plant models. Two points are important to consider regarding carbon allocation: (1) the ability of the models to account properly for the role of crown geometry in light capture that is the basis of assimilate production and (2) the way the plant architecture is used to distribute assimilates between sources and sinks.
Light capture and carbon assimilation
Fruit orchards and vineyards exhibit a wide variety of canopy shapes generally moulded by pruning and wiring. Purely geometrical models such as the ellipsoidal model of Charles-Edwards and Thornley (1973) represent tree crowns by geometrical shapes filled by foliage interacting with light as a turbid medium. These models, considering light interception at the scale of the whole crown, do not bring relevant information for addressing carbon partitioning at an infra-crown scale. Models splitting foliage into 3D cells (List and Küppers 1998; Röhrig et al. 1999; Sinoquet et al. 2001) give useful information about the spatial distribution of light interception but with only indirect association between light interception and plant topology. Alternatively, some FSPM light models simulate light interception at the scale of individual plant entities such as shoots (e.g., LIGNUM: Perttunen et al. 1996) or elementary organs, such as leaves, internodes or fruits (Yplant: Pearcy and Yang 1996; Vegestar: Adam et al. 2004; MMR: Dauzat and Eroy 1997; Nested Radiosity: Chelle and Andrieu 1998). Such models, where plant entities are connected to each other through a description of tree topology, are suitable for addressing carbon allocation between entities. Light models also differ in the way they account for the additional foliage irradiance resulting from multiple scattering within canopies that represents about 10–15% of the irradiance by incident light in the PAR. Radiosity is the more common method for simulating light exchanges between entities in a 3D space (Chelle and Andrieu 1998; Evers et al. 2005; Soler et al. 2003) but Ray-Tracing was recently applied to canopies (Allen et al. 2005) owing to the reduction of computation time allowed by the quasi-Monte Carlo method (Keller 1996).
Depending on the purpose, carbon assimilation modelling may integrate more or fewer features and details. For instance, in forest models working at yearly or seasonal time step, carbon assimilation may be reasonably restricted to semi-empirical relationships between PAR irradiance and photosynthesis (Perttunen et al. 1998; List and Küppers 1998). On the other hand, addressing specific questions such as the diurnal balance of assimilates production and consumption, requires a more comprehensive integration of biophysical processes with their interactions. As an example, a comprehensive model including the Farquhar’s assimilation model (Farquhar et al. 1980), the Ögren’s photoinhibition model (Ögren 1991) and the Ball–Berry–Leuning stomatal conductance model (Ball et al. 1987; Leuning 1995) was developed to simulate the carbon assimilation of coffee orchards grown under different irradiances and with variable fruit loads (Franck et al. 2006; Dauzat et al. 2006). The model allowed mapping the carbon assimilation throughout plant crowns according to their local physical environment (Fig. 5). An application was the analysis of factors responsible for the mid-day depression of carbon assimilation such as foliage temperature, photoinhibition or sugar accumulation in leaves. Results showed the fact that the actual photosynthesis may be far under the potential photosynthesis when the fruit:leaf ratio is low because of the negative effect of sugar accumulation. This leads to the challenging question of adjusting fruit loads that promote photosynthesis without being detrimental to vegetative growth. In this respect, FSPM are becoming useful tools for addressing emergent properties of complex systems.

Simulation of Coffea arabica photosynthesis at leaf scale by the Archimed model (Dauzat et al. 2006). Light interception and photosynthesis are calculated at the scale of individual leaves. Colors being ordered from blue to red, the redder the leaf, the higher the photosynthesis
Architecture and carbohydrates partitioning
Functional–structural models were first developed for forest trees (e.g., GROGRA: Kurth 1998; LIGNUM: Perttunen et al. 1998 or ALMIS: Eschenbach 2005) but recently a comprehensive model was developed for peach tree (L-Peach: Allen et al. 2005). In this model, plant organs are connected through a topological description allowing the simulation of carbohydrates fluxes between sink and source organs. The assumption is that fluxes are proportional to differences in carbohydrate concentration and inversely proportional to a resistance to transport. Carbon partitioning is an emergent property of such FSPM which made it possible to avoid empirical allocation coefficients, functional balance rules, or fixed allometric relationships (Allen et al. 2005). To illustrate this property and the way plant architecture is involved in carbon partitioning we will take the example of two simple systems: a branch bearing fruits and a root system.
The model of Bruchou and Génard (1999) has been used to analyse the competition between peach fruits located at different positions within a single branch. They studied a branch bearing two fruits and four leafy shoots, fruits and shoots being on the opposite extremities of the branch. The simulation predicted a decrease of 22% in dry matter for the fruit farthest from the leaves, which was consistent with experimental results. In order to analyse via the model, whether the differences in fruit growth resulted from a competition between fruits, the fruit situated between the leaves and the other fruit having an advantage, or from a pure distance effect, the cases where only the proximal or distal fruit was kept on the stem were simulated. In both situations, fruit growth was the same, which shows that, according to the model, there is no distance effect but a competition effect. This means that relative position of sinks and sources within the topology of the system were more important than the absolute distances between them.
A modelling framework has been recently developed to analyse carbon transfer within a complex root architecture (Vercambre, in preparation), generalizing approaches previously developed along a single axis (Thompson and Holbrook 2003; Hölttä et al. 2006). In this model, movement of the phloem sap is caused by a gradient in hydrostatic pressure, the pressure in the phloem being osmotically induced accordingly to the Münch’s theory. The sieve plate is assumed to be totally permeable to solutes, i.e., the reflection coefficient is nil. Only one form of carbohydrate is considered in the present case and other solutes are neglected. Furthermore, carbohydrate unloading is supposed to be limited by an enzymatic process. The model allows the simulation of the carbohydrates distribution in the root system (Fig. 6) and assesses the assimilate concentration throughout the root system according to its topology and root properties. Large concentration gradients appear along the tap-root axis, especially for lateral roots with limited phloem transfer capacities. The carbohydrate concentration gradient is relatively low for the main axis compared to lateral roots. Along the main axis, the presence of lateral roots leads to a large decrease in carbohydrate availability, due to carbohydrate unloading in lateral roots. The carbohydrate concentration is highly variable in the secondary root tips, depending on the location of the lateral root insertion on the main axis as well as on the length of this secondary root; the longer the root, the lower the carbohydrate concentration. Furthermore, a steep decrease in carbohydrate concentration is observed at the root tips. This decrease results from high sink activity of the root tips due to a high metabolic and growth activity.

Simulated phloem carbohydrate concentration throughout a tap root system 100-day old. Root collar carbohydrate concentration is fixed at 0.5 M and the xylem water potential is assumed to be uniform and nil. The darker the root, the smaller the concentration
These examples clearly show that we are now able to take into consideration the architecture–function interactions in models that can help us to analyse how the plant is functioning. The future challenges are certainly (1) to provide these models with experimental data on conductance, phloem concentrations, ..., etc., in order to test their underlying hypotheses and (2) to model the carbon partitioning in more complex systems such as the whole tree. Recent attempts on different species are promising (Allen et al. 2005; Eschenbach 2005). They allow the analysis of interesting emergent properties of the system such as different growth forms of trees resulting from simulations under various conditions.
Effect of cultural practices
Most agricultural practices are able to change carbon partitioning within the plant. There are numerous papers showing that high levels of nutrient supply increase shoot growth relatively to root growth (Cannell 1985). The root/shoot models predict that water stress would increase the root shoot ratio, which is actually what is observed in most cases (Cannell 1985; Ericsson et al. 1996). Effects of pruning and fruit thinning are the two other important practices having a strong influence on carbon partitioning. These effects will be analysed hereafter using structural–functional models.
Effect of pruning on 1-year old tree
Génard et al. (1998) studied the effect of pruning on carbon allocation, using a model simulating growth and development of peach trees during their first growing season. This model distinguishes four types of plant compartments: the main aerial axis, the secondary aerial axes, the rootstock and the new roots. Tree structure is described by the position of the secondary axes on the main axis. The processes considered by the model are: (1) the plastochron activity of the main axis, which defines the time needed to produce a new metamer and its related ramification; (2) the mobilization of rootstock reserves and carbon acquisition for tree growth; and (3) the partitioning and use of carbon for maintenance and growth. The balance between root and shoot growth is managed using the principles of “coordination theory”. If the shoot:root ratio is altered by pruning, it tends to be restored by compensatory growth through an alteration of the partitioning scheme. Growth correlations between the main and secondary axes are also considered in order to define the rules of assimilate partitioning. The model, validated by experimental data, accounted for several previously reported pruning responses. For example and as classically observed, the model predicts that pruning intensity and date of pruning have little effect on the seasonal carbon balance and consequently on the total dry weight at the end of the growing season (Webster and Shepherd 1984; Kikuchi et al. 1989; Neri et al. 1992). The model also predicts a growth increase of the axes remaining after pruning, tending to recreate a more functional root:shoot ratio which is well documented in the literature (Cannell 1985). Pruning also increased the rhythmic growth of shoot and root in the model simulations as formulated by Atkinson (1980). More recently, a structural–functional model simulating walnut tree growth in response to pruning has been proposed by Balandier et al. (2000) for several year old trees.
Effect of pruning and thinning on adult trees
In order to analyze the effects of management on trees bearing fruits, we are currently developing a “virtual tree” model describing carbon transfer within the plant between shoots bearing fruits, trunk and branches, and roots. This model is based on previous models developed for peach (Lescourret et al. 1998), coffee (Vaast et al. 2002) and mango (Léchaudel et al. 2005). It assumes that the plant is a set of shoots bearing fruits connected to each other by the branches. Carbon allocation between shoots bearing fruits and with the other plant parts is based on source–sink concepts as well as on a simplified version of the Münch transport theory. The leafy shoots have the first priority for growth. The balance between root and shoot growth is managed using the principles of “coordination theory”. The physiological processes considered are leaf and fruit photosynthesis, respiration of all the plant organs, carbon storage and remobilisation in leaves, branches, trunk and roots, and the growth of organs. Leaf photosynthesis is regulated by its reserve concentration.
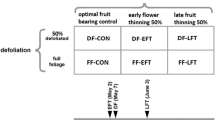
Simulations were performed on a peach tree with two main branches and 20 shoots bearing fruits. The effect of summer pruning and fruit thinning was analysed considering an unpruned tree with high fruit load, an unpruned and thinned tree (80% of fruits removed), a pruned (50% of leafy shoots removed) tree with high fruit load, and a pruned and thinned tree (Fig. 7).

Peach leaf photosynthesis rate (a) and carbon partitioning between fruits, leaves, aerial wood and roots (b) as predicted by the virtual tree for different levels of fruit thinning and summer pruning. The treatments are unpruned and unthinned tree (Unpr + Unth), unpruned and thinned tree (80% of fruits were removed, Unpr + Th), pruned (50% of leafy shoots were removed) and unthinned tree (Pr + Unth), and pruned and thinned tree (Pr + Th)
The photosynthesis rate was the highest for trees with high fruit load. The pruned thinned tree exhibited also a high photosynthesis rate during the first 10 days of simulation, and a decreased rate when the tree recovered its foliage area. Pruning and thinning had both an effect on growth and reserve accumulation of the different organs. However, the effect was very different on sink and source organs. The sink organs grew more and accumulated more reserves when the leaf:fruit ratio was higher. The shoots grew more when the tree was pruned and, for a given pruning intensity, they grew more when the tree was thinned. The effect of pruning on shoot growth can be interpreted according to the “coordination theory”. Nonetheless, the question arises as to why shoots grow more after thinning when the model assumes that they have a priority for carbon? The analysis of simulations shows that there is more carbon available for roots when the tree is thinned. The subsequent increase of root growth induces an increase of the shoot sink strength due to the application of the “coordination theory”. In most published studies, the negative effect of high fruit load on shoot growth is interpreted as a direct competition between fruits and shoots (Hurd et al. 1979; Grossman and DeJong 1995). Our simulation leads to an alternative interpretation of the fruit load effect on vegetative growth: i.e., the competition between fruits and roots for carbon decreases both the fruit and root growth, and the decrease in shoot growth does not originate from competition with fruits but only results, according to the coordination theory, from the decrease in root growth.
Conclusions
Carbon partitioning in plants is controlled by a number of factors that include photosynthesis, the number and location of competing sinks, storage capacity and vascular transport. Although there is considerable knowledge on individual processes in plants such as photosynthesis, translocation and cell growth, it appears that the controls actually regulating the carbon partitioning at the whole plant level are still poorly understood (Wardlaw 1990; Le Roux et al. 2001). Indeed, many processes are closely interrelated and more integrative research work based on modelling is greatly needed. Two main approaches could be followed.
The first approach is a top-down teleonomic approach which assumes that the different processes are interrelated through a common goal. This approach can yield general macroscopic laws applicable to a large range of plant species (West et al. 1997, 1999; Enquist 2002) and is able to represent carbon-partitioning strategies selected by evolutionary pressure. Such an approach is a matter of debate in the community of ecologists, but it has been increasingly applied during the last decade. It can bring a useful framework to modellers of carbon allocation and still requires further research.
The second approach is a bottom-up integration of our knowledge about source–sink and transport and chemical/biochemical models. Such integration must be comprehensive enough to allow simulating emergent properties of complex biological systems. For that purpose, modellers need to collaborate closely with physiologists to base their modelling concepts on realistic physiological theories of plant functioning. Three important areas of research should be strengthened. The first one is the root–shoot–fruit interactions. Many agricultural practices such as thinning, pruning, fertilisation and irrigation strongly influence these interactions and improvement of knowledge and models in that field are needed for designing more efficient crop models. The challenge is here to make root, plant and fruit scientists working together. The second one is the transport in the plant architecture of material and signals, with a special focus on interactions between compounds (e.g., water and sugars, sugars and hormones, ...). Special methodological investments are required to provide direct measurements of the various parameters (i.e., phloem resistance) and of the driving variables of the transport processes (turgor pressure and osmotic potential of the phloem) that are essential to validate and to improve transport models. The last important challenging area is the carbon allocation within the fruit tissues. Indeed this aspect, which is missing in the fruit models (Génard et al. 2007), is very important for fruit quality because it determines the size of the tissues and their composition in terms of sugars, acids, and other relevant biochemical compounds to the flavor and/or gustative properties of the fruits.
The development of more comprehensive models of carbon allocation will allow the analysis of the effects of plant management, as shown in this paper, and of variation in plant genetic properties on plant growth and fruit quality. Genetic analyses become possible when values of model parameters can be based on genetic information, as done on fruits trees by Quilot et al. (2005). Steps to include genetic information in plant models have been made by Buck-Sorlin et al. (2005), which open the way to explore genotype × environment interactions.
References
Adam B, Donès N, Sinoquet H (2004) VegeSTAR v.3.1: a software to compute light interception and photosynthesis by 3D plant mock-ups. In: Godin C, Hanan J, Kurth W et al (eds) Proceedings of the 4th international workshop on functional–structural plant models, 7–11 June 2004, Montpellier. Montpellier, p 414
Allen MT, Prusinkiewicz P, DeJong TM (2005) Using L-systems for modeling source–sink interactions, architecture and physiology of growing trees: the L-PEACH model. New Phytol 166:869–880
Atkinson D (1980) The distribution and effectiveness of the roots of tree crops. Hortic Rev 2:424–490
Balandier P, Lacointe A, Le Roux X, Sinoquet H, Cruiziat P, Le Dizes S (2000) SIMWAL: a structural–functional model simulating single walnut tree growth in response to climate and pruning. Ann For Sci 57:571–585
Ball J, Woodrow I, Berry J (1987) A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions. In: Biggens J (ed) Progress in photosynthesis research. Proceedings of the VIIth international photosynthesis congress, vol IV. Martinus Nijhoff Publishers, Dordrecht, pp 221–224
Bancal P, Soltani F (2002) Source–sink partitioning. Do we need Münch? J Exp Bot 25:1919–1928
Bassow SL, Ford ED (1990) A process based model of carbon translocation in trees: an exploration of the branch autonomy theory. Silva Carelica 15:77–87
BenMimoun M, Longuenesse JJ, Génard M (1996) Pmax as related to leaf:fruit ratio and fruit assimilate demand in peach. J Hortic Sci 71:767–775
Bidel LPR, Pagès L, Rivière LM, Pelloux G, Lorendeau JY (2000) MassFlowDyn I: a carbon transport and partitioning model for root system architecture. Ann Bot 85:869–886
Bosc A (2000) EMILION, a tree functional–structural model: presentation and first application to the analysis of branch carbon balance. Ann For Sci 57:555–569
Bruchou C, Génard M (1999) A space–time model of carbon translocation along a shoot bearing fruits. Ann Bot 84:565–576
Buck-Sorlin GH, Kniemeyer O, Kurth W (2005) Barley morphology, genetics and hormonal regulation of internode elongation modelled by a relational growth grammar. New Phytol 166:859–867
Cannell MGR (1971) Production and distribution of dry matter in trees of Coffea arabica L. in Kenya as affected by seasonal climatic differences and the presence of fruit. Ann Appl Biol 67:99–120
Cannell MGR (1985) Attributes of trees as crop plants. In: Cannell MGR, Jackson JE (eds) Dry matter partitioning in tree crops, National Environment Research Council. Penicuik, Great Britain, pp 160–193
Causton DR, Venus JC (1981) The biometry of plant growth. Edward Arnold, London
Chalmers DJ, Canterford RL, Jerie PH, Jones TR, Ugalde TD (1975) Photosynthesis in relation to growth and distribution of fruit in peach trees. Aust J Plant Physiol 2:635–645
Chang R (2000) Physical chemistry for the chemical and biological sciences. University Science Books, Sausalito
Charles-Edwards DA (1976) Shoot and root activities during steady-state plant growth. Ann Bot 40:767–772
Charles-Edwards DA, Thornley HM (1973) Light interception by isolated plant: a simple model. Ann Bot 37:919–928
Chelle M, Andrieu B (1998) The nested radiosity model for the distribution of light within plant canopies. Ecol Modell 111:75–91
Chen J, Reynolds JF (1997) A coordination model of whole-plant carbon allocation in relation to water stress. Ann Bot 80:45–55
Costes E (2004) Integrating knowledge of tree biology and physiology into models of fruit tree development: a review. Acta Hortic 636:575–589
Dale JE, Sutcliffe JF (1986) Water relations of plant cells. In: Steward FC, Sutcliffe JF, Dale JE (eds) Plant physiology: a treatise, vol 9, water and solutes in plants. Academic, Orlando, pp 1–48
Daudet FA, Lacointe A, Gaudillère JP, Cruiziat P (2002) Generalized Münch coupling between sugar and water fluxes for modelling carbon allocation as affected by water status. J Theor Biol 214:481–498
Dauzat J, Eroy MN (1997) Simulating light regime and intercrop yields in a coconut based farming system. Eur J Agron 7:63–74
Dauzat J, Franck N, Vaast P (2006) Using virtual plants for upscaling carbon assimilation from the leaf to the canopy level. Application to coffee agroforestry systems. In: Proceedings of the 21st international scientific colloquium on coffee. ASIC, pp 1037–1044
Davidson RL (1969) Effect of root/leaf temperature differentials on root/shoot ratios in some pasture grasses and clover. Ann Bot 25:59–104
Droppelmann K, Berliner P (2000) Biometric relationships and growth of pruned and non-pruned Acacia saligna under runoff irrigation in northern Kenya. For Ecol Manag 126:349–359
Enquist BJ (2002) Universal scaling in tree and vascular plant allometry: toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiol 22:1045–1064
Ericsson T, Rytter L, Vapaavuori E (1996) Physiology of carbon allocation in trees. Biomass Bioenergy 11:115–127
Eschenbach C (2005) Emergent properties modelled with the functional structural tree growth model ALMIS: computer experiments on resource gain and use. Ecol Modell 186:470–488
Evers JB, Vos J, Fournier C, Andrieu B, Chelle M, Struik PC (2005) Towards a generic architectural model of tillering in Gramineae, as exemplified by spring wheat (Triticum aestivum). New Phytol 166:801–812
Farquhar GD, von Caemmerer S, Berry JA (1980) A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149:78–90
Fishman S, Génard M (1998) A biophysical model of fruit growth: simulation of seasonal and diurnal dynamics of mass. Plant Cell Environ 21:739–752
Ford ED (1992) The control of tree structure and productivity through the interaction of morphological development and physiological processes. Int J Plant Sci 153:147–162
Franck N, Vaast P, Génard M, Dauzat J (2006) Soluble sugars mediate sink feedback down-regulation of leaf photosynthesis in field-grown Coffea arabica. Tree Physiol 26:517–525
Génard M, Lescourret F (2004) Modelling fruit quality: ecophysiological, agronomical and ecological perspectives. In: Dris R, Jain SM (eds) Production practices and quality assessment of food crops, vol 1, preharvest practice. Kluwer, Dordrecht, pp 47–82
Génard M, Souty M (1996) Modeling the peach sugar contents in relation to fruit growth. J Am Soc Hortic Sci 121:1122–1131
Génard M, Pagès L, Kervella J (1998) A carbon balance model of peach tree growth and development for studying the pruning response. Tree Physiol 18:351–362
Génard M, Lescourret F, Gomez L, Habib R (2003) Changes in fruit sugar concentrations in response to assimilate supply, metabolism and dilution: a modeling approach applied to peach fruit (Prunus persica). Tree Physiol 23:373–385
Génard M, Bertin N, Borel C, Bussières P, Gautier H, Habib R, Léchaudel M, Lecomte A, Lescourret F, Lobit P, Quilot B (2007) Towards a virtual fruit focusing on quality: modelling features and potential uses. J Exp Bot 58:917–928
Godin C, Sinoquet H (2005) Functional–structural plant modelling. New Phytol 166:705–708
Grechi I, Vivin P, Hilbert G, Milin S, Robert T, Gaudillère JP (2007) Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine. Environ Exp Bot 59:139–149
Grossman YL, Dejong TM (1994) Peach—a simulation-model of reproductive and vegetative growth in peach-trees. Tree Physiol 14:329–345
Grossman YL, Dejong TM (1995) Maximum fruit-growth potential following resource limitation during peach growth. Ann Bot 75:561–567
Gucci R, Grappadelli LC, Tustin S, Ravaglia G (1995) The effect of defruiting at different stages of fruit-development on leaf photosynthesis of golden-delicious apple. Tree Physiol 15:35–40
Havis AL (1962) Effects of time of fruit thinning of Redhaven peach. Proc Am Soc Hortic Sci 80:172–176
Henton SM, Greaves AJ, Piller GJ, Minchin PEH (2002) Revisiting the Münch pressure-flow hypothesis for long-distance transport of carbohydrates: modelling the dynamics of solute transport inside a semipermeable tube. J Exp Bot 53:1411–1419
Hölttä T, Vesala T, Sevanto S, Perämäki M, Nikinmaa E (2006) Modeling xylem and phloem water flows in trees according to cohesion theory and Münch hypothesis. Trees 20:67–78
Hurd RG, Gay AP, Mountifield AC (1979) The effect of partial flower removal on the relation between root, shoot and fruit growth in the indeterminate tomato. Ann Appl Biol 93:77–89
Iglesias DJ, Lliso I, Tadeo FR, Talon M (2002) Regulation of photosynthesis through source: sink imbalance in citrus is mediated by carbohydrate content in leaves. Physiol Plant 116:563–572
Johnson DS (1995) Effect of flower and fruit thinning on the maturity of ‘Cox’s Orange Pippin’ apples at harvest. J Hortic Sci 70:541–548
Keller A (1996) Quasi-Monte Carlo methods in computer graphics: the global illumination problem. Lect Appl Math 32:455–469
Kikuchi T, Asada T, Shiozaki Y, Saito T, Kamata S (1989) Effect of summer pruning on the next season’s shoot growth of young apple trees. J Japan Soc Hortic Sci 58:491–497
King DA (2005) Linking tree form, allocation and growth with an allometrically explicit model. Ecol Modell 185:77–91
Kurth W (1998) Some new formalisms for modelling the interactions between plant architecture, competition and carbon allocation. Bayreuther Forum Ökologie 52:53–98
Lacointe A (2000) Carbon allocation among tree organs: a review of basic processes and representation in functional–structural tree models. Ann For Sci 57:521–533
Léchaudel M, Génard M, Lescourret F, Urban L, Jannoyer M (2002) Leaf-to-fruit ratio affects water and dry-matter content of mango fruit. J Hortic Sci Biotechnol 77:773–777
Léchaudel M, Génard M, Lescourret F, Urban L, Jannoyer M (2005) Modelling effects of weather and source–sink relationships on mango fruit growth. Tree Physiol 25:583–597
Le Roux X, Lacointe A, Escobar-Gutierrez A, Dizes S (2001) Carbon-based models of individual tree growth: a critical appraisal. Ann For Sci 58:469–506
Lescourret F, Génard M (2005) A virtual peach fruit model simulating changes in fruit quality during the final stage of fruit growth. Tree Physiol 25:1303–1315
Lescourret F, Ben-Mimoun M, Génard M (1998) A simulation model of growth at the shoot-bearing fruit level. I. Description and parameterization for peach. Eur J Agron 9:173–188
Lescourret F, Génard M, Habib R, Fishman S (2001) Variation in surface conductance to water vapor diffusion in peach fruit and its effects on fruit growth assessed by a simulation model. Tree Physiol 21:735–741
Leuning R (1995) A critical appraisal of a combined stomatal-photosynthesis model for C3 plants. Plant Cell Environ 18:339–355
List R, Küppers M (1998) Light climate and assimilate distribution within segment oriented woody plants growth in the simulation program MADEIRA. In: Kastner-Maresch A, Kurth W, Sonntag M, Breckling B (eds) Individual-based structural and functional models in ecology. Bayreuther Forum Ökologie 52:141–153
Marcelis LFM (1993) Effects of assimilate supply on the growth of individual cucumber fruits. Physiol Plant 87:313–320
Marcelis LFM (1996) Simulation of biomass allocation in greenhouse crops: a review. Acta Hortic 328:49–67
Minchin PEH, Thorpe MR, Farrar JF (1993) A simple mechanistic model of phloem transport which explains sink priority. J Exp Bot 44:947–955
Muller-Landau HC, Condit RS, Chave J, Thomas SC, Bohiman SA, Bunyavejchewin S, Davies S, Foster R, Gunatilleke S, Gunatilleke N, Harms KE, Hart T, Hubbell SP, Itoh A, Kassim AR, LaFrankie JV, Lee HS, Losos E, Makana JR, Ohkubo T, Sukumar R, Sun IF, Nur Supardi MN, Tan S, Thompson J, Valencia R, Villa Muños G, Wills C, Yamakura T, Chuyong G, Shivaramaiah Dattaraja H, Esufali S, Hall P, Hernandez C, Kenfack D, Kiratiprayoon S, Suresh HS, Thomas D, Vallejo MI, Ashton P (2006) Testing metabolic ecology theory for allometric scaling of tree size, growth and mortality in tropical forests. Ecol Lett 9:575–588
Neri D, Sansavini S, Sugiyama N (1992) Summer and root pruning of split-root potted peach trees. Acta Hortic 322:177–189
Niklas KJ, Enquist BJ (2003) An allometric model for seed plant reproduction. Evol Ecol Res 5:79–88
Ögren E (1991) Prediction of photoinhibition of photosynthesis from measurements of fluorescence quenching components. Planta 184:538–544
Pearcy RW, Yang W (1996) A three-dimensional crown architecture model for assessment of light capture and carbon gain by understory plants. Oecologia 108:1–12
Perttunen J, Sievänen R, Nikinmaa E, Salminen H, Saarenmaa H, Vakeva J (1996) Lignum a tree model based on simple structural units. Ann Bot 77:87–98
Perttunen J, Sievänen R, Nikinmaa E (1998) LIGNUM: a model combining the structure and the functioning of trees. Ecol Modell 108:189–198
Proebsting EL, Mills HH (1981) Effects of season and crop load on maturity characteristics of ‘Bing’ cherry. J Am Soc Hortic Sci 106:144–146
Quilot B, Génard M, Kervella J, Lescourret F (2002) Ecophysiological analysis of genotypic variation in peach fruit growth. J Exp Bot 53:1613–1625
Quilot B, Génard M, Lescourret F, Kervella J (2005) Simulating genotypic variation of fruit quality in an advanced peach × Prunus davidiana cross. J Exp Bot 56:3071–3081
Reginato G, Robinson T, García de Cortázar V (2007) Predicted crop value for nectarines and cling peaches of different harvest season as a function of crop load. HortScience 42:239–345
Reynolds JF, Chen JL (1996) Modelling whole-plant allocation in relation to carbon and nitrogen supply: coordination versus optimization: opinion. Plant Soil 185:65–74
Reynolds JF, Thornley JHM (1982) A shoot:root partitioning model. Ann Bot 49:585–597
Röhrig M, Stützel H, Alt C (1999) A three-dimensional approach to modeling light interception in heterogenous canopies. Agron J 91:1024–1032
Sharkey PJ, Pate JS (1976) Translocation from leaves to fruits of a legume, studied by a phloem bleeding technique: diurnal changes and effects of continuous darkness. Planta 128:63–72
Shinozaki K, Yoda K, Hozumi K, Kira T (1964) A quantitative analysis of plant form. The pipe model theory. II. Further evidence of the theory and its application in forest ecology. Japan J Ecol 14:133–139
Sievänen R, Nikinmaa E, Nygren P, Ozier-Lafontaine H, Perttunen J, Hakula H (2000) Components of functional–structural tree models. Ann For Sci 57:399–412
Sinoquet H, Le Roux X, Adam B, Ameglio T Daudet A (2001) RATP: a model for simulating the spatial distribution of radiation absorption, transpiration and photosynthesis within canopies: application to an isolated tree crown. Plant Cell Environ 24:395–406
Soler C, Sillion FX, Blaise F, De Reffye (2003) An efficient instantiation algorithm for simulating radiant energy transfer in plant models. ACM Trans Graph 22:204–233
Syvertsen JP, Goñi C, Otero A (2003) Fruit load and canopy shading affect leaf characteristics and net gas exchange of ‘Spring’ navel orange trees. Tree Physiol 23:899–906
Thompson MV, Holbrook NM (2003) Application on a single-solute non-steady-state phloem model to the study of long-distance assimilate transport. J Theor Biol 220:419–455
Thornley JHM (1972) A model to describe the partitioning of photosynthate during vegetative plant growth. Ann Bot 36:419–430
Thornley JHM (1998) Modelling shoot:root relations: the only way forward? Ann Bot 81:165–171
Thornley JHM, Johnson IR (1990) Plant and crop modelling. Oxford University Press, NY
Upmeyer DJ, Koller HR (1973) Diurnal trends in net photosynthetic rate and carbohydrate levels of soybean leaves. Plant Physiol 51:871–874
Vaast P, Dauzat J, Génard M (2002) Modeling the effects of fruit load, shade and plant water status on coffee berry growth and carbon partitioning at the branch level. Acta Hortic 584:57–62
Vaast P, Angrand J, Franck N, Dauzat J, Génard M (2005) Fruit load and branch ring-barking affect carbon allocation and photosynthesis of leaf and fruit of Coffea arabica in the field. Tree Physiol 25:753–760
Vos J, Marcelis LFM, Evers JB (2007) Functional–structural plant modelling in crop production. In: Vos J, Marcelis LFM, de Visser PHB, Struik PC, Evers JB (eds) Functional–structural plant modelling in crop production. Springer, The Netherlands, pp 1–12
Wardlaw IF (1990) Tansley review no. 27. The control of carbon partitioning in plants. New Phytol 116:341–381
Webster AD, Shepherd UM (1984) The effect of summer shoot tipping and rootstock on the growth, floral bud production, yield and fruit quality of young sweet cherries. J Hortic Sci 59:175–182
West GB, Brown JH, Enquist BJ (1997) A general model for the origin of allometric scaling laws in biology. Science 276:122–126
West GB, Brown JH, Enquist BJ (1999) A general model for the structure and allometry of plant vascular systems. Nature (Lond) 400:664–667
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. Treutter.
Rights and permissions
About this article
Cite this article
Génard, M., Dauzat, J., Franck, N. et al. Carbon allocation in fruit trees: from theory to modelling. Trees 22, 269–282 (2008). https://doi.org/10.1007/s00468-007-0176-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-007-0176-5