
Abstract
Anthropogenic activities have increased disturbances and alien woody invasion in mountain ecosystems worldwide. Whether disturbances promote or counteract upward movement of woody aliens is poorly understood. We assessed if the most successful woody invader of low mountains of central Argentina (Gleditsia triacanthos) might expand its elevational distribution in response to the principal disturbances of these ecosystems (fire and livestock browsing) across increasing climatic severity. We assessed seedling emergence, growth and mycorrhizal colonization on sown plots distributed in burned and unburned sites, with and without browsing at the lower and upper elevation belts (i.e. 1000 and 2400 m a.s.l.). Additionally, several abiotic variables were measured to relate their influence on the seedling establishment. Disturbances reduced seedling emergence at both elevations. Burned conditions increased seedling growth and arbuscular colonization only in the lower belt. Seedling success (total seedling biomass per plot) was not modified by disturbances at the upper elevation, but was reduced by browsing and enhanced by fire in the lower elevation. The overall reduction in seedling emergence and growth in the upper elevation despite the higher soil nutrient content places climate as the strongest regulator of G. triacanthos seedling establishment. Accordingly, climate rather than disturbances would be the main limiting factor of upward expansion of this woody alien. Our findings differ from general patterns described for mountain invasion by herbaceous species, highlighting that mountain invasibility is highly growth-form dependent, and that upper range expansion by woody aliens interacting with multiple disturbances should be assessed worldwide.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Biological invasions have been described as one of the major drivers of global change (Sala et al. 2000). Mountain ecosystems are still mostly free of alien plants (Pauchard et al. 2009; Pyšek et al. 2011) and climate has been assumed as the main responsible factor limiting alien species’ spread towards highlands (Millennium Ecosystem Assessment 2005). However, alien plants are currently spreading upward, possible mediated by the increase in global temperature, propagule pressure and anthropogenic disturbances (Alexander et al. 2016). While disturbances might promote alien expansion in the lower belt, their influence on alien expansion toward higher elevations remain unclear, mainly due to opposite evidences (e.g., Callaway et al. 2002; D’Antonio et al. 2001; Milbau et al. 2013; Lembrechts et al. 2016). Additionally, it has been suggested that the magnitude and direction of the combined effects of multiple disturbances on alien expansion are dependent on species and ecosystem properties (Lonsdale 1999; Dietz and Edwards 2006).
In mountain ecosystems with marked climatic gradients, competition usually dominates the outcome of alien success at the lower elevation, whereas facilitation dominates at higher elevations (Callaway et al. 2002; D’Antonio et al. 2001). Two of the most important disturbances influencing alien plant dynamics are fire and browsing by domestic cattle. Fire removes native vegetation, which might have antagonist effects on post-burned seedling establishment, depending on elevation. For example, fire might relax competition at low elevations, promoting seedling emergence and growth, whereas it might interfere in facilitation interactions at high elevations (D’Antonio et al. 2001; Callaway et al. 2002). Indeed, vegetation removal imposed by fire might increase seedling stress by decreasing soil humidity and increasing soil temperature fluctuations (Certini 2005); all these effects might reduce seedling establishment mainly in the upper belt of mountains (Körner 1999). Additionally, along the entire elevation gradient, fire might enhance seedling growth through nutrient release into the soil (Quiroz et al. 2011). On the other hand, browsing by livestock reduces seedling establishment particularly in alien palatable species (Mazía et al. 2001; Capó et al. 2016). Moreover, seedling resprouting after browsing may be reduced under the harsher climates that prevail at higher elevations (Körner 1999; Marcora et al. 2013). Alternatively, browsing might promote alien seedling establishment if livestock browse selectively on native species (Kimball and Schiffman 2003; Relva et al. 2010). Finally, both disturbances might reduce seedling establishment by increasing soil impedance (Kozlowski 1999).
It has been recently proposed that, to gain a more comprehensive understanding of the plant invasion process, symbiotic interactions between alien plants and soil biota should be considered, since most alien plants need to rely on compatible belowground mutualists in their new environment (Nuñez and Dickie 2014; Urcelay et al. 2017). Although arbuscular mycorrhizal fungi (AMF) are among the most abundant soil symbiotic organisms and are particularly relevant for some range-expanding alien species (e.g., Gundale et al. 2014), there is evidence of a decrease in their diversity and root colonization with elevation (Gai et al. 2012), as well as after a fire (Longo et al. 2014, 2016). Additionally, complex effects of the interaction between herbivores, mycorrhizae and host plants have been observed (Gehring and Whitham 1994). As mycorrhizal colonization might be affected by climate, disturbances and soil characteristics in complex ways (van der Heyde et al. 2017), the indirect effect of AMF on seedling establishment cannot be easily predicted.
Most experiments testing interactive disturbance effects on seedling establishment of invasive plants under contrasting climatic conditions have been performed with grass and forb species, whereas experiments with woody alien species are almost absent in the literature (Stevens and Latimer 2015, but see Milbau et al. 2013). Woody plants were fairly recently recognized as major invaders due to their substantial and growing impacts on biodiversity, ecosystem functioning, and human livelihoods (Richardson et al. 2014). Understanding the mechanisms underlying woody alien upward expansion is, therefore, of key importance, particularly during early establishment, when seedlings are primarily limited by biotic and abiotic factors (Körner 1999; Nuñez and Dickie 2014; Tecco et al. 2016).
In Sierras Grandes Mountains of central Argentina, fire and livestock are the principal drivers of landscape distribution and vegetation dynamics (Cingolani et al. 2003; Argañaraz et al. 2015). While invasion by several woody species threatens most vegetation remnants in the surrounding low-mountain areas (Giorgis and Tecco 2014; Zeballos et al. 2014; Giorgis et al. 2016), the upward expansion of most of them is still incipient (Giorgis et al. 2011a). That is the case of the North American tree Gleditsia triacanthos L. (honey locust), an invader of several ecosystems worldwide (Richardson and Rejmánek 2011), which has been described as one of the most successful woody invaders of low-mountain ecosystems and whose current distribution is still restricted to the lower belts (Giorgis et al. 2011b). Results of seed addition experiments in Sierras Grandes suggest that G. triacanthos can germinate throughout the elevation range but a low proportion of those emerged seedlings can successfully overwinter in the upper belt (Tecco et al. 2016). The removal of vegetation driven by disturbances has been found to be favorable for G. triacanthos establishment in the lowland grasslands (Mazía et al. 2001; Chaneton et al. 2004; Mazía et al. 2010). Finally, this species is browsed by livestock (Capó et al. 2016) and colonized by AMF (Tecco et al. 2013).
Whether post-burned conditions and browsing promote or counteract seedling emergence, growth and mycorrhizal root colonization of G. triacanthos seedlings is unknown; in addition, whether their effects differ between elevations with contrasting environmental conditions is also unknown. Considering that seedling emergence, growth and survival of woody species decrease with elevation in response to climatic conditions in mountains across the world (e.g., Körner 1999; Marcora et al. 2013; Tecco et al. 2016), we hypothesize that the relative influence of disturbances on seedling establishment of G. triacanthos becomes dependent on climate with increasing elevation (D’Antonio et al. 2001; Callaway et al. 2002), with disturbance-driven restrictions being increased and disturbance-driven facilitation being lessened towards the upland. The net outcome of this multiple interaction on seedling success is not straightforward given the antagonist effects that each disturbance can promote. We set out an experimental approach that specifically assessed seedling emergence, growth and AMF root colonization of G. triacanthos on sown plots distributed in (1) burned and unburned sites, (2) with and without livestock browsing and (3) in the lower and the upper mountain belt of a mountain system in central Argentina. Since disturbances might affect different microsite variables that are expected to affect seedling emergence, growth and mycorrhizal colonization, sites were characterized in terms of soil impedance, moisture, temperature and physicochemical soil properties to relate those possible changes to response variables.
Methods
Study site
The study was conducted in the Sierras Grandes Mountains, Córdoba, central Argentina. Mean annual temperature in the lower belt is 15.7 °C, dropping to 7.4 °C at the mountain top (Marcora et al. 2008). There is no frost-free period above 1800 m a.s.l. Mean annual precipitation varies between 750 and 970 mm along the elevational gradient, with most rainfall concentrated in the warmer months, from October to April, and with a positive effect of elevation on rainfall (Colladon et al. 2010). Accordingly, environmental changes along the elevational gradient are characterized by significant decreases in soil temperature and increasing soil moisture towards the upper belt (Tecco et al. 2016).
Two contrasting elevations were selected near the Linderos Road at 1000 (32°5′S; 64°41′W) and 2400 m a.s.l. (32°3′S; 64°54′W), representative of the lower and upper elevational belts. The vegetation of selected areas is characterized by mosaics of tussock grasslands and short grazing lawns in the upper belt (Cingolani et al. 2004), whereas the lower belt is characterized by tall and short grasslands (Giorgis et al. 2011b).
The oldest economic activity in this area is livestock rearing (sheep and cattle), with anthropogenic winter–spring fire to promote forage regrowth (Argañaraz et al. 2015). At the end of winter in September 2013, an extensive fire occurred that affected the entire elevational range of the Sierras Grandes Mountains. The experiment was established 2 months after that fire event.
Study species
Gleditsia triacanthos L. (Fabaceae) is a thorny tree native to the temperate forests of North America, where it tolerates low temperatures (up to − 34 °C in the north of its distribution area; Little 1971). This species has been recorded as a woody invader in several ecosystems worldwide (Rejmánek and Richardson 2013). In the mountain systems of central Argentina, G. triacanthos is the most frequent woody invader of the Chaco Serrano woodlands (i.e., the lowest elevation in this study; Giorgis et al. 2011b), forming almost mono-specific stands or being co-dominant with other native or alien woody species (Fernández et al. 2017). Fruits are consumed and dispersed by cattle (Ferreras and Galetto 2010), which also eat the saplings (Capó et al. 2016). Regardless of the high invasiveness of this species, its elevational distribution remains below 1500 m a.s.l. (Giorgis et al. 2011a, b).
Experimental design
An experimental approach was used to evaluate the effects of post-burned conditions and livestock browsing at two contrasting elevations (i.e., 1000 and 2400 m a.s.l.) on G. triacanthos seedling emergence, growth and mycorrhizal colonization. The lower elevation was chosen as representative of the actual distribution of adult individuals, whereas at the upper elevation the species can potentially establish but is not currently present (Tecco et al. 2016). Elevation sites were chosen in an attempt to encompass similar fire intensity and browsing. At each elevation, we selected a 2-h area subjected to livestock grazing and including both burned and unburned sites. The sown plots were distributed randomly within each burned or unburned site, but ensuring that all had similar micro-topography (e.g., little or no slope). Each sown plot was at a minimum distance of 50 m from another of the same treatment. At each elevation, seeds of G. triacanthos were sown in 18 plots in burned sites and in 18 plots in unburned sites. Half of the sown plots in each burned and unburned site were protected from livestock-browsing using cylindrical cages staked to the ground (Tecco et al. 2016). Rings (45 cm diameter × 50 cm height) were erected with a 2-cm-mesh stainless solid net and walls of these cylindrical structures were further covered with a 1-mm-mesh stainless net. The upper extreme of the rings was also covered with a 1-cm-mesh stainless net. No net was placed at the bottom of the rings. Cages did not prevent arthropod herbivory or neighboring vegetation growth. Uncaged seeds were protected with square (50 × 50 cm) 1-cm-mesh green net staked to the ground, which prevented seeds from being washed away and allowed seedling emergence. This net did not protect the seedlings from herbivores. The experimental design comprised a total of 72 sown plots (2 elevations × 2 fire conditions × 2 browsing conditions × 9 replicates). A total of 100 seeds of G. triacanthos were sown in each plot. Since these seeds have physical dormancy, they were mechanically scarified with sandpaper before sowing (Ferreras et al. 2015).
Site characterization
Burned and unburned sites at both elevations were characterized in terms of livestock stocking rate, soil impedance, soil moisture, soil temperature, physicochemical soil properties and vegetation and bare ground cover. For a better characterization, most of these variables were measured at the beginning (i.e., recently burned conditions) and at the end of the experiment (i.e. end of the growing season) to have a better picture of their changes and potential effects on seedling emergence and growth along the experiment. Soil moisture and temperature were measured with greater frequency (monthly and every 3 h, respectively), because they are expected to suffer high oscillations. Soil physicochemical properties were measured only at the beginning of the experiment due to logistical constraints.
Stocking rate was estimated by calculating the proportion of fresh dung in 50 randomly selected quadrats (30 × 30 cm) per site. Soil impedance (kg cm−2) was measured using a penetrometer (E-280 Pocket Penetrometer) at two randomly selected points in each sown plot. Soil moisture was measured at 7 cm with a “Moisture Probe Meter” (MPB-160-B of ICT International Pty Ltd) at each sown plot (N = 72). Soil temperature was measured with sensors buried at ~ 7 cm depth (Thermochron iButton DS1921G). Three sensors were randomly placed within each elevation and fire condition (N = 12). Soil samples used for physicochemical determination were obtained from the upper 5 cm of each sown plot at the beginning of the experiment. These 72 soil samples were randomly grouped within each treatment to obtain three mixed soil samples per elevation and fire condition (N = 12). Soil was analyzed in terms of pH, organic matter, organic C, Total N, C:N ratio, P, Ca2+ and K+ (see methods in Online Appendix 1).
Vegetation and bare ground cover were measured in all sown plots. For this, percentages of growth forms (i.e., shrubs, herbs, grasses and ferns), rocks, litter and bare ground cover were recorded using a scale ranging from 0.05 (less than 1%), 1, 2, 3, 5, 8, and 10, and increasing every 5% up to 100%.
Seedling establishment
The number of emerged seedlings per plot was recorded monthly from November 2013 to the end of the growing season (April 2014). Occurrence of predation and/or removal of sown seeds by insects was recorded monthly. We considered that predation occurred when observed insects predating at any moment of the experiment or when the sum of seedlings and ungerminated seeds was less than 100. Then, all these signs of seed predation were categorized as presence or absence of seed predation (i.e. each plot had zero or one for seed predation). Seedling growth (i.e., height and plant aboveground dry biomass) was measured at the end of the growing season (April 2014), when all seedlings were harvested. In the laboratory, aboveground biomass was measured with a balance (± 0.001 mg) after drying at 60 °C for 48 h. The total seedling biomass per plot was then recorded as a measure of the success of G. triacanthos seedlings under each condition (i.e., the growth of all seedlings that emerged and survived in each plot). For simplicity and to avoid confusion of this variable with mean individual seedling biomass, we will hereafter refer to total seedling biomass per plot as seedling success.
Root mycorrhizal colonization
To assess mycorrhizal colonization in each seedling, harvested roots were separated and washed with water. All dead and damaged roots were discarded. All thin roots (< 2 mm without apparent suberin) that could be potentially colonized were cleared and stained, following Grace and Stribley (1991). Then they were mounted on semi-permanent slides in polyvinyl–lactic acid–glycerol. Root endophyte was quantified by the magnified intersection method (McGonigle et al. 1990) using a compound microscope (Nikon Eclipse E200), at 200 × magnification. Eighty to one hundred intersections per slide (i.e. per individual) were scored, depending on sample size. Vesicles (lipids reservoirs), arbuscules (intricately branched haustoria are considered the major site of exchange between fungus and host) and hyphae were identified (Smith and Read 2008). The presence of arbuscules in roots is used to determine AMF colonization in plants. However, since these structures are ephemeral and may be absent in roots, hyphae and vesicles are also used to indicate the presence of AMF. Percentage of AMF colonization (i.e., vesicle, arbuscule and total) was assessed as the proportion of total root intersections that were colonized and was calculated as follows:
Fungal colonization = 100 × (Number of intersections with fungal structure/Total number of intersections counted).
Data analysis
Stocking rates were analyzed using GLMM with binary error structure (dung presence/absence; glmer function, lme4 package) using the logit function, and date as random factor (i.e., begging and end of the experiment). The effect of fire, elevation and interactions between these factors on soil impedance, moisture and temperature was analyzed using general linear mixed models (GLMM) with Gaussian error structure correction for the heteroscedasticity (gls function, nlme package) including date as random factor (begging and end for soil impedance, month for soil moisture, hours for temperature). Soil physicochemical properties were analyzed with general linear models (GLM) with Gaussian error structure and correction of heteroscedasticity variance (ident function). Non-metrical multidimensional scaling (NMDS) was performed for percentages of soil cover by growth forms, based on Euclidean distances using vegan package. Furthermore, the envfit function was used to assess which variable (i.e., shrubs, herbs, ferns, grasses, rocks, litter and bare ground cover percentage) was the most important in explaining differences among plots subjected to different treatments at the begging and end of the experiment.
The effect of fire, browsing and elevation and interactions among these factors on seedling emergence, growth and root colonization was assessed. The number of emerged seedlings per plot 1 month after sowing and seedling success were analyzed using GLM with Gaussian error structure correcting for the heteroscedasticity (gls function, nlme package) using the varIdent function. Individual seedling height and dry biomass were analyzed using GLMM with Gaussian error structure (lmer function, lme4 package) with individuals nested into plots. Dry biomass data were log10 transformed. Signals of seed predation per plot (presence/absence) were analyzed via GLM with quasibinomial error structure due to overdispersion. Additionally, percentages of total, arbuscular and vesicular colonization were averaged per plot and analyzed using GLM (gls function). Arbuscular colonization heteroscedasticity was corrected using varIdent function. A posteriori differences were assessed using simultaneous tests for general linear hypotheses (glht function, multicomp package). All the analyses were performed with R version 3.2.4 (R Development Core Team 2016). Differences were considered significant at p ≤ 0.05.
Results
Site characterization
Stocking rate did not differ between elevations or fire conditions (Table 1). Soil impedance was higher in burned than in unburned plots in the lower belt (Table 1). Soil moisture and temperature were highest at the lower elevation, and differed between burned and unburned plots in the upper belt (Table 1). Differences in most soil physicochemical properties were determined by elevation rather than by fire (Table 1), i.e., soils of the upper belt showed significantly higher values of organic matter, C and N, whereas pH, Ca2+ and K+ were higher in soils of the lower belt. The highest soil P content was recorded in burned plots in the upper belt (Table 1).
The main differences in soil cover of sown plots at the beginning of the experiment (i.e., 2 months after fire) were between fire conditions rather than elevations (Fig. 1). Burned plots showed a higher percentage of bare ground, whereas unburned plots exhibited the highest grass, forb and litter cover percentages (Table 2). At the end of the experiment, the differences between burned and unburned plots were less marked and showed lower soil cover heterogeneity than at the beginning of the experiment (Fig. 1). Nonetheless, percentages of bare ground remained high at the end of the experiment in burned plots, and differences between elevations became more important, particularly due to grass and herb cover (Table 2). All in all, at the beginning and end of the experiment, the percentages of bare ground (r2 = 0.95, p = 0.001; r2 = 0.79, p = 0.001, respectively), grass cover (r2 = 0.80, p = 0.001; r2 = 0.70, p = 0.001) and herb cover (r2 = 0.62, p = 0.001; r2 = 0.73, p = 0.001) were the most important variables explaining the differences among sown plots (Fig. 1).

Non-metrical multidimensional scaling (NMDS) on soil cover of sown plots both at the a start (initial) and b end (final) of the experiment (2 and 7 months after burning, respectively). The resulting polygons for each category (burn, unburned, 1000 and 2400 m a.s.l.) are graphed to facilitate interpretation. Vectors represented each soil cover categories including shrubs (Sh), ferns (Fs), herbs (Hs), grasses (Gs), rocks (Rs), litter (Lr), and bare ground (Bsc)
Seedling establishment
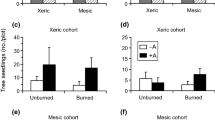
Seedling emergence was significantly higher in undisturbed plots (i.e., unburned without browsing) than in disturbed plots, irrespective of elevation (Fig. 2a; Table 3). Differences between treatments in terms of height and individual seedling aboveground biomass were also observed (Table 3). In the lower belt, seedling height and biomass were highest in burned plots, whereas biomass was lowest in plots exposed to livestock browsing (Fig. 2b, c; Table 3). Contrarily, no effects of fire or browsing were detected at the upper belt.

Seedling establishment of Gleditsia triacanthos in response to elevation, fire and browsing conditions. a Seedling emergence 1 month after sowing; b seedling height; c individual seedling biomass; d arbuscular mycorrhizal colonization (AMC). The last three variables were measured per individual seedling at the end of the growing season. Bars correspond to means + 1 SE. Letters indicate mean differences between treatments (Tukey, p ≤ 0.05)
In terms of seedling success, total seedling biomass per plot was not modified by browsing at the upper belt, but was reduced by browsing at the lower belt (Fig. 3). Contrarily, fire had no effects on total seedling biomass at both elevations (Table 3).

Total aboveground biomass of G. triacanthos per plot at the end of one growing season. Bars correspond to means + 1 SE. Letters indicate mean differences between treatments (Tukey, p ≤ 0.05)
Predation of sown seeds by ants (mainly Acromyrmex lundi) was recorded before seedling emergence and also differed among treatments. There were signs of seed predation in 83% of burned plots vs 19% of unburned plots [ZFire = 4.820; p(> |Z|) < 0.0001], irrespective of elevation or browsing [ZElevation= 1.864, p(> |Z|) = 0.06 and ZBrowsing= 0.07, p(> |Z|) = 0.94].
Root mycorrhizal colonization
Both individually and combined, fire and browsing tended to promote total mycorrhizal colonization at both elevations, although differences in mycorrhizal percentage were not statistically significant (Table 3). Colonization by arbuscules was significantly higher in burned plots, but only at the lower elevation (Fig. 2d; Table 3). There were no differences in vesicle colonization among treatments.
Discussion
The net effects of combined disturbances and climate on the upward expansion of alien plants, particularly woody species, are poorly understood (Alexander et al. 2016; Pauchard et al. 2009). Indeed, disentangling these complex interactions in which several local factors are involved highlights the need for local assessments (Lonsdale 1999; Alba et al. 2015). Therefore, our work contributes novel data for the understanding of upward spread of woody aliens, through the evaluation of some of the main factors structuring vegetation in mountains (i.e., fire, browsing and climate). In line with global patterns, seedling success decreased with elevation, confirming that climate is the major constraint (Körner 1999) despite the increasing soil quality towards the upper elevations. Yet, and in accordance with our hypothesis, the response of G. triacanthos seedling emergence, growth and mycorrhizal root colonization to post-burned conditions and browsing varied with elevation, as reflected by most of the significant interaction terms (Table 3). The magnitude and direction of those responses, however, did not follow a common trend, but seemed to be dependent on climate and not on other factors that change between elevations and characterize sites.
Disturbance-driven changes in microenvironment regardless of elevation
Site characterization showed that disturbances had similar effects in both elevations, that is, fire explained the main differences in vegetation and bare ground cover between burned and unburned sites regardless of elevation (Table 2, Fig. 1) and livestock rates did not differ between elevations or burned conditions (Table 1).
Several biotic and abiotic factors that affect seedling establishment are expected to change along elevation gradients and to be modified by disturbance (Körner 1999). Accordingly, all physicochemical soil variables differed between elevations and some of them between fire conditions in the studied mountains (Table 1). Soils from the upper belt exhibited better conditions for seedling establishment—lower impedance and higher nutrient content—than soils from the lower belt. In turn, the lower belt seemed to provide better conditions—higher soil moisture and temperature. Fire increased this trend of differences between elevations in terms of impedance and moisture (i.e., the highest impedance in the lower belt and the lowest soil moisture in the upper belt) and soil quality (i.e., increasing soil phosphorus content in burned plots only in the upper elevation). Overall, these results suggest some antagonism between climatic and the micro-environmental abiotic scenarios to which seedlings are exposed along the elevation gradient (i.e., better climatic conditions on worse physicochemical soil conditions).
Disturbance-driven changes in seedling establishment
Fire had antagonistic effects at each development stage of G. triacanthos establishment, i.e., negative on seedling emergence and positive on growth (Fig. 2a–c), particularly in the lower belt. Accordingly, the final outcome was an absence of fire-driven changes in seedling success at both elevations (Fig. 3).
Livestock browsing had negative effects on seedling emergence at both elevations, and on seedling growth at the lower belt. Although browsing effects might become more negative with elevation due to lower resprouting rates under harsher climatic conditions (Marcora et al. 2013), the negative effects of browsing were not increased at the upper elevation; rather, the opposite occurred, i.e., browsing exclusion showed the highest effect in seedling success at the lowest elevation (Fig. 3), even when stocking rates were similar between elevations (Table 1).
Assessing each seedling development stage separately might help us to understand the disturbance-driven mechanisms affecting early establishment of the species. Seedling emergence was higher in undisturbed plots at both elevations suggesting that both disturbances would be constraining, but not hindering, the expansion of this invader at the early seedling establishment stage (te Beest et al. 2015). Livestock might reduce seedling emergence through browsing and trampling (Adams 1975; Tjelele et al. 2015). In turn, environmental stresses (Körner 1999) and seed exposure to predation (Muschetto et al. 2015) could be hypothesized as possible explanations to the lower emergence in burned plots. For instance, given the high percentage of bare ground in burned plots (Table 2), seeds and seedlings might have been exposed to harsh microclimatic conditions, such as drastic oscillations in temperature and humidity in surface soil layers (Gimeno-García et al. 2000). Seed predation is one of the most important factors determining the lack of seedling regeneration of many species (Ordóñez and Retana 2004 and references therein). In our study, we observed ants removing pieces of imbibed seeds. Ants are among the most efficient seed predators in burned areas (Ordóñez and Retana 2004) and the abundance of seed-harvesting ants increases after a fire in some mountain ecosystems (Arnan et al. 2007).
Regarding seedling growth, fire increased both individual seedling height and biomass only at the lower elevation (Fig. 2b, c). In grasslands, grasses and forbs are superior competitors over tree seedlings (De Steven 1991); therefore, this result is consistent with the reduced competition due to vegetation removal by fire (Chaneton et al. 2004; Tjelele et al. 2015). This mechanism could be expected to be at play in the study area given the overall reduction in neighboring vegetation and litter after the fire (Table 2). Contrarily, the expected increase in soil nutrients induced by fire would not explain the increased seedling growth, considering that the highest phosphorus concentration was found in burned sites at the upper elevation (Table 1), whereas the highest seedling growth was recorded in burned sites at the lower elevation (Fig. 2b, c).
The effects of fire on emergence and growth of G. triacanthos reported for this mountain system partly differ from those observed in Inland Pampas Grasslands in central Argentina, where burning increased seedling emergence in wet years, whereas growth of establishing trees was significantly reduced in burned grassland plots (Chaneton et al. 2004; Mazía et al. 2010). These differences further support the hypothesis that the influence of different disturbances on seedling establishment and performance of alien species would be variable among ecosystems (Alba et al. 2015).
While total mycorrhizal colonization was similar in both elevations and disturbance conditions, an increase in arbuscule colonization was observed in burned plots only at the lowest elevation. This increase of arbuscule colonization in burned sites might occur via changes in AMF spore composition, i.e., higher abundance of ruderal AMF species that increase colonization under stressful conditions (Grümberg et al. 2015; Longo et al. 2016). Contrarily, at the upper elevation the arbuscular colonization was not affected by disturbances possibly due to contrasting strategies between fungal communities along the gradient. Indeed, stress-tolerant fungus species are expected to dominate under harsh conditions such as those of highlands, whereas ruderal and competitive fungus species would prevail at lower elevations (Chagnon et al. 2013). For instance, unlike ruderal AMF species, stress-tolerant species are predicted to respond slowly to disturbances and reduced competition. Most importantly, the absence of differences in arbuscule colonization between treatments in the upper elevation mirrored the pattern of seedling growth at this elevation, and further supports climate as the major constraint on upward expansion. While the benefits that increased arbuscular colonization provides to G. triacanthos seedlings need to be confirmed through experimental testing, the patterns observed in the field do show maximum seedling growth under burned conditions in the lowest elevation. Plant interactions with soil biota have proved to be relevant to understand the success of some range-expanding alien species (e.g., Gundale et al. 2014). Accordingly, our findings on AMF colonization suggest that the plant–fungus interaction might influence plant invasiveness. Our results also call for specific experimental assessment and further highlight the relevance of soil biota in mountain invasion (Urcelay et al. 2017).
Context-dependent effects of fire and browsing on alien invasion
Seedling success of G. triacanthos was severely reduced in the upper belt in the absence of disturbances, whereas in the lower belt, the individual and interacting effects of disturbances boosted changes in seedling establishment (i.e., main disturbance-driven responses in growth or AMF recorded at the lower elevation). This result is in line with our hypothesis, which postulates that the relative influence of disturbances on vegetation processes becomes dependent on climate with increasing elevation (D’Antonio et al. 2001; Callaway et al. 2002). We infer supremacy of climate over soil physicochemical properties underlying seedling success, since our results show that climatic restrictions overcompensate favorable soil nutrient conditions for seedling growth towards increasing elevation. Similarly, other studies found that harsher climatic conditions constrain alien seedling growth, despite nutrient increases and competitive releases promoted by disturbances (Poll et al. 2009; Ansari and Daehler 2010). The predominance of climate influence might also explain the lack of increase in seedling growth in burned plots at the upper elevation despite the higher phosphorus content in soils (Table 1). Our results disagree with those of Lembrechts et al. (2015) for high-latitude mountain regions, who reported disturbance as the main positive determinant of invasion success of herbaceous species along the entire elevation gradient. These differences further highlight the importance of discriminating major growth forms when exploring general invasion patterns, since the different forms respond differently to local environmental and climatic pressures (Larson et al. 2001; Giorgis et al. 2016). Studies involving other woody species and mountain systems are necessary to confirm the generality of this pattern.
The overall success of this woody alien species in this fire-prone mountain ecosystem (Argañaraz et al. 2015) will certainly depend on fire response at other stages of its life cycle. G. triacanthos is known to have heat-tolerant seeds (Ferreras et al. 2015), post-fire seedling establishment (this study), and post-fire survival of adult trees through resprouting (Herrero et al. 2016). These responses show that this invader is equally adapted to fire as the resident woody species (Torres et al. 2014; Jaureguiberry and Díaz 2015). Cattle browsing might prevail as the major constraint to its persistence and expansion (Capó et al. 2016) in the lower belt of central Argentina mountains, despite the role of cattle as seed dispersers (Ferreras and Galetto 2010).
Fewer but worse: upward spread of woody aliens
As forbs and graminoids are the major components of the alien flora in mountain systems (Alexander et al. 2016), the pooling of all growth forms suggests that herbaceous species might drive general patterns of mountain invasion. Although less representative in number, woody aliens are also undergoing upper range expansion (Pauchard et al. 2009; Giorgis et al. 2011a; Alexander et al. 2016), which—given their overwhelming impact worldwide (Richardson and Rejmánek 2011)—might become an important threat to the unique biodiversity and important ecosystem services of mountain systems (Pauchard et al. 2009; Alexander et al. 2016).
The presence of livestock rearing throughout the mountain range might guarantee the effective dispersal of G. triacanthos and helps to overcome the propagule pressure limitation that characterizes most mountain invasion (Pauchard et al. 2009; Giorgis et al. 2016). In turn, we suggest that climate rather than disturbances might be the main limiting factor for G. triacanthos upward expansion in mountains of central Argentina. However, the potential for seedling emergence throughout the elevation gradient reinforces that climate alone might not prevent early seedling establishment of this invader well above its current elevation distribution (Tecco et al. 2016). Accordingly, upper range expansion of several woody alien species is expected in this mountain ecosystem (Tecco et al. 2016; Giorgis et al. 2011a, 2016; Urcelay et al. 2017). Although their response to multiple disturbances along the elevational gradient remains to be tested, woody native and alien species have shown high post-fire survival (Herrero et al. 2016), which might allow for their spread once propagule pressure restrictions towards higher elevations are overcome (Giorgis et al. 2016), emphasizing that climate would be the most important limiting factor for this growth form.
Upward movement of lowland woody alien species has also been reported for other mountains worldwide (Pauchard et al. 2009; Alexander et al. 2016). Even if climate proves a major limitation for early establishment in several woody species in high mountains, global warming might reduce these restrictions and consequently allow for acceleration of the invasion process. Detailed understanding of the ecology of individual species is still necessary to formulate general principles that apply widely across species and mountain systems. Further experimental approaches as the one used in this study might overcome the lack of evidence of woody invasion in mountains and contribute to comprehensive predictions.
References
Adams SN (1975) Sheep and cattle grazing in forests: a review. J Appl Ecol 12(1):143–152. https://doi.org/10.2307/2401724
Alba C, Skálová H, McGregor KF, D’Antonio C, Pyšek P (2015) Native and exotic plant species respond differently to wildfire and prescribed fire as revealed by meta-analysis. J Veg Sci 26(1):102–113. https://doi.org/10.1111/jvs.12212
Alexander JM, Lembrechts JJ, Cavieres LA, Daehler C, Haider S, Kueffer C, Liu G, McDougall K, Milbau A, Pauchard A, Rew LJ, Seipel T (2016) Plant invasions into mountains and alpine ecosystems: current status and future challenges. Alp Bot 126(2):89–103. https://doi.org/10.1007/s00035-016-0172-8
Ansari S, Daehler DD (2010) Life history variation in a temperate plant invader, Verbascum thapsus along a tropical elevational gradient in Hawaii. Biol Invasions 12(12):4033–4047. https://doi.org/10.1007/s10530-010-9810-z
Argañaraz JP, Pizarro GG, Zak M, Bellis LM (2015) Fire regime, climate, and vegetation in the Sierras de Córdoba, Argentina. Fire Ecol 11(1):55–73. https://doi.org/10.4996/fireecology.1101055
Arnan X, Rodrigo A, Retana J (2007) Uncoupling the effects of shade and food resources of vegetation on mediterranean ants: an experimental approach at the community level. Ecography 30(2):161–172. https://doi.org/10.1111/j.0906-7590.2007.04796.x
Callaway RM, Brooker RW, Choler P, Kikvidze Z, Lortie CJ, Michalet R, Paolini L, Pugnaire FI, Newingham B, Aschehoug ET, Armas C, Kikodze D, Cook BJ (2002) Positive interactions among alpine plants increase with stress. Nature 417(6891):844–848. https://doi.org/10.1038/nature00812
Capó EA, Aguilar R, Renison D (2016) Livestock reduces juvenile tree growth of alien invasive species with a minimal effect on natives: a field experiment using exclosures. Biol Invasions 18(10):2943–2950. https://doi.org/10.1007/s10530-016-1185-3
Certini G (2005) Effects of fire on properties of forest soils: a review. Oecologia 143:1–10. https://doi.org/10.1007/s00442-004-1788-8
Chagnon PL, Bradley RL, Maherali H, Klironomos JN (2013) A trait-based framework to understand life history of mycorrhizal fungi. Trends Plant Sci 9:484–491. https://doi.org/10.1016/j.tplants.2013.05.001
Chaneton EJ, Mazía J, Machera M, Uchitel A, Ghersa CM (2004) Establishment of Honey Locust (Gleditsia triacanthos) in burned pampean grasslands. Weed Technol 18(1):1325–1329. https://doi.org/10.1614/0890-037X(2004)018[1325:EOHLGT]2.0.CO;2
Cingolani AM, Cabido MR, Renison D, Solís Neffa V (2003) Combined effects of environment and grazing on vegetation structure in Argentine granite grasslands. J Veg Sci 14(2):223–232. https://doi.org/10.1016/j.rse.2004.05.008
Cingolani AM, Renison D, Zak M, Cabido M (2004) Mapping vegetation in a heterogeneous mountain rangeland using Landsat data: an alternative method to define and classify land-cover units. Remote Sens Environ 92(1):84–97. https://doi.org/10.1111/j.1654-1103.2003.tb02147.x
Colladon L, Felici GS, Pazos I. (2010) Anuario pluviométrico 2005 -2010. Cuenca del río San Antonio: Sistema del Río Suquía, Provincia de Córdoba. 1st edn. Instituto Nacional del Agua, Villa Carlos Paz
D’Antonio CM, Hughes RF, Vitousek PM (2001) Factors influencing dynamics of two invasive C4 grasses in seasonally dry Hawaiian woodlands. Ecology 82(1):89–104. https://doi.org/10.1890/0012-9658(2001)082[0089:FIDOTI]2.0.CO;2
De Steven D (1991) Experiments on mechanisms of tree establishment in old-field succession: seedling emergence. Ecology 72(3):1066–1075. https://doi.org/10.2307/1940606
Dietz H, Edwards JP (2006) Recognition that causal processes change during plant invasion helps explain conflicts in evidence. Ecology 87:1359–1367. https://doi.org/10.1890/0012-9658(2006)87[1359:RTCPCD]2.0.CO;2
Fernández RD, Ceballos SJ, Malizia A, Aragón R (2017) Gleditsia triacanthos (Fabaceae) in Argentina: a review of its invasion. Aust J Bot 65:203–213. https://doi.org/10.1071/BT16147
Ferreras AE, Galetto L (2010) From seed production to seedling establishment: important steps in an invasive process. Acta Oecol 36(2):211–218. https://doi.org/10.1016/j.actao.2009.12.005
Ferreras AE, Funes G, Galetto L (2015) The role of seed germination in the invasion process of Honey locust (Gleditsia triacanthos L., Fabaceae): comparison with a native confamilial. Plant Species Biol 30(2):126–136. https://doi.org/10.1111/1442-1984.12041
Gai JP, Tian H, Yang FY, Christie P, Li XL, Klironomos JN (2012) Arbuscular mycorrhizal fungal diversity along a Tibetan elevation gradient. Pedobiol 55:145–151. https://doi.org/10.1016/j.pedobi.2011.12.004
Gehring CA, Whitham TG (1994) Interactions between aboveground herbivores and the mycorrhizal mutualists of plants. Trends Ecol Evol 9(7):251–255. https://doi.org/10.1016/0169-5347(94)90290-9
Gimeno-García E, Andreu V, Rubio JL (2000) Changes in organic matter, nitrogen, phosphorus and cations in soil as a result of fire and water erosion in a Mediterranean landscape. Eur J Soil Sci 51(2):201–210. https://doi.org/10.1046/j.1365-2389.2000.00310.x
Giorgis M, Tecco PA (2014) Árboles y arbustos invasores de la provincia de Córdoba (Argentina): una contribución a la sistematización de bases de datos globales. B Soc Argent Bot 49(4):581–603
Giorgis MA, Tecco PA, Cingolani AM, Renison D, Marcora PI, Paiaro V (2011a) Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina. Biol Invasions 13(6):1423–1434. https://doi.org/10.1007/s11258-009-9674-4
Giorgis MA, Cingolani AM, Chiarini F, Chiapella J, Barboza G, Espinar LA, Moreno R, Gurvich DE, Tecco PA, Subils R, Cabido M (2011b) Composición florística del bosque chaqueño serrano de la provincia de Córdoba. Argentina. Kurtziana 36(1):9–43
Giorgis MA, Cingolani AM, Tecco PA, Cabido MR, Poca M, von Wehrden H (2016) Testing alien plant distribution and habitat invasibility in mountain ecosystems: growth form matters. Biol Invasions 18(7):2017–2028. https://doi.org/10.1007/s10530-016-1148-8
Grace C, Stribley DP (1991) A safer procedure for routine staining of vesicular-arbuscular mycorrhizal fungi. Mycol Res 95(10):1160–1162. https://doi.org/10.1016/S0953-7562(09)80005-1
Grümberg BC, Urcelay C, Shroeder MA, Vargas-Gil S, Luna MC (2015) The role of inoculums identity in drought stress mitigation by arbuscular mycorrhizal fungi in soybean. Biol Fertil Soils 51(1):1–10. https://doi.org/10.1007/s00374-014-0942-7
Gundale MJ, Kardol P, Nilsson MC, Nilsson U, Lucas RW, Wardle DA (2014) Interactions with soil biota shift from negative to positive when a tree species is moved outside its native range. New Phytol 202(2):415–421. https://doi.org/10.1111/nph.12699
Herrero ML, Torres R, Renison D (2016) Do wildfires promote woody species invasion in a fire-adapted ecosystem? Post-fire resprouting of native and non-native woody plants in central Argentina. Environ Manage 57(2):308–317. https://doi.org/10.1007/s00267-015-0616-8
Jaureguiberry P, Díaz S (2015) Post-burning regeneration of the chaco seasonally dry forest: germination response of dominant species to experimental heatshock. Oecologia 177(3):689–699. https://doi.org/10.1007/s00442-014-3161-x
Kimball S, Schiffman P (2003) Differing effects of cattle grazing on native and alien plants. Conserv Biol 17(6):1681–1693. https://doi.org/10.1111/j.1523-1739.2003.00205.x
Körner C (1999) Alpine plant life. Springer, Berlin
Kozlowski TT (1999) Soil compaction and growth of woody plants. Scand J For Res 14:596–619
Larson DL, Anderson PJ, Newton W (2001) Alien plant invasion in mixed-grass prairie: effects of vegetation type and anthropogenic disturbance. Ecol App 11(1):128–141. https://doi.org/10.1890/1051-0761(2001)011%5B0128:APIIMG%5D2.0.CO;2
Lembrechts JJ, Milbau A, Nijs I (2015) Trade-off between competition and facilitation defines gap colonization in mountains. AoB Plants 7:1–13. https://doi.org/10.1093/aobpla/plv128 (plv128)
Lembrechts JJ, Pauchard A, Lenoir J, Nuñez MA, Geron C, Ven A, Bravo-Monasterio P, Teneb E, Nijs I, Milbau A (2016) Disturbance is the key to plant invasions in cold environments. Proc Natl Acad Sci USA 113(49):14061–14066. https://doi.org/10.1073/pnas.1608980113
Little EL Jr (1971) Conifers and important hardwoods. In: Atlas of United States trees, vol 1. 1st edn. US department of agriculture, Washington
Longo S, Nouhra E, Goto BT, Berbara RL, Urcelay C (2014) Effects of fire on arbuscular mycorrhizal fungi in the mountain chaco forest. For Ecol Manag 315:86–94. https://doi.org/10.1016/j.foreco.2013.12.027
Longo S, Cofré N, Soteras F, Grilli G, Lugo M, Urcelay C (2016) Taxonomic and functional response of arbuscular mycorrhizal fungi to land use change in central Argentina. In: Pagano MC (ed) Recent advances on mycorrhizal fungi. Springer, Cham, pp 81–90. https://doi.org/10.1007/978-3-319-24355-9_7
Lonsdale WM (1999) Global patterns of plant invasions and the concept of invasibility. Ecology 80(5):1522–1536. https://doi.org/10.1890/0012-9658(1999)080[1522:GPOPIA]2.0.CO;2
Marcora PI, Hensen I, Renison D, Seltmann P, Wesche K (2008) The performance of Polylepis australis trees along their entire altitudinal range: implications of climate change for their conservation. Divers Distributions 14(4):630–636. https://doi.org/10.1111/j.1472-4642.2007.00455.x
Marcora PI, Renison D, Pais-Bosch AI, Cabido M, Tecco PA (2013) The effect of altitude and grazing on seedling establishment of woody species in central Argentina. For Ecol Manag 291:300–307. https://doi.org/10.1016/j.foreco.2012.11.030
Mazía NC, Chaneton EJ, Ghersa CM, León RJ (2001) Limits to tree species invasion in pampean grassland and forest plant communities. Oecologia 128(4):594–602. https://doi.org/10.1007/s004420100709
Mazía CN, Chaneton EJ, Machera M, Uchitel A, Feler MV, Ghersa CM (2010) Antagonistic effects of large-and small-scale disturbances on exotic tree invasion in a native tussock grassland relict. Biol Invasions 12(9):3109–3122. https://doi.org/10.1007/s10530-010-9702-2
McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA (1990) A new method which gives an objective measure of colonization of roots by vesicular—arbuscular mycorrhizal fungi. New Phytol 115(3):495–501. https://doi.org/10.1111/j.1469-8137.1990.tb00476.x
Milbau A, Shevtsova A, Osler N, Mooshammer M, Graae B (2013) Plant community type and small-scale disturbances, but not altitude, influence the invisibility in subarctic ecosystems. New Phytol 197(3):1002–1011
Millennium Ecosystem Assessment (2005) Millennium Ecosystem Assessment Ecosystems and human well-being: Biodiversity synthesis. World Resources Institute, Washington
Muschetto E, Mazía N, Cueto GR, Busch M (2015) Are rodents a source of biotic resistance to tree invasion in Pampean grasslands? Tree seed consumption under different conditions. Aust Ecol 40(3):255–266. https://doi.org/10.1111/aec.12208
van der Heyde M, Ohsowski BM, Abbott LK, Hart M (2017) Arbuscular mycorrhizal fungus responses to disturbance are context-dependent. Mycorrhiza 27:431–440
Nuñez MA, Dickie IA (2014) Invasive belowground mutualists of woody plants. Biol Invasions 16(3):645–661. https://doi.org/10.1007/s10530-013-0612-y
Ordóñez JL, Retana J (2004) Early reduction of post-fire recruitment of Pinus nigra by post-dispersal seed predation in different time-since-fire habitats. Ecography 27(4):449–458. https://doi.org/10.1111/j.0906-7590.2004.03886.x
Pauchard A, Kueffer C, Dietz H, Daehler CC, Alexander J, Edwards PJ, Arévalo JR, Cavieres LA, Guisan A, Haider S, Jakobs G, McDougall K, Millar CI, Naylor BJ, Parks CG, Rew LJ, Seipel T (2009) Ain’t no mountain high enough: plant invasions reaching new elevations. Front Ecol Environ 7(9):479–486. https://doi.org/10.1890/080072
Poll M, Naylor BJ, Alexander JM, Edwards PJ, Dietz H (2009) Seedling establishment of Asteraceae forbs along altitudinal gradients: a comparison of transplant experiments in the native and introduced ranges. Divers Distributions 15(2):254–265. https://doi.org/10.1111/j.1472-4642.2008.00540.x
Pyšek P, Jarošik V, Pergl J, Wild J (2011) Colonization of high altitudes by alien plants over the last two centuries. PNAS 108(2):439–440
Quiroz C, Cavieres L, Pauchard A (2011) Assessing the importance of disturbance, site conditions, and the biotic barrier for dandelion invasion in an Alpine habitat. Biol Invasions 13:2889–2899. https://doi.org/10.1007/s10530-011-9971-4
Rejmánek M, Richardson DM (2013) Trees and shrubs as invasive alien species—2013 update of the global database. Divers Distributions 19(8):1093–1094. https://doi.org/10.1111/ddi.12075
Relva MA, Nuñez M, Simberloff D (2010) Introduced deer reduced native plant cover and facilitate invasion of non-native tree species: evidence for invasional meltdown. Biol Invasions 12:303–311. https://doi.org/10.1007/s10530-009-9623-0
Richardson DM, Rejmánek M (2011) Trees and shrubs as invasive alien species—a global review. Divers Distributions 17(5):788–809. https://doi.org/10.1111/j.1472-4642.2011.00782.x
Richardson DM, Hui C, Nuñez MA, Pauchard A (2014) Tree invasions: patterns, processes, challenges and opportunities. Biol Invasions 16(3):473–481. https://doi.org/10.1007/s10530-013-0606-9
Smith S, Read D (2008) Colonization of roots and anatomy of arbuscular mycorrhiza. Mycorrhizal Symbiosis. Academic Press, London
Stevens JT, Latimer AM (2015) Snowpack, fire, and forest disturbance: interactions affect montane invasions by non-native shrubs. Glob Change Biol 21(6):2379–2393. https://doi.org/10.1111/gcb.12824
te Beest M, Mpandza NJ, Olff H (2015) Fire and simulated herbivory have antagonistic effects on resistance of savanna grasslands to alien shrub invasion. J Veg Sci 26(1):114–122. https://doi.org/10.1111/jvs.12208
R Development Core Team (2016) R: A language and environment for statistical computing. Foundation for statistical computing. Vienna, Austria. http://www.R-roject.org. Accessed 17 Mar 2016
Tecco PA, Urcelay C, Días S, Cabido M, Pérez-Harguindeguy N (2013) Contrasting functional trait sindromes underlay woody alien success in the same ecosystem. Aust Ecol 38(4):443–451. https://doi.org/10.1111/j.1442-9993.2012.02428.x
Tecco PA, Pais-Bosch AI, Funes G, Marcora PI, Zeballos SR, Cabido M, Urcelay C (2016) Mountain invasions on the way: are there climatic constraints for the expansion of alien woody species along an elevation gradient in Argentina? J Plant Ecol 9(4):380–392. https://doi.org/10.1093/jpe/rtv064
Tjelele J, Ward D, Dziba L (2015) The effects of seed ingestion by livestock, dung fertilization, trampling, grass competition and fire on seedling establishment of two woody plant species. PLoS One 10(2):e0117788. https://doi.org/10.1371/journal.pone.0117788
Torres RC, Giorgis MA, Trillo C, Volkmann L, Demaio P, Heredia J, Renison D (2014) Post-fire recovery occurs overwhelmingly by resprouting in the chaco serrano forest of central Argentina. Austral Ecol 39(3):346–354. https://doi.org/10.1111/aec.12084
Urcelay C, Longo S, Geml J, Tecco PA, Nouhra E (2017) Co-invasive exotic pines and their ectomycorrhizal symbionts show capabilities for wide distance and altitudinal range expansion. Fungal Ecol 25:50–58. https://doi.org/10.1016/j.funeco.2016.11.002
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M (2000) Global biodiversity scenarios for the year 2100. Science 287(5459):1770–1774. https://doi.org/10.1126/science.287.5459.1770
Zeballos SR, Tecco PA, Cabido M, Gurvich DE (2014) Composición de especies leñosas en comunidades invadidas en montañas del centro de Argentina: su relación con factores ambientales locales. Rev Biol Trop 62(4):1673–1681
Acknowledgements
Authors acknowledge the assistance of Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), and Secretaría de Ciencia y Técnica (SECyT) of Universidad Nacional de Córdoba, both of which supported this investigation. We also thank M. Cabido for promoting this project, E. Kowaljow for field assistance, M. Pereyra for identification of ant species and J. Brasca for English edition of this article. We especially thank land owners (J. Astrada, Atum Pampa, Olguín), who provided access to the study site. The authors specially acknowledge the suggestions of the editor and two anonymous reviewers that improved the article.
Author information
Authors and Affiliations
Contributions
MP, FA and TP conceived the ideas and designed methodology; MP, FA, TP, FG and ZS collected the data; UC and LS measured mycorrhizal colonization; MP, FA, TP and ZS analyzed the data; all authors were involved in the writing of the manuscript, contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Disclosure of potential conflicts of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Edith B. Allen.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Marcora, P.I., Ferreras, A.E., Zeballos, S.R. et al. Context-dependent effects of fire and browsing on woody alien invasion in mountain ecosystems. Oecologia 188, 479–490 (2018). https://doi.org/10.1007/s00442-018-4227-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-018-4227-y