
Abstract
The phenology of avian migration appears to be changing in response to climate change. Seemingly contradictory differences in the timing of these annual cycles have been reported in published studies. We show that differences between studies in the choice of songbird species, as well as in the measurements of migration phenology, can explain most of the reported differences. Furthermore, while earlier spring arrival is evident across these studies, trends in timing of departure show large variation between species and according to individual timing of migration (early-arriving vs. late-departing individuals). Much of the variation in departure between species could be explained by each species’ migratory status. We present a detailed analysis of migrants recorded at a Danish migration site, and reveal that although shifts in migration timing can be demonstrated for almost all species, these shifts are either most pronounced in the early arriving/late departing individuals or the changes are similar. Thus most individuals do not seem to change their breeding-area residence time (BART). As BART is likely to reflect ecologically important factors, e.g. number of clutches, we expect that only small effects have been exerted on the breeding ecology of the studied species in the time period investigated.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Large numbers of avian species migrate annually between their northerly breeding areas and tropical wintering regions (Moreau 1972; Rappole 1995), with an estimated 5,000 million birds migrating to Africa from the Palearctic ecozone alone (Moreau 1972). Pronounced decreases in many populations of long-distance migratory songbirds have been recorded on both sides of the Atlantic (see e.g. Berthold 1991; Rappole 1995). However, elucidating the causes of these decreases is a complex task, making targeted conservation action difficult. For example, accelerating climatic changes appear to affect the timing of seasonal events (e.g. Easterling et al. 1997), which in turn may have important consequences for population dynamics (Hughes 2000; Walther et al. 2002). Also, while we are already witnessing earlier spring arrivals among migratory birds (e.g. Bairlein and Winkel 2001; Hüppop and Hüppop 2003; Marra et al. 2005; Tøttrup et al. 2006), both advanced as well as delayed autumn departures have been reported for mixed groups of migrants with various migration strategies (e.g. Bairlein and Winkel 2001; Cotton 2003; Jenni and Kéry 2003). Similarly, reported changes in the duration of breeding-area residence time (BART), a factor considered to be more directly linked to ecological performance than migration timing, are not consistent (e.g. Bairlein and Winkel 2001; Cotton 2003; Jenni and Kéry 2003; Lehikoinen et al. 2004).
Here, we compare trends in the timing of migration of European songbirds across published studies, encompassing temporal changes derived from data from the UK, Germany and Switzerland (Cotton 2003; Hüppop and Hüppop 2003; Jenni and Kéry 2003), with trends from a Danish data set. Each of these published studies is based on a distinct subset of species, with each subset having different migration strategies. In addition, the time periods considered, as well as the measures of population arrival, differ between these studies. By selectively comparing species common to these studies, we intend to determine whether trends in timing of migration among European songbirds are similar between sites. Since BART durations are correlated to biologically important variables, such as the number of clutches in 1 year or the time period available to feed young or moult, they are suitable for testing the hypothesis that climate changes may lead to extended reproductive periods on breeding grounds. Accordingly, this hypothesis is tested by identifying differences in BART trends among insectivorous long- and short-distance migrants from primarily Fenno-Scandian and Finnish breeding populations, counted at Christiansø, an island in the Baltic Sea along a major migration route. In addition, we investigate whether there are differences between subsets within each population with respect to migration timing (earliest arriving and last departing individuals), by focusing on BART as a measure of the duration of population subsets present in the breeding area, as well as changes in BART over time. Finally, we discuss ecological implications that these changes might have on breeding strategies, as well as future population survival.
Materials and methods
Comparing trends between studies
The data sets used for comparing trends in migration timing of migratory European songbirds were obtained from the UK (Cotton 2003), Germany (Hüppop and Hüppop 2003), Switzerland (Jenni and Kéry 2003) and Denmark (Tøttrup et al. 2006, in press; this study), and cover a time span of three decades (ca. 1970–2000). Only insectivorous songbirds occurred in common between any two sites. The data from the UK, Germany and Switzerland only included one population measure (i.e. median or first/last individual), and none were from the same season. Trends calculated for each species, population measure, country and season from these data are compared with similar trends calculated from the Danish data set covering the period 1976–1997. To make pair-wise comparisons between study sites, we compare mean trends of species with the same migratory behaviour between studies using the same population measures, over approximately the same time period (approximately 1970–2000), including only species common to the studies compared (see Table S1 in ESM). Differences between mean trends of species with the same migratory behaviour are modelled using an ANOVA on the interaction between the categorical variables population measure (median, first/last), migratory distance category (long, short distance) and season (spring, autumn). The effect of Country is tested by including an additional interaction with Country (the UK, Germany, Switzerland, Denmark),
The Swiss data set, published by Jenni and Kéry (2003), considers the estimated peak autumn passage of migration at Col de Bretolet as the mean of the three periods: 1958–1969, 1970–1982 and 1988–1999. In their original report, a relatively large number of short-distance seed-eaters were included, compared to other studies. We use the trend between the two periods 1970–1982 and 1988–1999, i.e. a span of 22 years. The German data set published by Hüppop and Hüppop (2003) considers mean timing of spring migration at Helgoland for the period 1960–2000. Additional data on autumn migration at Helgoland are published by Hüppop and Hüppop (2005). However, these autumn data were not included in the analysis since they had a smaller number of species in common with the Danish data set. The data set from the UK published by Cotton (2003) considers first arrival in spring and last departure in autumn as measures of migration timing in Oxfordshire, for the period 1971–2000.
Danish data
The Danish data are derived from the standardised mist-netting of more than 568,000 individual birds covering 184 species in the period 1976–1997 on the Danish island of Christiansø (55°19′N, 15°11′E) in the Baltic Sea. The 1979 and 1980 data sets are excluded, due to non-continuous coverage during the autumn season (Rabøl and Rahbek 2002). The captured migrants have their main breeding areas in Sweden and Finland, and pass the isolated island of Christiansø biannually during their spring and autumn migration.
Four measures of population migration timing are calculated for each season and species, these being dates of first (spring) arriving or last (autumn) departing birds, as well as the day (in Julian days where day 1 = 1 January) when 5, 50 and 95%, respectively, of the total number of individuals in a season had been trapped (Fig. 1). For each species, the value of the slope of the least square regression is used to describe phenological change over time. We only included a seasonal measure for a species if visual inspection of the cumulative arrival (or departure) showed the migration period not to extend beyond the dates covered by mist-netting activities and was not affected by summering or wintering birds. The methods and procedures are described in more detail in Tøttrup et al. (2006).

Measures of timing of migration, illustrated with the song thrush Turdus philomelos’ migration during 1985 at Christiansø, Denmark. For each season we calculated trapping dates when 50% of the season total remained to be trapped. Breeding-area residence time (BART) is calculated as the difference between autumn and spring measures [earliest arriving and last departing individual, and 50% of the total number of trapped individuals in a season (spring/autumn, respectively)]. Similar measures were calculated for 5 and 95% quantiles. These are not shown on the figure to ease interpretation. Mar March, Apr April, Jun June, Jul July, Aug August, Sep September, Oct October, Nov November
Detailed analysis of the Danish data set
For each species, BART was calculated as the difference between autumn departure and spring arrival dates. We calculated four measures of BART corresponding to the four measures of population timing described above: total (first/last), as well as 5, 50 (median) and 95% BART. These estimates of changes over time are presented in the ESM and figures for ease of interpretation.
To test for differences in trends among BART durations between migratory categories (short- vs. long-distance migrants) as well as between different measures of BART, we use general linear models with Yearly Arrival as the dependent variable and Year as the main predictor variable of interest. Parameter estimates throughout the text are based on this statistical model. These estimates may differ somewhat from those obtained using values of slopes averaged over species.
First, we tested for differences between two essentially different measures of BART (total and 50%). Subsequently, we analysed changes in the three measures of BART based on quantiles (5, 50 and 95%) separately from the analysis including the total BART. Contrary to the total BART measure, these quantile measures are not influenced by trends in capture effort, and a stronger autocorrelation is likely to arise between these measures than through the use of the first arrival/last departure measure.
Due to likely autocorrelation between the measures of BART, individual data points cannot be considered to be independent, and as a result of this the analysis of temporal changes is complemented with an analysis using: (1) ordinary least square regression applied to the difference between measures of BART (e.g. 50% BART minus 5% BART), and (2) repeated measures multivariate approaches (cf. Tøttrup et al. 2006). These analyses are used to confirm the significance of differences between measures of BART. The results of repeated measures multivariate approaches are very similar to the results of ordinary least square regressions. Here, we only present the results of ordinary least square regressions.
Temporal changes are tested using full statistical models, and include Year as a predictor variable. In models with more than three parameters, we use backward elimination of parameters with probabilities >0.05 to identify models for inference. All analyses in this paper were generated using SAS software (version 8.2).
Results
Residence time unchanged
When restricting the analysis of the four European studies to a comparable subset of data containing only the species in common among the study sites, similar trends for each species group (short- and long-distance migrants) and measure of timing (early/late and median) over the three-decade time span become apparent. There are differences in trends between measures, migratory distance categories and seasons (P < 0.0001, F = 15.2, df = 7; Fig. 2), while no differences between countries where observed (P = 0.46, F = 0.95, df = 6). Spring arrivals were generally seen to be earlier for all species, most noticeably for the earliest arriving individual of short-distance migrants (Fig. 2, lower half). In contrast, autumn departure could be seen to vary more between group and population subsets, leading to more varied residence times (Fig. 2, upper half). The overall picture of first arrivals and last departures is that of a generally small increase in residence time for the mixed group of sub-Saharan migrants, while the residence time for the group of short-distance migrants is considerably more extended. Median residence time increased slightly for sub-Saharan migrants, while decreasing for short-distance migrants.

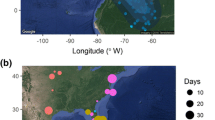
Comparison of results from four, geographically diverse European studies covering changes over three decades (ca. 1970–2000) in arrival and departure time for different population subsets of migratory, insectivorous European songbirds. Trend lines indicate the change in arrival and departure time for different population subsets in spring and autumn. The lines have been parallel displaced to cross the x-axis at equal points for each group. The colours of the trend lines indicate which site data are from (shown on the map). The comparison is carried out on data collected from England (Cotton 2003), Switzerland (Jenny and Kéry 2003), Germany (Hüppop and Hüppop 2003), and Denmark (Tøttrup et al. 2006, in press). For Germany and Switzerland, means and peaks, respectively, are shown instead of medians
The significance of the results in Fig. 2 is elucidated by a more detailed analysis of data from the Danish migration study site, situated close to the breeding grounds of the migratory populations. Here, the average total BART (first arrival to last departure) for short-distance migrants was 230 days (n = 7, SE = 2) and 161 days (n = 7, SE = 2) for long-distance migrants. The average 50% BART for short-distance migrants was 177 days (n = 8, SE = 2) and for long-distance migrants 105 days (n = 14, SE = 2).
The Danish study
Based on the species-specific trends for earliest arriving and last departing individuals, there is a substantial change in durations of total BART for short-distance migrants, while differences in BART durations are considerably less for long-distance migrants (Fig. 3). The other measures of timing (5, 50 and 95% BART measures, respectively) show smaller changes, with the largest changes being for the 95% quantile of long-distance migrants (Fig. 3). Using a general linear model with Yearly Arrival/Departure as the dependent variable, and Year as main predictor variable, we find an overall difference in the migration timing trends between migration distance categories and population parts (significant Year×Distance category interaction in the model, with differences between BART measures as the dependent variable; F = 15.6, df = 1, P < 0.0001). A comparison of BART durations for earliest arriving and last departing individuals among short- and long-distance migrants reveals an increase in duration for short-distance migrants (0.68 days year−1, SE = 0.34, t = 2.00, P = 0.046), whereas smaller changes are apparent for sub-Saharan migrants (−0.08 days year−1, SE = 0.20, t = −0.39, P = 0.69; see also Fig. 3). This temporal difference is not evident when BART is estimated based on 50% of the passing migrating population (Fig. 3). However, a tendency toward reversed changes in the time spent on the breeding grounds for the 50% BART measure is apparent, with 50% BART being largely unchanged for short-distance migrants (−0.20 days year−1, SE = 0.34, t = −0.59, P = 0.56), while long-distance migrants exhibit extended 50% BART (0.21 days year−1, SE = 0.20, t = 1.01, P = 0.31; see also Fig. 3).

Mean temporal changes in BART (±SD) for different population subsets of migratory, insectivorous European songbirds (see Table S1 in ESM for details). Based on data from migrants mist netted during the period 1976–1997 at Christiansø, Denmark. Species are categorised according to timing of arrival/departure
The observed BART durations were not symmetrical around the 50% quantile (the median), i.e. there was a difference between 5/50 and 50/95% BART measures (P < 0.0001, F = 16.89, df = 1). For the three measures based on 5, 50 and 95% quantiles, the trends differed between migration-distance categories and quantiles (significant Year×Distance category interaction in the model, with differences between BART measures as dependent variable; F = 11.85, df = 2, P < 0.0001). Overall, BART has generally become extended for the majority of the long-distance migrants (50 and 95%; 0.21 days year−1, SE = 0.17, t = 1.20, P = 0.23 and 0.32 days year−1, SE = 0.24, t = 1.33, P = 0.18, respectively), whereas far less change is evident for short-distance migrants (<0.17 days year−1; see also Fig. 3).
Discussion
Our analysis of a subset of the published passage time data containing only the species in common among the study sites reveals that insectivorous songbirds do in fact show similar trends among sites (Fig. 2). Accordingly, spring arrival can be seen to be progressively earlier, whereas the timing of autumn departure varies between group and population subsets. Thus, the earlier reported differences can be assumed to be the result of varying time scales, species groups, as well as indicators of migration timing used in these studies.
Differences in the timing of autumn departure are indicative of varying durations of BART. The largest change observed in BART durations on the Danish migration site is the considerable extension in the duration of short-distance migrants in the breeding area (first arrival to last departure), while substantially smaller changes in BART durations are evident among sub-Saharan migrants. Earliest arriving and last departing individuals do not necessarily reflect the timing of the entire population, and many of these individuals could be abnormal birds that are unable to reproduce properly. Accordingly, the median residence time can be seen to change much less over time, and it may be assumed that most birds probably follow an advanced annual schedule, with little change in the time spent at the breeding grounds.
Using ringing data in order to deduce the migration timing for the population, our analyses rely on the assumption that the samples are representative for the population. Ringing data collected during autumn migration is dominated by large numbers of first-year birds, while in the spring young birds may also be over-represented in data sampling (Rabøl and Rahbek 2002). Sex- or age-differentiated migration strategies within migratory species have been also been documented (e.g. Ottosen et al. 2002; Catry et al. 2004, 2005). However, such intraspecific differences are generally smaller than interspecific differences and it seems likely that the large-scale changes reported here also apply to the actual breeding population. In considering the timing of different population parts (quantiles), we also assume that the migration timing of each bird reflects its breeding strategy, and that random variation (around a common mean) is less than variation due to the different population parts (a factor which we also take into account in this paper). Studies showing that individuals within a population follow different strategies in migration timing (e.g. Bêty et al. 2004), heritability in migration timing (Berthold and Pulido 1994) as well as fitness consequences related to arrival (e.g. Lozano et al. 1996) support this view. A confounding factor could also be that some of the data on some species could involve more than one population. However, at least for the Danish data set this is presumably only the case for very few species. There is little evidence regarding how timing in spring is related to timing in autumn for individual birds, i.e. whether the early-arriving birds are those leaving late. Since presence on the breeding grounds is likely to be important for keeping good territories and early-arriving species are generally also those leaving late, it may not be unreasonable to assume such a relationship. However, this is not implicit in our BART analysis, which is concerned with the time the population is present on the breeding grounds.
It is generally acknowledged that early arrival at breeding grounds is crucial for successful reproduction. Even though the first (and presumably most fit) migrants arrive earlier, both long- and short-distance migrants show a similar change in autumn departure, resulting in unchanged duration of reproduction. However, even though the first and presumably most fit individuals of short-distance migrants arrive earlier (Kokko 1999; Smith and Moore 2005), both long- and short-distance migrants show a corresponding change in autumn departure, and the reproductive period for the vast majority of individuals remains unchanged (Fig. 3). From Helgoland, Hüppop and Hüppop (2005) reported the mean time spent north of the site to be significantly extended for both long- and short-distance migrants. At Helgoland, autumn departure has generally been stable for both groups. At Christiansø median departure has also been stable for long-distance migrants, whereas it has advanced slightly for short-distance migrants. This autumn departure pattern is similar to the pattern that Jenni and Kéry (2003) reported from Col de Bretolet for the songbird species occurring at Christiansø (but quite different from that reported for all species including species feeding predominantly on seeds).
Since the median BART only showed small changes, the impact of the phenological changes found at the Danish site is expected to be small. Nevertheless, a tendency toward an extension of the median and short-term residence time of long-distance migrant populations can be discerned. This indicates that late-arriving long-distance migrants may experience longer breeding periods. This could change the timing of annual events giving more time for e.g. moult, or ultimately to an increase in the number of offspring or brood numbers per season—a phenomenon normally pertaining strictly to short-distance migratory species (Jenni and Kéry 2003).
Three factors could be responsible for earlier spring arrival (Noordwijk 2003): (1) conditions in the wintering area, (2) speed of migration, and (3) conditions in the breeding area. The lack of significant change in BART for the majority of the populations indicates that changes in reproductive tactics are constrained by either: (1) the endogenous migration programme, or (2) environmental prerequisites in the wintering or breeding area or along the migratory route. The extended BART for late-arriving long-distance migrants suggests that these birds are time-constrained, and that some individuals are able to extend their breeding season.
The observed spring trends of earlier arrival to the breeding grounds correspond to a general increase in spring temperatures in temperate regions (Easterling et al. 1997). Autumn temperatures in Europe have not increased accordingly (Easterling et al. 1997), and only small changes in the timing of autumn migration are observed for all species. Together with the earlier arrival of long-distance migrants, it seems reasonable to assume that overall temperature increases in the breeding area will mean that more long-distance migrants will be able to start their migration in time. Earlier winter area arrival for the last departing long-distance migrants is indicated by the advanced autumn departure time (Fig. 2), which also shows unchanged total BART (first to last individual; Fig. 3). Since conditions in the stop-over sites immediately south of the Sahara become harsher soon after the rains which usually fall June–August (e.g. Ottosson et al. 2005), these may be restricting the extension of BART.
Generally, migration timing is found to be under strict genetic control in long-distance migrants, whereas variation in migration timing in short-distance migrants has been demonstrated to be dependent on phenotypic variation (Berthold 1996). A long-distance migrant, the pied flycatcher Ficedula hypoleuca, showed little change in arrival dates over time even though laying dates advanced (Both and Visser 2001). Long-distance migrants may be constrained in changing their timing in response to stronger climate change in the breeding area. We found a non-significant advancement of 0.16 days year−1 in this species. Hüppop and Hüppop (2005) found a significant change of the same magnitude at Helgoland. Additionally, we found overall significant changes in both groups of migrants (see Table S1 in ESM, Fig. 1) similar to the changes reported by Hüppop and Hüppop (2003), indicating that both groups, regardless of strategies, are able to adjust migration timing according to the weather/climate they experience before the onset of, and during, migration.
Many populations of long-distance migrants have experienced pronounced decreases in numbers (Berthold 1991; Terborgh 1992; Fuller et al. 1995; Rappole 1995; Siriwardena et al. 1998, but see also Rabøl and Rahbek 2002 for other trends in Fenno-Scandian populations). In contrast, temperate zone sedentary species as well as short-distance migrants may benefit from a warmer winter climate, due to decreased winter mortality and an increased number of successful offspring per brood (Bairlein and Winkel 2001). Given that all the songbird species included in this analysis are able to reproduce in their first summer, rapid effects of changes in fitness due to climate change can be expected to be seen in these species. However, correlating population indices at Christiansø (Rabøl and Rahbek 2002) with median BART did not reveal any significant effect of BART, but this would not have been expected due to the overall lack of change in median BART. Clearly, density-dependent factors influencing population sizes, as well as inter-specific interactions from increased competition and species expansions may prove to be important factors affecting the timing of the annual cycle of migratory birds. The identification of consistent patterns in migration timing, as revealed here for both short- and long-distance migratory species, may help to elucidate the implications and consequences of accelerating climate change on avian populations.
References
Bairlein F, Winkel W (2001) Birds and climate change. In: Lozan JL, Grassl H, Hupfer P (eds) Climate of the twenty first century: changes and risks. GEO, Hamburg, pp 278–282
Berthold P (1991) Patterns of avian migration in light of current global “greenhouse” effects: a central European perspective. Acta Congr Int Ornithol 22:780–786
Berthold P (1996) Control of bird migration. Chapman & Hall, London
Berthold P, Pulido F (1994) Heritability of migratory activity in a natural bird population. Proc R Soc Lond B 257:311–315
Bêty J, Giroux JF, Gauthier G (2004) Individual variation in timing of migration: causes and reproductive consequences in greater snow geese Anser caerulescens atlanticus. Behav Ecol Sociobiol 57:1–8
Both C, Visser ME (2001) Adjustment to climate change is constrained by arrival date a long-distance migrant bird. Nature 411:296–298
Catry P, Campos A, Almada V, Cresswell W (2004) Winter segregation of migrant European robins Erithacus rubecula in relation to sex, age and size. J Avian Biol 35:204–209
Catry P, Lecoq M, Araújo A, Conway G, Felgueiras M, King MB, Rumsey S, Salima H, Tenreiro P (2005) Differential migration of chiffchaffs Phylloscopus collybita and P. ibericus in Europe and Africa. J Avian Biol 36:184–190
Cotton PA (2003) Avian migration phenology and global climate change. PNAS 100:12219–12222
Easterling DR, Horton B, Jones PD, Peterson TC, Karl TR, Parker DE, Salinger MJ, Razuvayev V, Plummer N, Jamason P, Folland CK (1997) Maximum and minimum temperature trends for the globe. Science 277:364–367
Fuller RJ, Gregory RD, Gibbons DW, Marchant JH, Wilson JD, Baillie SR, Carter N (1995) Population declines and range contraction among lowland farmland birds in Britain. Conserv Biol 9:1425–1441
Hughes L (2000) Biological consequences of global warming: is the signal already apparent? Trends Ecol Evol 15:56–61
Hüppop O, Hüppop K (2003) North Atlantic Oscillation and timing of the spring migration in birds. Proc R Soc Lond B 270:233–240
Hüppop K, Hüppop O (2005) Atlas zur Vogelberingung auf Helgoland. Teil 3. Veränderungen von Heim- und Wegzugzeiten von 1960 bis 2001. Vogelwarte 43:217–248
Jenni L, Kéry M (2003) Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants. Proc R Soc Lond B 270:1467–1471
Kokko H (1999) Competition for early arrival in migratory birds. J Anim Ecol 68:940–950
Lehikoinen E, Sparks TH, Zalakevicius M (2004) Arrival and departure dates. In: Møller AP, Fiedler W, Berthold P (eds) Birds and climate change. Elsevier, Amsterdam, pp 1–31
Lozano GA, Perreault S, Lemon RE (1996) Age, arrival date and reproductive success of male American redstarts Setophaga ruticilla. J Avian Biol 27:164–170
Marra PP, Francis CM, Mulvihill RS, Moore FR (2005) The influence of climate on the timing and rate of spring bird migration. Oecologia 142:307–315
Moreau RE (1972) The Palaearctic-African bird migration system. Academic Press, London
Noordwijk A (2003) The earlier bird. Nature 422:29
Ottosson U, Bairlein F, Hjort C (2002) Migration patterns of Palaearctic Acrocephalus and Sylvia warblers in north-eastern Nigeria. Vogelwarte 41:249–262
Ottosson U, Waldenström J, Hjort C, McGregor R (2005) Garden Warbler Sylvia borin migration in sub-Saharan West Africa: phenology and body mass changes. Ibis 147:750–757
Rabøl J, Rahbek C (2002) Population trends in the Baltic passerine migrants, elucidated by a combination of ringing data and point- and summer-count indices. Dansk Orn Foren Tidsskr 96:15–38
Rappole J (1995) The ecology of migrant birds. Smithsonian Institution Press, Washington, D.C.
Siriwardena GM, Baillie SR, Buckland ST, Fewster RM, Marchant JH, Wilson JD (1998) Trends in abundance of farmland birds: a quantitative comparison of smoothed common birds census indices. J Appl Ecol 35:24–43
Smith RJ, Moore FR (2005) Arrival timing and seasonal reproductive performance in a long-distance migratory landbirds. Behav Ecol Sociobiol 57:231–239
Terborgh J (1992) Perspectives on the conservation of neotropical migrant landbirds. In: Hagan JM III, Johnston DW (eds) Ecology and conservation of Neotropical migrant landbirds. Smithsonian Institution Press, Washington, D.C.
Tøttrup AP, Thorup K, Rahbek C (2006) Patterns of change in timing of migration in North European songbird populations. J Avian Biol 37:84–92
Tøttrup AP, Thorup K, Rahbek C (2006) Changes in timing of autumn migration in north European songbird populations. Ardea (in press)
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJ, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395
Acknowledgements
We thank all the ringers working on Christiansø during the study period, as well as Christiansø Research Station. Special thanks are extended to Tom S. Romdal for comments on earlier drafts of this manuscript. The Danish National and Nature Agency and The National Environmental Research Institute, Department of Coastal Zone Ecology financed the collection of the data that made this paper possible. A. P. Tøttrup and C. Rahbek acknowledge the Danish National Science Foundation grant J. no. 21-03-0221 for their support of this research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Katrin Böhning-Gaese.
Electronic supplementary material
Rights and permissions
About this article
Cite this article
Thorup, K., Tøttrup, A.P. & Rahbek, C. Patterns of phenological changes in migratory birds. Oecologia 151, 697–703 (2007). https://doi.org/10.1007/s00442-006-0608-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-006-0608-8