
Abstract
Intraclutch variation in offspring size should evolve when offspring encounter unpredictable environmental conditions. This form of bet-hedging should maximise the lifetime reproductive success of individuals that engage it. We documented the numbers of eggs and means and variances of yolk volume in 15 frog species that occur in tropical savanna woodland. We experimentally determined the effects of initial yolk volume on larval growth patterns in four species. Intraclutch variation in yolk volume occurred to some degree in all species surveyed. Some species had very low, others had very high, intraclutch variation in yolk volume, but all species in which some clutches were highly variable also produced clutches with low variability. Species that occur in areas where the offspring environment is likely to be unpredictable had elevated levels of intraclutch variation in egg provisioning. There was no trade-off between egg size and number in any species surveyed. Under benign laboratory conditions, tadpoles from eggs with larger yolk volumes hatched at larger sizes, and these size differences persisted through a substantial proportion of the larval stage. This indicates that intraclutch variation in egg size has major offspring and thus parental fitness consequences, and is therefore a functional selection variable. This study provides evidence in support of models which predict that intraclutch variation in offspring provisioning can evolve in organisms that reproduce in unpredictable habitats.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Smith and Fretwell (1974) proposed a model of energy expenditure by parents on their offspring that considers the effects on parental fitness of offspring size, offspring fitness, resources invested and resources available. Their model predicts that for any simple, upwardly convex offspring fitness function, there is a single, optimal offspring size that maximises parental fitness, and therefore that for any combination of species and environment there is a single optimal level of investment in all young (see also Lloyd 1987; Winkler and Wallin 1987). These models do not account for the effect of environmental variability on offspring fitness (McGinley et al. 1987). This variation may lead to multiple fitness functions; when this occurs there is no single optimal offspring size. This suggests that these models are unlikely to apply to anurans that reproduce in temporary water bodies, since a major feature of these habitats is their great variability and unpredictability, both within and among years (Alford 1999).
It has been argued that divergence in propagule size within and among clutches is an expression of phenotypic variation allowing progeny to cope with environmental instabilities, and that this can lead to increased parental fitness (Capinera 1979; Kaplan and Cooper 1984). Highly variable and unpredictable offspring conditions should select for intraclutch variation in offspring size (McGinley et al. 1987; Einum and Fleming 2004). In unpredictable environments, large offspring should survive even under harsh environmental conditions, while all offspring can survive during ‘good’ years. This form of bet-hedging should maximise the lifetime reproductive success of individuals that engage in it, and be selected for under these conditions. One possible evolutionary response is within-clutch variation in egg size (Parichy and Kaplan 1992). McGinley et al. (1987) incorporated variation in habitat and offspring quality of several potential types into a model that predicts what strategy should occur under various conditions. Their model suggested that within-clutch variation in propagule size should be favoured by selection in either or both of conditions when offspring are relatively likely, but not certain, to encounter a high-quality environment, or conditions when variation among the environments they encounter is high. It may also occur if offspring and parental fitness is not a monotonically increasing function of investment into offspring (Kaplan 1992; Kaplan and Cooper 1984).
Although there is a substantial body of theory regarding how propagule size should evolve under various scenarios, there is relatively little documentation of how common variation in propagule size is, and how such variation affects the ecology, and ultimately the fitness, of offspring. Without this information, the strength and even the existence of selection differentials between propagules of different sizes remain speculative. The aims of this study were to address this lack of data by determining the extent to which variation in propagule size (egg yolk volume) occurs in an assemblage of anurans and documenting its consequences for rates of growth and development.
A number of studies have suggested that intraclutch variability in yolk volume in anurans is an adaptation to environmental variability. Crump (1981b) found that anuran species which breed in temporary as opposed to permanent ponds have higher variability in egg size within clutches and suggested that this pattern results from bet-hedging in response to environmental uncertainty in these temporary ponds (Crump 1984). Unpredictability in the larval environment is suggested as a reason for the high variability in egg size within clutches of Crinia (Ranidella) signifera (Williamson and Bull 1989, 1995) and Bombina orientalis (Kaplan 1992; Kaplan and King 1997). Tejedo and Reques (1992), however, attributed variation in egg size within clutches in Bufo calamita to maternal energetic conditions rather than the offspring environment. We tested the hypothesis that species that breed in less predictable environments will have higher intraclutch variation in yolk volume. We expected that species in the study area that are obligate burrowers should have elevated levels of intraclutch variation in yolk volume. These species all burrow to escape desiccation and are extremely explosive breeders, reproducing only after heavy rain, and can be found in drier more arid habitats (Barker et al. 1995; Cogger 2000) where the periods of rain or pond persistence can be uncertain.
Many anuran species have an aquatic larval stage. This biphasic lifecycle has advantages and disadvantages (Harris 1999; Slade and Wassersug 1975; Wassersug 1975). A biphasic life history allows offspring to exploit resources otherwise unavailable to adults and this allows a relatively small investment into each egg. The disadvantages are that offspring are vulnerable to a large suite of aquatic predators and competitors, and to the drying of temporary aquatic habitats (Alford 1999; Wassersug 1975). These advantages and disadvantages lead to trade-offs in levels of parental investment into eggs. Lower investment per egg allows the production of more eggs, while greater investment per egg hastens growth and development and reduces exposure to the hazards of the aquatic environment (Crump 1981a; Kaplan and Cooper 1984). We therefore expected a trade-off between egg size and number to exist, however we did not expect any relationships between intraclutch variation in yolk volume and mean egg size or clutch size since we predicted variation in offspring size to have evolved independently in response to the predictability of the larval environment.
Variation in yolk volume could be irrelevant to natural selection if it does not affect the fitness of offspring. Several of the studies that have documented variation have also examined its effects on rates and patterns of growth. Yolk volume can affect hatchling size and early development (Crump 1984; Tejedo and Reques 1992; Williamson and Bull 1989) but some of these studies also found that the effects of egg size on larvae disappeared by metamorphosis when larvae were reared in very-high-quality environments (Crump 1984; Tejedo and Reques 1992). When reared in low quality environments, larvae from larger-yolked eggs reached metamorphosis faster and at larger body sizes than those from smaller-yolked eggs (Kaplan 1992; Parichy and Kaplan 1992). In contrast, Berven and Chadra (1988) showed that larvae from small eggs had a longer larval period than larvae from large eggs, but in a high quality environment metamorphosed at a larger size than larvae from large eggs. We expected offspring from large eggs to be larger and have higher growth and development rates than offspring from small eggs from within the same clutch. In this case there would be fitness differentials between different sized offspring leading to parental fitness differentials between females with different levels of intraclutch variation in yolk volume. In more unpredictable offspring environments, such as those that obligate burrowing species in this study may experience, the bet-hedging strategy of having higher levels of intraclutch variation in yolk volume would be selected for.
Materials and methods
The anuran fauna of savannas in the Townsville region
Anurans that occur in the tropical savanna woodland areas west of Townsville, Australia (146°45′E 19°19′S), breed in seasonally intermittent water bodies and in perennial farm ponds that only occasionally dry. The temporary water bodies are extremely stochastic habitats existing only during the wet season and even then are prone to frequent drying. Although the perennial ponds only occasionally dry completely, water levels in both ponds and other temporary water bodies can fluctuate considerably thus affecting larval densities and resource availabilities. Twenty-three species breed in this area (M. Dziminski, personal observation). Townsville receives an average annual rainfall of 1108.5 mm and has mean annual maximum and minimum daily temperatures of 28.7°C and 19.7°C. The climate is characterized by marked wet and dry seasons, and rainfall is extremely episodic.
Patterns of intraclutch variation in yolk volume within and among species
Frogs were collected between September 1999 and April 2000 from 43 breeding sites, which were temporary and permanent farm ponds, temporary streams, borrow pits, and soakages. All were located in the lowland Alice River area (20–80 m elevation) and upland Hervey’s Range (300–400 m). Sites were visited on wet nights and amplectant pairs were gathered before oviposition to ensure all eggs were collected, placed in water in 20 l buckets with lids, and transported to the laboratory. Wet and dry bulb temperatures were recorded at each collection site to determine air temperature and relative humidity.
In the laboratory, each pair was placed in a separate plastic container (from 2 l to 20 l depending on the size of the species) containing aged tap water to a depth of 3–6 cm, and left until the morning to deposit eggs. The following morning clutches were removed to shallow plastic trays, spread out so that all eggs could be seen, and photographed to allow determination of the number of eggs per clutch. Samples of 80–800 eggs were then photographed backlit and at a higher magnification for the measurement of yolk volume. A millimetre scale was photographed at the same time as each clutch, so that measurements from the photographs could be calibrated to absolute units. A sample of 10–50 eggs per clutch was preserved in 5% formalin for determination of dry weights. These steps were completed before late cleavage (past the 16-cell stage) occurred. After this stage eggs become asymmetrical (Rugh 1951) and increase in size (Kaplan 1979). All eggs of each clutch were at the same stage of development when photographed. Parents were weighed and their snout–vent length (SVL) was measured. The parents and most of the clutch were returned to the collection site, usually on the first day after collection.
Negatives of egg photographs were scanned to digital images using a Polaroid SprintScan film scanner. The digital images were analysed using the computer graphics analysis program UTHSCSA ImageTool (Wilcox et al. 1995). The number of eggs per clutch was counted, and the area, maximum diameter and minimum diameter of the yolks of all viable eggs were measured from the higher magnification subsets of eggs, and converted to millimetres. Yolk volumes were estimated from the measured area using the formula:
where V = volume in cubic units and A = area in square units.
To ensure that this measurement of yolk volume was strongly related to egg provisioning, we examined the relationship between our estimation of yolk volume and the dry mass of yolk. Four each of the small, medium and large sized eggs from single clutches of three species that exhibit intraclutch variation in yolk volume (Cyclorana novaehollandiae, a burrowing hylid; Litoria nasuta, a ground hylid; and Limnodynastes ornatus, a burrowing myobatrachid) were selected arbitrarily from the samples preserved at the time the eggs were photographed. Each egg was assigned a number and individually digitally photographed to scale. The yolk volume of each egg was determined as outlined above; and each egg was then dried for 12 h at 70°C and weighed to 0.1 mg precision on an electronic analytical balance. Dry mass and estimated yolk volume are highly correlated in each species (regression analyses: C. novaehollandiae: slope=2.238, r 2=0.739, P<0.001, n=12; L. nasuta: slope=1.933, r 2=0.872, P<0.001, n=12; L. ornatus: slope=1.793, r 2=0.589, P<0.01, n=12). Therefore our estimate of yolk volume is an adequate measure of egg provisioning.
Number of eggs per clutch, mean yolk volume, the coefficient of variation (CV = standard deviation (SD)/mean) of yolk volume, and the ratio of the 95th to the 5th percentile of yolk volume were determined for each clutch. The two measures of intraclutch variability, the CV and the 95th to the 5th percentile ratio, are highly correlated (Pearson correlation: r=0.972, P<0.0001, n=154), so only the CV was used as a measure of variability in further analyses. We examined patterns of variability among genera by taking the mean for each genus of the maximum CV of yolk volume exhibited by clutches of each species in that genus. For genera in which we had data on only one species, we used the maximum CV for that species. The three clutch parameters we examined could be governed entirely by parental size, or more complex relationships might exist between them. To examine possible relationships, we carried out Spearman rank correlations for each species for which eight or more clutches were collected. Because apparent relationships between clutch parameters might be caused by their mutual correlations with parental body size, we also calculated partial correlations among the clutch parameters controlling for the effect of female mass. Female mass was chosen since it is an accurate measure of size and is highly correlated to SVL (Pearson correlation: r=0.928, P<0.0001, n=154). Finally we used one-way ANOVAs to determine whether CV, number of eggs per clutch, mean yolk volume and female size differed significantly between obligate burrowing and non-burrowing species.
Effect of intraclutch variation in yolk volume on hatching and growth of tadpoles
Larvae from large and small eggs taken from a single highly variable clutch of each of four species (L. nasuta, Cyclorana alboguttata, C. brevipes and C. novaehollandiae) were raised (Table 1). Each tadpole was raised in an individual container. To ensure that a wide range of yolk volume sizes was included in each experiment, equal numbers of the largest and smallest eggs from within a clutch were visually selected. Each egg was then assigned a number and individually digitally photographed to scale. The yolk volume of each egg was then determined as outlined above.
Each egg was placed in a circular plastic container (11 cm wide by 6 deep) containing 250 ml of aged tap water. Rearing experiments were conducted on one laboratory table divided into four areas, one for each species. Each experimental unit (one tadpole in one container) was assigned a random position within its area, with the constraint that two eggs from the same size class were never adjacent within a row or column of the array. Experimental units were positioned as tightly as possible within blocks to minimise the effects of different spatial locations.
Litoria nasuta eggs hatched after 3 days, this was designated day 0. The tadpoles were first weighed when they were large enough on day 8. L. nasuta tadpoles were then weighed and their water was changed daily. All Cyclorana eggs hatched 2 days after deposition, which was designated day 0. Cyclorana tadpoles were first weighed on day 6 and then were weighed and the water was changed every 3 days.
All tadpoles were fed a ground and sieved (250 μm) 3:1 mixture by weight of lucerne (alfalfa) pellets and TetraMin tropical fish food. Tadpoles were fed daily at a rate of 10 mg/day. Tadpoles were weighed by first placing them on a section of 60 micron plankton mesh, and blotting this on a cotton towel to remove excess water. The tadpole was then weighed to 0.1 mg precision by tipping it into a tared container of water on an electronic analytical balance.
Using initial egg size as a covariate, we carried out a repeated measures analysis of covariance (ANCOVA) for each species to examine the effects of yolk volume on the growth of tadpoles. This indicated whether there was an overall effect of initial yolk volume on the growth of tadpoles, whether initial yolk volume affected the shape of the growth curves, and determined whether the growth patterns of tadpoles from eggs with different initial yolk volumes differed. We then examined the forms of the growth curves for each species by plotting the time course of mass at age for each individual. This also enabled us to compare the relative growth patterns of individuals from eggs with large and small yolks. Mass was plotted on a logarithmic scale, which makes it easy to distinguish whether animals grow at constant, increasing, or decreasing proportional rates, and how the proportional growth rates of individuals change relative to one another over time. Exponential growth, growth at a constant proportional rate as occurs in many larvae (Alford and Jackson 1993), will lead to a linear relationship between size and age on such a plot. If individuals grow at different rates, but the proportional differences in their growth rates remain constant over time, their curves of size at age will remain separated by a constant distance.
After initially interpreting the growth patterns of each species graphically, we then examined in detail how the growth pattern of each species responded to initial egg volume by carrying out regressions of ln(mass at time t + 1) on ln(mass at time t) for each individual that survived to the end of the rearing experiment. The recursion equation for mass at time t + 1 obtained from each of these regressions is a power curve, of the form
where a is eintercept and k is the slope of the regression. Differences in the value of a, and thus the value of the intercepts of our regressions, to which a is directly related, can be interpreted as differences in the relative growth rates of individuals at size 1, at which size the value of k makes no difference to the growth rate. Higher values of a indicate that animals will grow more rapidly. The value of k indicates how the relative growth rate changes with mass. Values of k>1 mean that the relative growth rate increases with increasing mass, while values <1 indicate that it decreases. To determine whether individuals hatching from eggs of different yolk volumes followed different growth patterns, we performed regressions relating the values of the parameters a and k for each individual to that individual’s egg yolk volume.
Finally, to summarize the effects of egg yolk volume on growth patterns, we constructed response surface models for each species relating egg yolk volume to mass throughout the period of the experiment performed on that species. We used five initial egg yolk volumes, spanning the range of volumes found for the species, and estimated the parameters of the recursion relation for mass for individuals with each egg yolk volume from the regressions of these parameters on egg yolk volume for that species. We used mean masses on the first day of our experiments for individuals that hatched from eggs with volumes most similar to that defining each class as starting masses, and iterated the recursion equations to estimate mass at intervals over periods of time similar to those in our experiments.
Results
Relationships among female size, clutch size and yolk volume
We collected and examined a total of 158 clutches of 15 species in the summer of 1999/2000. We measured the yolk volumes of a total of 47,560 eggs. The number of eggs per clutch is significantly positively correlated with female mass only in Litoria caerulea, L. rothii and L. fallax (Table 2). Mean yolk volume increased with female mass only in L. caerulea (Table 2). There were no correlations between the number of eggs per clutch and mean yolk volume in any species even when controlling for female size (Table 2).
Comparison of obligate burrowing and non-burrowing species
There were no significant differences in female size or the number of eggs per clutch between obligate burrowers and non-burrowers (Table 3). Obligate burrowing species had a significantly larger mean egg size (Table 3, F 1, 13=5.348, P<0.05), and also had a significantly higher CV (Table 3, F 1, 13=21.437, P<0.001).
Patterns of intraclutch variation in yolk volume
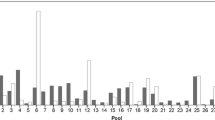
At least some clutches of all species had low CVs of yolk volume (Fig. 1). Some species also had some clutches with a high CV of yolk volume. These tend to be the larger species (Fig. 1). The baseline of the CV of yolk volume for most species is between 0.1 and 0.05 (Fig. 1). There was no relationship between the SD of CV and number of clutches examined across species. The species with the clutch in which the most variation in yolk volume occurred is C. alboguttata; the yolk of the egg at the 95th percentile was 3.3 times as large as that of the egg at the 5th percentile in this clutch. The species with the clutch in which the least variation in yolk volume occurred is Litoria fallax; the yolk of the egg at the 95th percentile was only 1.2 times as large as that of the egg at the 5th percentile within this clutch.

Intraclutch variation in yolk volume in 15 frog species from tropical savanna woodland in the Townsville area. Each point represents the CV of a clutch; species in each group arranged in order of mean female mass. Values below species = n
The genus Cyclorana had the highest degree of intraclutch variation in yolk volume, with a mean maximum CV of 0.32. The mean maximum CVs of yolk volume of the other genera were: 0.27 for Limnodynastes; 0.2 for Uperoleia; 0.17 for Litoria and 0.09 for Crinia. High intraclutch variation in yolk volume always occurred in Cyclorana (Hylidae), occurred in only some species of Litoria (Hylidae) and occurred in the myobatrachid genera Limnodynastes and Uperoleia but not Crinia, however, our sample sizes for the latter three genera are small.
The CV of yolk volume was significantly positively correlated with female mass only in L. fallax and L. rubella (Table 2), and was significantly negatively correlated with number of eggs per clutch in L. gracilenta (Table 2). Partial correlations controlling for the effect of female mass were not significant except for a negative relationship between the number of eggs per clutch and the CV of yolk volume in L. gracilenta (Table 2).
Effect of intraclutch variation in yolk volume on growth of tadpoles
The repeated measures ANCOVA found a significant effect of yolk volume on the ‘shapes’ (slope and curvilinearity) of the growth curves and the overall treatment means in all four species (Table 4).
Litoria nasuta tadpoles grew at increasing relative rates throughout the experiment (Fig. 2a). Tadpoles from eggs with a larger yolk volume were usually initially larger, and the proportional differences in size between individuals from eggs with large and small yolks remained approximately constant over time, as indicated by the spread of the lines for individuals (Fig. 2a). All of the slope coefficients of the individual regressions of mass at time t + 1 on mass at time t are greater than 1, indicating that growth rate increased with increasing size (Fig. 2b), and the values of the slope coefficients are positively correlated with egg yolk volume, indicating that growth rates of individuals from eggs with larger yolks increased more rapidly with mass than did those of individuals from smaller-yolked eggs. The differences in size between individuals from eggs with large and small yolks remained proportionally similar throughout the experiment because the effect of yolk volume on slope was approximately offset by its effect on the intercept, which decreased with increasing yolk volume (Fig. 2c). Although the relative size differences between individuals from large- and small-yolked eggs remained nearly constant throughout the experiment, growth rates calculated for individuals late in the experiment (Table 5) were greater for individuals from small-yolked eggs, which would have caused a rapid divergence if animals had remained in the aquatic habitat any longer. However, the experiment was terminated after day 21 because the larger tadpoles had begun to metamorphose. The response surface (Fig. 2d) for L. nasuta illustrates the very large differences in absolute size between individuals from large and small yolked eggs that existed late in the experiment, and the fact that yolk volume had a strong effect upon time to metamorphosis.

a, e, i, m Growth of tadpoles from eggs with large yolk volumes (dashed lines) and eggs with small yolk volumes (solid lines); b, f, j, n Relationship between yolk volume and the slope (growth rate) from the regressions of ln(mass at time t + 1) on ln(mass at time t) for each individual; c, g, k, o Relationship between yolk volume and the intercept (size at first measurement) from the regressions of ln(mass at time t + 1) on ln(mass at time t) for each individual; d, h, l, p response surface models for each species relating egg yolk volume to mass throughout the experimental period
The growth patterns of the three Cyclorana species were similar to one another (Fig. 2), and differed greatly from that of L. nasuta. In all three species, there was a tendency for individuals from eggs with larger yolk volumes to be larger at each age than individuals from smaller-yolked eggs (Fig. 2e, i, m). All three species followed approximately exponential growth patterns [linear increase of log(mass) with time] for the first 7–9 days of measurement (Fig. 2e, i, m). During this exponential phase, the proportional differences in size among individuals remained nearly constant, as indicated by the spread among the growth curves. At about day 13–14 of the experiment, the growth rate of each species began to decrease markedly with time, and during this phase, proportional differences between individuals from eggs with large and small yolks decreased. The coefficients of the individual regressions of mass at time t + 1 on mass at time t were significantly affected by egg yolk volume (Table 5). However, their relationships to egg yolk volume were the reverse of those found in L. nasuta. The slopes of the regressions are all less than 1, indicating that relative growth rate decreased with increasing size. The slopes also decreased with increasing egg yolk volume, indicating that the decrease in relative growth rate with increasing size occurred more rapidly in individuals from eggs with larger yolks. The decreasing slopes were countered, particularly in individuals near the beginnings of their growth curves, by relatively large intercepts, which were positively related to yolk volume (Table 5, Fig. 2g, k, o). These meant that small individuals grew relatively rapidly, and individuals from eggs with larger yolks grew more rapidly than individuals from smaller yolks. The overall patterns produced by these relationships are illustrated in the response surfaces (Fig. 2h, l, p). Individuals from eggs with the largest yolks initially grew rapidly, and reached sizes of 100 mg in about half the time taken by individuals from eggs with the smallest yolks. Because relative growth rates of all individuals decreased with increasing mass, individuals from larger-yolked eggs began to grow at slower rates earlier than those from smaller-yolked eggs. Although in two of the three species (C. novaehollandiae and C. alboguttata, Fig. 2h, l) large differences remained between individuals from large- and small-yolked eggs late in the experiment, in all three species the sizes of individuals from eggs with different yolk volumes would eventually have converged. All individuals were in early stages of hindlimb development at the termination of the experiment on day 24.
Discussion
We examined the clutch parameters of an assemblage of tropical frogs in detail. We precisely counted the numbers of eggs per clutch, and precisely measured yolk volumes of large samples of eggs using digital imaging and processing methods. We sampled clutches of 15 of the 23 species that occur in tropical savanna woodland around Townsville. This provided a more complete picture of the clutch size parameters and degree of variation within and among clutches and species for an assemblage than has been amassed by any other study to date (cf. Crump 1981b, 1984; Kaplan 1992; Tejedo and Reques 1992; Williamson and Bull 1989).
Relationships among female size, clutch size and yolk volume
Significant correlations between frog size and the number of eggs per clutch and mean yolk volume occurred only in the genus Litoria. The number of eggs per clutch depended significantly on female size in three species (Table 2). In the remaining species body size and fecundity were not significantly correlated. This differs from the findings of other studies, (e.g. Williamson and Bull 1995), which have found that clutch size increases with body size within a species, a relationship so common that it has been proposed as being typical for anurans (Crump 1974; Kaplan and Salthe 1979; Kuramoto 1978). There were no relationships between the number of eggs per clutch and mean egg size even when controlling for female size. This indicates that there is no trade-off between yolk volume and egg number, as a negative correlation would exist if a trade-off occurred. Trade-offs between components of reproductive output are expected if there is a body size constraint on reproductive output (Kaplan and Salthe 1979; Vitt and Congdon 1978). Our results thus suggest that all 15 species in this study do not expend the maximum amount of reproductive effort allowed by their body size. This also occurs in B. calamita (Tejedo 1992), but not in C. signifera (Williamson and Bull 1995) or Rana temporaria (Cummins 1986); in both of these species there is a trade-off between egg size and number. This lack of trade-off between egg size and number, however should not influence the evolution of variability in egg size as this component should evolve separately in response to the predictability of the offspring environment.
Patterns of intraclutch variation
We found relatively high levels of intraclutch variation in many of the species that we examined. In all species, apart from L. gracilenta, the extent of intraclutch variation in yolk volume was not correlated with mean yolk volume or clutch size, even when the effects of female body size were accounted for. This suggests that intraclutch variability is evolutionarily independent from yolk volume and clutch size. This agrees with theory which suggests that mean yolk volume and clutch size evolve in response to trade-offs among fecundity, growth, and survival of offspring for any given simple constant environment (Lloyd 1987; Smith and Fretwell 1974), while the occurrence and degree of intraclutch variation are determined by the variability of the physical and biological environments that a species encounters (Capinera 1979; Kaplan and Cooper 1984). It is therefore not surprising that intraclutch variability is an independent phenomenon occurring over a range of species of small and large sizes. It has previously been documented in small species (C. signifera, ∼1 g; Williamson and Bull 1995), medium sized species (B. orientalis, ∼7 g; Kaplan 1992) and larger species (B. calamita, ∼70 g; Tejedo and Reques 1992). Our study is the first to show that both high and low levels of intraclutch variation occur within species for a variety of species occupying the same area. The fact that there was no relationship across species between the SD of CV and sample size, indicates that differences among species in the number of clutches examined had no effect on CV; this reinforces our conclusion that there are species-specific differences in how much variation there is within clutches. Although our within-clutch sample size varied from 80 to 800, estimates of variance are usually thought to stabilize for sample sizes of 30 or higher (Zar 1999), so our estimates of CV should not have been affected by the differences in within-clutch sample size. Our samples of the genera Crinia and Uperoleia included few clutches because the small size of adults of these species, and the cryptic locations in which they enter amplexus (under leaf litter and vegetation) made females of these species difficult to locate. Since species with higher sample sizes that had higher levels of variation also had some clutches with low variation, it is quite possible that the genera Crinia and Uperoleia may produce some clutches with higher levels of variation, not detected in our survey.
The lack of correlation of degree of intraclutch variation with other clutch and egg parameters may reflect the high intraspecific variation we found in levels of intraclutch variation (Fig. 1). Nine of the 15 species with relatively large sample sizes (≥8 clutches) produced some clutches with relatively high levels of intraclutch variation in yolk volume, but each of these species also produced some clutches with low levels of variation. Our analyses showed that intraspecific differences among clutches in the degree of yolk size variability were not a consequence of simple relationships between variability and body size or other reproductive parameters. Variation in yolk volume was significantly positively correlated with female size only in L. rubella and L. fallax, the two smallest species of that genus we examined. In L. gracilenta, intraclutch variability decreased with the number of eggs per clutch, and there was no trade-off between mean egg size and number. This suggests that in this species, as total reproductive investment decreases, the remaining energy is disproportionately invested in larger eggs and withdrawn from smaller eggs.
Comparison of obligate burrowing and non-burrowing species
Obligate burrowing species had larger mean egg sizes. This could be predicted since these species can occur in drier, more arid areas (Barker et al. 1995; Cogger 2000) where temporary ponds are of shorter duration. In these environments, a larger egg size, which results in increased growth rate and reduced development time (Berven and Chadra 1988; Kaplan 1992; Parichy and Kaplan 1992; this study) is required for successful reproduction.
Species that are obligate burrowers had higher levels of intraclutch variation in yolk volume than non-burrowing species. This pattern supports the prediction that intraclutch variation in offspring size should be favoured in anurans that use unpredictable temporary ponds for reproduction (Crump 1981b, 1984), a relationship that has also been found in fish (Koops et al. 2003). The variability within species in the degree of intraclutch variation may reflect an ability of females to adjust their level of intraclutch variation in response to environmental or biotic conditions, producing clutches that best fit the level of unpredictability of the local environment (Capinera 1979). Under this model, females may respond to cues suggesting that the environment occupied by their present clutch will be predictable by producing an optimal egg size for that environment; if such cues are absent or suggest an unpredictable offspring environment, they will produce clutches containing more variable eggs. More data for single species from a number of localities and specific data on offspring conditions would be needed to test this prediction.
Our data do allow us to eliminate one possible explanation for variation within species in levels of intraclutch variation in egg volume. If eggs are provisioned sequentially, females could be at different stages of egg provisioning when the optimal conditions for spawning occur. In this case, females who had completed egg provisioning would produce clutches of relatively large eggs with low variation in yolk volume, and females ‘caught out’ at earlier stages of egg provisioning might produce more variable clutches with some relatively large and some relatively small eggs. The lack of significant within-species correlations of level of within-clutch variability with mean egg volume indicate that the more variable clutches include eggs that are both larger and smaller than the less variable clutches, which is inconsistent with this explanation. It remains possible that some individual frogs of some species simply do not regulate egg size well. If this is the case, the fitness consequences we have demonstrated should lead to strong selection for or against these individuals, depending on environmental conditions. The present day distributions of intraclutch variability within populations could reflect the long-term results of fluctuating selection. In our species assemblage, high intraclutch variation in yolk volume occurred across most genera and in both families. If it is a derived character it has probably evolved more than once, especially in the burrowing species of the myobatrachid and hylid families. It is also possible that it is a primitive character. Resolving this will require more data across a wider range of amphibian taxa.
Effect of intraclutch variation in yolk volume on growth of tadpoles
For intraclutch variation in egg provisioning to evolve, the differences in size and growth among larvae from eggs with different yolk volumes must lead to differences in fitness. These fitness effects are likely to arise because egg provisioning level affects the ecology of the resulting larvae. Relationships of larval ecology to growth and development are well documented, as are size-specific ecological interactions (e.g. Wilbur and Collins 1973; review in Alford 1999). A number of studies examining general ecological interactions in anuran larval communities have found that larger sizes and higher growth rates provide benefits under density dependant competitive interactions (e.g. Morin and Johnson 1988; Savage 1952; Steinwascher 1978; Travis and Trexler 1986) and predation (Persson 1988; Richards and Bull 1990; Wilbur 1988). Higher growth rates may allow individuals to escape (by earlier metamorphosis) from the effects of density or from other environmental effects, such as pond evaporation (Crump 1989; Tejedo and Reques 1995). The amount of energy invested in each egg is known to affect the resultant offspring size or growth (Lloyd 1987; Smith and Fretwell 1974; Winkler and Wallin 1987). Our growth experiment identified differences in tadpole size and growth patterns that resulted directly from differences in initial yolk volume. This is consistent with other studies that have also found effects of initial egg provisioning on offspring growth and size (Andrén et al. 1989; Crump 1984; Kaplan 1992; Tejedo and Reques 1992; Williamson and Bull 1989).
In L. nasuta, growth was slightly faster than exponential throughout the experiment, with the size-specific growth rate slowly increasing to the end of the experiment at day 20, shortly before metamorphosis. In the three Cyclorana species, early growth appeared to be exponential, occurring at near-constant size-specific rates. Growth rates of Cyclorana began to decrease between day 12 and 15. Within a clutch, in all four species, eggs with greater investment produce initially larger offspring; a result that is consistent with previous studies (Crump 1984; Kaplan 1992; Tejedo and Reques 1992; Williamson and Bull 1989). We also found that the initial size differences persist over a substantial fraction of larval life, and may lead to earlier metamorphosis.
In all four species, tadpoles from eggs with smaller yolk volumes were initially smaller than those from larger-yolked eggs but had higher proportional growth rates. In L. nasuta, the growth rates of tadpoles from larger-yolked eggs increased more rapidly with increasing size than the growth rates of tadpoles from eggs with smaller yolks, so that by the end of the experiment, as the tadpoles from larger yolks neared metamorphosis, they were growing proportionally more rapidly than tadpoles from smaller yolks. This indicates that, under benign conditions, tadpoles of L. nasuta from larger-yolked eggs metamorphose substantially earlier than those from eggs with smaller yolks. Even if the lagging tadpoles eventually metamorphose at the same body size, they will have a greater stage-specific exposure to risks such as predation and disease, and therefore are likely to have lower survival to first reproduction, and lower fitness.
Our growth experiment showed that tadpoles from eggs with larger yolks hatched at larger sizes than tadpoles from eggs with smaller yolks. This size difference persisted throughout a substantial fraction of the larval stage. Even if the body sizes of individuals from large and small yolked eggs converged before metamorphosis, it is likely that the individuals from the larger yolks would metamorphose earlier, and it is certain that they would spend less time at smaller sizes. Many predators of tadpoles are size-limited (Alford 1999), and individuals from large-yolked eggs would experience lower cumulative risk from such predators. It therefore is likely that the variation we observed in egg provisioning usually leads to variation in the fitness of individuals under field conditions. In the three Cyclorana species, tadpoles from larger-yolked eggs initially had higher growth rates than those from smaller yolks, but size-specific growth rate decreased more rapidly in individuals from larger yolks. By the end of the experiment, it appeared (Fig. 2) that tadpoles from smaller-yolked eggs were catching up in body size to those from larger yolks. Because this experiment ended before any individuals progressed beyond early stages of hindlimb development, we cannot be certain that the individuals from larger-yolked eggs would metamorphose any earlier or at larger sizes than those from smaller-yolked eggs, when raised under our experimental conditions. We suspect that one of the causes of the convergence in size between individuals from large and small yolks later in our experiment may be that our rearing conditions were suboptimal. All three of the Cyclorana species can metamorphose with body weights of two or more g (R.A.A., personal observation), and may require larger volumes of water than were available in our experimental containers. The fact that their growth curves appeared to be approaching asymptotic body sizes of less than 1 g suggests that growth conditions were not good, and that the faster-growing individuals approached the limits set by their rearing environment earlier than the slower-growing ones. Even if that is the case, the tadpoles from larger yolks might have metamorphosed first, as occurs in other species (Kaplan 1992). All tadpoles in our experiment were fed the same amount. Tadpoles from eggs with less yolk volume were smaller and therefore had relatively more food available. The energy from this extra food may have been used to produce an elevated proportional growth rate in tadpoles from smaller eggs. This could be controlled for by providing food ad libitum, but this could also lead to fouling effects (Alford 1999).
These fitness differentials between offspring from different sized eggs have important parental fitness consequences in species that use offspring environments that vary unpredictably in quality. If the offspring environment happens to be of good quality, for example a persistent pond or a pond with low predation or competition pressures, then all offspring may survive. If however the offspring environment happens to be of poor quality, for example a quickly drying pond or a pond with high predation or competition pressures, then only the offspring from large eggs may survive. This strategy ensures that at least some offspring will survive. This form of bet-hedging should maximise the lifetime reproductive success of individuals that engage in it, and be selected for under these conditions.
Conclusions
We found that many species, with a range of phylogenetic affinities and body sizes, in an assemblage of savanna woodland frogs, produce some clutches having high variability in egg provisioning. We found no trade-off between egg size and number in any species, however variation in egg provisioning should evolve independently in response to the offspring environment. We demonstrated that species that occur in habitats where the offspring environment is likely to be unpredictable have elevated levels of intraclutch variation in egg provisioning. We also showed that this variation is likely to lead to fitness effects on offspring, resulting in effects on parental fitness, therefore becoming a selection variable. Our results are consistent with the prediction by McGinley et al. (1987) that under certain circumstances, bet-hedging by varying investment in offspring within clutches should evolve in response to environmental unpredictability. We suspect that variation similar to that which we found may be common in frogs in many habitats; it may have been previously undiscovered simply because making the necessary measurements has been difficult.
References
Alford RA (1999) Ecology: resource use, competition and predation. In: McDiarmid RW, Altig R (eds) Tadpoles: the biology of anuran larvae. University of Chicago Press, Chicago
Alford RA, Jackson GD (1993) Do cephalopods and the larvae of many taxa grow asymptotically? Am Nat 141:717–728
Andrén C, Marden MM, Nilson G (1989) Tolerance to low pH in a population of moor frogs, Rana arvalis, from an acid and a neutral environment: a possible case of rapid evolutionary response to acidification. Oikos 56:215–223
Barker J, Grigg GC, Tyler MJ (1995) A field guide to Australian frogs. Surrey Beatty & Sons, Sydney
Berven KA, Chadra BG (1988) The relationship among egg size, density and food level on larval development in the wood frog (Rana sylvatica). Oecologia 75:67–72
Capinera JL (1979) Quantitative variation in plants and insects: effect of propagule size on ecological plasticity. Am Nat 114:350–361
Cogger HG (2000) Reptiles and amphibians of Australia, 6th edn. Reed New Holland, Sydney
Crump ML (1974) Reproductive strategies in a tropical anuran community. Miscellaneous Publ Mus Nat Hist Univ Kansas 61:1–68
Crump ML (1981a) Energy accumulation and amphibian metamorphosis. Oecologia 49:167–169
Crump ML (1981b) Variation in propagule size as a function of environmental uncertainty for tree frogs. Am Nat 117:724–737
Crump ML (1984) Intraclutch egg size variability in Hyla crucifer (Anura: Hylidae). Copeia 1984:302–308
Crump ML (1989) Effect of habitat drying on developmental time and size at metamorphosis in Hyla pseudopuma. Copeia 1989:794–797
Cummins CP (1986) Effects of aluminium and low pH on growth and development in Rana temporaria tadpoles. Oecologia 69:279–283
Einum S, Fleming IA (2004) Environmental unpredictability and offspring size: conservative versus diversified bet-hedging. Evol Ecol Res 6:443–455
Harris RN (1999) The anuran tadpole: evolution and maintenance. In: McDiarmid RW, Altig R (eds) Tadpoles: the biology of anuran larvae. University of Chicago Press, Chicago
Kaplan RH (1979) Ontogenetic variation in ovum “size” in two species of Ambystoma. Copeia 1979:348–350
Kaplan RH (1992) Greater maternal investment can decrease offspring survival in the frog Bombina orientalis. Ecology 73:280–288
Kaplan RH, Cooper WS (1984) The evolution of developmental plasticity in reproductive characteristics: an application of the “adaptive coin-flipping” principle. Am Nat 123:393–410
Kaplan RH, King EG (1997) Egg size is a developmentally plastic trait: evidence from long term studies in the frog Bombina orientalis. Herpetologica 53:149–165
Kaplan RH, Salthe SN (1979) The allometry of reproduction: an empirical view in salamanders. Am Nat 113(5):671–689
Koops MA, Hutchings JA, Adams BK (2003) Environmental predictability and the cost of imperfect information: influences on offspring size variability. Evol Ecol Res 5:29–42
Kuramoto M (1978) Correlations of quantitive parameters of fecundity in amphibians. Evolution 32:287–296
Lloyd DG (1987) Selection of offspring size at independence and other size-versus-number strategies. Am Nat 129:800–817
McGinley MA, Temme DH, Geber MA (1987) Parental investment in offspring in variable environments: theoretical and empirical considerations. Am Nat 130:370–398
Morin PJ, Johnson ES (1988) Experimental studies of asymetric competition among anurans. Oikos 53:398–407
Parichy DM, Kaplan RH (1992) Maternal effects on offspring growth and development depend on environmental quality in the frog Bombina orientalis. Oecologia 91:579–586
Persson JW (1988) Asymmetries in competitive and predatory interactions in fish populations. In: Ebenmann B, Persson L (eds) Size-structured populations. Springer, Berlin Heidelberg New York, pp 203–218
Richards SJ, Bull CM (1990) Size-limited predation on tadpoles of three Australian frogs. Copeia 1990:1041–1046
Rugh R (1951) The frog. Its reproduction and development. McGraw-Hill, New York
Savage RM (1952) Ecological, physiological and anatomical observations on some species of anuran tadpoles. Proc Zool Soc Lond 122:467–514
Slade NA, Wassersug RJ (1975) On the evolution of complex life cycles. Evolution 29:568–571
Smith CC, Fretwell SD (1974) The optimal balance between size and number of offspring. Am Nat 108:499–506
Steinwascher K (1978) Interference and exploitation competition among tadpoles of Rana utricularia. Ecology 59:1039–1046
Tejedo M (1992) Absence of the trade-off between the size and number of offspring in the natterjack toad (Bufo calamita). Oecologia 90:294–296
Tejedo M, Reques R (1992) Effects of egg size and density on metamorphic traits in tadpoles of the natterjack toad (Bufo calamita). J Herpetol 26:146–152
Tejedo M, Reques R (1995) Plasticity in metamorphic traits of natterjack tadpoles: the interactive effects of density and pond duration. Oikos 71:295–304
Travis J, Trexler JC (1986) Interactions among factors affecting growth, development, and survival in experimental populations of Bufo terrestris (Anura: Bufonidae). Oecologia 69:110–116
Vitt LJ, Congdon JD (1978) Body shape, reproductive effort, and relative clutch mass in lizards: resolution of a paradox. Am Nat 112:595–608
Wassersug RJ (1975) The adaptive significance of the tadpole stage with comments on the maintenance of complex life cycles in anurans. Am Zool 15:405–417
Wilbur HM (1988) Interactions between growing predators and growing prey. In: Ebenmann B, Persson L (eds) Size structured populations. Springer, Berlin Heidelberg New York, pp 157–172
Wilbur HM, Collins JP (1973) Ecological aspects of amphibian metamorphosis. Science 182:1305–1314
Wilcox CD, Dove SB, McDavid WD, Greer DB (1995) UTHSCSA ImageTool. Version 2a. University of Texas Health Science Center, San Antonio
Williamson I, Bull CM (1989) Life history variation in a population of the Australian frog Ranidella signifera: egg size and early development. Copeia 1989:349–356
Williamson I, Bull CM (1995) Life-history variation in a population of the Australian frog Ranidella signifera: seasonal changes in clutch parameters. Copeia 1995:105–113
Winkler DW, Wallin K (1987) Offspring size and number: a life history model linking effort per offspring and total effort. Am Nat 129:708–720
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice Hall, New Jersey
Acknowledgements
Sacha Jellinek provided excellent assistance with fieldwork and experiments and the amount of data collected would not have been possible without his help. R. Black, J. D. Roberts and P. Doughty provided very helpful comments on the manuscript. All research in this study received ethical clearance from the James Cook University Experimentation Ethics Review Committee (Approval Number A452) and all collection of animals was conducted with and within the guidelines of a Scientific Purposes Permit (Permit Number N0/001257/96/SAA) issued by the Department of Environment and Heritage, Queensland, Australia. The research was funded by a minor research grant to RAA from James Cook University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Roland Brandl
Rights and permissions
About this article
Cite this article
Dziminski, M.A., Alford, R.A. Patterns and fitness consequences of intraclutch variation in egg provisioning in tropical Australian frogs. Oecologia 146, 98–109 (2005). https://doi.org/10.1007/s00442-005-0177-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-005-0177-2