
Abstract
Corridors connect otherwise isolated habitat patches and can direct movement of animals among such patches. In eight experimental landscapes, we tested two hypotheses of how corridors might affect dispersal behavior. The Traditional Corridor hypothesis posits that animals preferentially leave patches via corridors, following them into adjacent patches. The Drift Fence hypothesis posits that animals dispersing through matrix habitat are diverted into patches with corridors because they follow corridors when encountered. House flies (Musca domestica L.), a species that prefers the habitat of our patches and corridors, were released in a central patch (100×100 m) and recaptured in peripheral patches that were or were not connected by a corridor. Flies were captured more frequently in connected than unconnected patches, thereby supporting the Traditional Corridor hypothesis. The Drift Fence hypothesis was also supported, as flies were captured more frequently in unconnected patches with blind (dead end) corridors than in unconnected patches of equal area without blind corridors. A second experiment tested whether these results might be dependent on the type of patch-matrix boundary encountered by dispersing flies and whether edge-following behavior might be the mechanism underlying the observed corridor effect in the first experiment. We recorded dispersal patterns of flies released along forest edges with dense undergrowth in the forest (“closed” edges) and along edges with little forest understory (“open” edges). Flies were less likely to cross and more likely to follow closed edges than open edges, indicating that when patch and corridor edges are pronounced, edge-following behavior of flies may direct them along corridors into connected patches. Because edges in the first experiment were open, these results also suggest that corridor effects for flies in that experiment would have been even stronger if the edges around the source patches and corridors had been more closed. Taken together, our results suggest that corridors can affect dispersal of organisms in unappreciated ways (i.e., as drift fences) and that edge type can alter dispersal behavior.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Habitat loss and fragmentation isolate populations of organisms and reduce their size, thereby increasing the risk of local extinction (Brown and Kodricbrown 1977; Gonzalez et al. 1998; Harrison and Bruna 1999). This threat is especially severe for species reluctant to enter matrix habitat (Berggren et al. 2002). One means of overcoming this reluctance is the creation of corridors, defined as strips of habitat connected to patches of the same habitat type (Rosenberg et al. 1997; Beier and Noss 1998). Corridors are hypothesized to facilitate movement of organisms between fragments, increasing gene flow and maintaining or reestablishing populations in small patches of habitat (Dunning et al. 1995; Aars and Ims 1999; Mech and Hallett 2001).
The utility of habitat corridors remains controversial, however. Some studies conclude that corridors indeed increase movement of animals between otherwise isolated habitat fragments (Beier and Noss 1998; Debinski and Holt 2000; Tewksbury et al. 2002; Haddad et al. 2003). Other studies find little evidence of such an effect (Arnold et al. 1991; Date et al. 1991; Rosenberg et al. 1998; Bowne et al. 1999; Collinge 2000; Danielson and Hubbard 2000). In part, the controversy has been generated by a mismatch in scale between experimental studies and conservation plans. Most experiments on the efficacy of corridors have been conducted on small scales, and experiments conducted on larger scales have often been poorly replicated (Holt and Debinski 2003).
Another reason that corridor studies have aroused controversy is that most have failed to distinguish between effects due to increased patch colonization caused by dispersal along corridors versus those caused by increased area associated with corridors —perhaps some corridor effects are actually area effects. Finally, corridors may function in unexplored ways (Simberloff et al. 1992). For example, instead of channeling the movement of organisms between connected patches of habitat, corridors may function as “drift fences,” intercepting individuals as they disperse through matrix habitat and redirecting them into associated patches (Taylor 1987; Anderson and Danielson 1997; Haddad and Baum 1999).
Two different mechanisms by which corridors may facilitate patch colonization are expressed by non-mutually exclusive hypotheses. The Traditional Corridor hypothesis posits that corridors function as movement conduits. Animals preferentially leave source patches via corridors, following them to adjacent, connected patches. The Drift Fence hypothesis posits that corridors function as drift fences—animals dispersing through matrix habitat colonize a patch with a corridor by encountering the corridor and following it (Haddad and Baum 1999). Corridors do not have to connect patches to function in this way.
We tested the Traditional and Drift Fence hypotheses with house flies (Musca domestica L.) in large scale experimental landscapes. Habitat patches were areas of second growth, some connected by corridors and others unconnected. The design controlled for increased area associated with the presence of a corridor.
Corridors are most likely to function as hypothesized in situations where habitat-restricted animals perceive a stark difference between favorable habitat in the patch they occupy and unfavorable matrix habitat (Ricketts 2001). Because results from the corridor experiment described above revealed that flies often entered the matrix habitat, the difference between patch and matrix habitat was likely not as distinct for flies as we anticipated. Therefore, we conducted a second experiment to evaluate how different types of habitat boundaries influence fly dispersal. In particular, we compared dispersal along field–forest boundaries in which the forest had little understory (“open” edge) and those in which the forest had a dense understory (“closed” edge). To facilitate comparison, we selected open edges that were nearly identical in openness to those encountered by flies in the corridor experiment. A second purpose of the edge experiment was to examine whether edge-following behavior might have been the behavioral mechanism underlying the flies’ use of corridors.
2 Materials and methods
2.1 Corridor experiment
2.1.1 Study site
The corridor experiment was conducted at the Savannah River National Environmental Research Park near Aiken, South Carolina (USA). Forty patches of second growth were created in mature stands of loblolly pine (Pinus taeda) and longleaf pine (P. palustris). Trees were cut and removed in the winter of 1999–2000 and debris was burned in the spring of 2000. At the time of our experiment, patches had a dense cover of grasses, herbs, and shrubs (<1.5 m height), with frequent small trees that were regenerating from stump sprouts. From a fly’s perspective, the patches had a rich diversity of microhabitats, including many areas of shade. Due to prescribed burns every 3–5 years previously, the forest understory in the matrix was sparse.
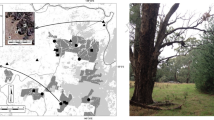
We created eight experimental units, each consisting of five patches, with one 100×100 m central patch (“source” patch) surrounded by four “receiver” patches, one on each side and 150 m away (Fig. 1). One receiver patch (“connected receiver”) was connected to the source patch by a 25-m-wide strip of identical habitat. The three remaining receiver patches were of two types, both unconnected and equal in area to the connected receiver patch plus its corridor. “Winged receiver” patches were 100×100 m with two blind-ended, 75 m corridors projecting from opposite sides of the patch, perpendicular to the travel direction of flies dispersing from the source patch. “Rectangular receiver” patches were 100×137.5 m. The “extra” 37.5×100 m of these patches represents the total area of the corridor and was added to the side farthest from the source patch, such that the patch’s cross-sectional dimension relative to flies dispersing from the source patch remained 100 m. Four of the eight experimental units had two winged and one rectangular receiver patch, and the other four had one winged and two rectangular receiver patches. All had a single connected receiver patch.

Enlargement of one experimental landscape, showing patch types (source, connected receiver, winged receiver, rectangular receiver), point where flies were released in source patch, and location of traps in receiver patches. Patch habitat was young second growth; shading represents mature forest. A total of eight experimental landscapes were located at the Savannah River Site (SRS) in South Carolina, USA and were separated by at least 5 km. Four experimental landscapes had two winged and one rectangular receiver patches, and four had one winged and two rectangular receiver patches. Locations of receiver patches around each source patch were randomized for each landscape
Patch type was assigned randomly within experimental units, as was compass direction of the corridor. Care was taken to minimize disturbance to the understory of the forest matrix when removing trees from the patches. A buffer area extending >150 m from the outside edge of all receiver patches was free of paved roads and other clearings and was the same type of forest that occurred between patches.
2.1.2 Study species
We obtained flies for release and recapture from colonies at the USDA-ARS, Center for Medical, Agricultural, and Veterinary Entomology (CMAVE) on the University of Florida campus in Gainesville, Florida. Larvae were grown on a grain diet, pupae were separated by flotation, and adults were allowed to emerge in screen cages, where they were provided with water and food (sugar, powdered milk and dried egg yolk) until release (Hogsette 1992).
To determine the extent to which house flies might distinguish between our patch and matrix habitats, we conducted a small study by placing fly traps (see below) in ten open habitat sites and ten forest understory sites. After 4 days of trapping, significantly more flies were captured in the open than understory sites (Mann–Whitney U=7.5; P=0.03).
2.1.3 Release and recapture
At the center of each source patch, we released approximately 3,750 flies (estimated by mass of pupae) that were 2- to 3-days-old. Flies were released between 07:00 and 09:00 in June and July 2001. A single release was made in each experimental unit, all on days with no precipitation. To eliminate potential directional biases, we released the flies through an opening in the top of the cage and left the area as quickly as possible.
We used jug-style Captivator Fly Traps (Farnam Companies, Inc., Phoenix, AZ, USA), with entrance holes at the top. A floor of window screen prevented flies from falling into an aqueous bait solution in the bottom of the trap. We prepared the bait solution by diluting 120 mL of “ Musca attractant” (Farnam Companies, Inc., Phoenix, AZ, USA) in 3.8 L of water and added 500 mL of this solution to each trap.
We placed four traps in each receiver patch (n=16 traps per experimental unit), positioned on or near the edge of the patch closest to the source patch and suspended approximately 1 m above the ground. Two traps were placed on the patch’s edge, 12.5 m in each direction from the midpoint of that edge. The other two traps were placed within the patch, 18 m at a 45° angle from the patch corners nearest the other two traps (Fig. 1). These locations were determined by the position of poles already established in the patches (Tewksbury et al. 2002), from which we could hang the traps.
We prepared bait and hung baited traps the afternoon before each release of flies. Before releasing flies the next morning, we checked all traps for naturally occurring flies, which were excluded in subsequent analyses. These checks revealed that naturally occurring flies were present but rare (averaging approximately 0.1 fly per trap), so we assumed that all flies captured after a release at a given experimental unit were from that release. Flies were collected the afternoon of the day following a release, approximately 32 h after release. A pilot study showed that cumulative captures reached an asymptote by this time.
2.2 Edge experiment
2.2.1 Study site
The edge experiment was conducted at the Katharine Ordway Preserve-Swisher Memorial Sanctuary in western Putnam County, Florida. The preserve is comprised of fields and upland forests that primarily consist of longleaf pine (P. palustris) and turkey oak (Quercus laevis). In many places, the interface between the forest and adjacent fields is sharply delineated, creating edges similar in structure to those that flies encountered in the corridor experiment. Depending on burn frequency, soil type, and other factors, some edges are “closed” (dense understory on the forest side) and some are “open” (little understory on the forest side). We chose ten edges that were clearly defined (i.e., a distinct boundary between field and forest, as in the corridor experiment) and fairly linear for approximately 250 m. Five edges were open, and five were closed.
To quantify the difference between these edge types and to confirm that the open edges were similar to those encountered by flies in the corridor experiment, one person held a white cloth (0.5 by 0.5 m) 1 m above the ground, 25 m into the forest from the edge. Standing in the field, 2 m away from the edge, a second person scored by quartile the proportion of cloth that was not obscured by foliage (1=0–25% visible, 2=26–50% visible, 3=51–75% visible, 4=76–100% visible). At each edge, we scored openness in this way at nine points, separated by 25 m along a transect parallel to the edge. For comparison, we collected identical data along two randomly selected edges for each experimental unit in the corridor study’s patches.
2.2.2 Release and recapture
Along each edge, we positioned a total of 14 baited Captivator traps in three parallel 200-m transects. One transect was along the edge, one was 50 m into the woods, and one was 50 m into the field. The forest and field transects each had five traps, spaced 50-m apart. The edge transect had 4 traps, positioned exactly halfway between adjacent traps in the field and forest transects (i.e., at 50 m, 100 m, 150 m, and 200 m). There was no trap at 0 m for the edge transect because that was where we released the flies. To control for effects of weather on fly behavior, we used a paired design, releasing flies at one open and one closed edge on a given day. In all other respects, techniques for raising, releasing, and recapturing flies were identical to those described for the corridor experiment.
2.3 Analyses
We treated the corridor experiment as an unbalanced randomized block design (unbalanced due to the different number of winged and rectangular patches in each experimental unit) and used a Mixed Linear Model to analyze number of flies captured per trap (Proc MIXED, SAS v. 8.1, SAS Institute). Experimental unit was treated as a random effect and patch-type (winged, rectangular, or connected) as a fixed effect. Transformation of data was unnecessary. A post-hoc contrast (with Bonferroni correction) between connected and rectangular patches tested the Traditional Corridor hypothesis, which predicts more captures in connected than rectangular patches. A post hoc contrast between winged and rectangular patches tested the Drift Fence hypothesis, which predicts more captures in winged than rectangular patches.
For the edge experiment, we used a General Linear Model (Proc GLM, SAS v. 8.1, SAS Institute) that included the following independent factors: type of edge (open, closed), habitat in which a trap was placed (forest, edge, field), position along the transect, date of release, and the interaction between habitat and position. All other interaction terms were removed from the full model because they were non-significant (P values >0.27). Number of flies captured per trap was the dependent variable.
The General Linear Model tested whether type-of-edge influenced the general pattern of dispersal by flies released at an edge. We conducted a second analysis to determine the extent to which the type of edge influenced the probability that flies would be captured along the edge (i.e., the extent to which they followed edges during dispersal). Using a contingency table (Proc FREQ, SAS v. 8.1, SAS Institute), we compared the proportion of flies caught along the edge versus the proportion caught in field and forest, combined, for each type of edge.
3 Results
3.1 Corridor experiment
A total of 1,226 flies were captured, averaging 153±19 (SE) per experimental unit. Type of patch strongly influenced capture rate (F 3,118=24.32; P=0.006; Fig. 2). Captures also varied among the eight experimental landscapes (Z=7.68, P=0.045). Variation among sites was mainly due to differences in total captures, not direction of differences in captures among patch types. Connected patches had significantly higher captures than rectangular patches (F 1,118=8.37, P=0.009), which supports the Traditional Corridor hypothesis. Likewise, winged patches had significantly higher captures than rectangular patches (F 1,118=8.18, P=0.010), which supports the Drift Fence hypothesis. The difference in captures between patch types, 21%, was practically identical for both of these post hoc comparisons, suggesting a similar degree of support for the two hypotheses.

Average number of flies captured per trap (+SE) in connected (n=8), rectangular (n=12), and winged (n=12) receiver patches at the SRS. Flies were caught significantly more often in connected than rectangular patches, supporting the Traditional Corridor hypothesis, and were caught significantly more often in winged than rectangular patches, supporting the Drift Fence hypothesis
3.2 Edge experiment
Open and closed edges at the Ordway Preserve differed in the degree to which they obscured view of the white sheet in the forest (median scores of 1 and 3, respectively, χ2=4.3, df=1, P<0.05).
A total of 909 flies were captured. Flies released along closed and open edges showed significantly different patterns of dispersal (Fig. 3). Most importantly for the interpretation of results from the corridor experiment, the type of edge impacted the probability of capture (F 1,130=2.7; P=0.10; Table 1). In particular, more flies were captured at closed edge sites than at open edge sites, especially in traps located along the edge and in the field (Fig. 3b, c). This pattern confirms that flies preferred open or semi-open (edge) habitat. However, the Habitat term was non-significant (P=0.58), indicating no overall difference in capture rates among traps in the three habitats (Table 1). Trap distance from the release site was inversely related to the number of flies caught (F 4,130=15.7; P<0.001). Day of release had a significant effect on probability of fly capture, presumably due to differences among days in wind, temperature, and humidity (F 4, 130=20.1; P<0.001). The only significant interaction term was Habitat×Day (F 8, 120=9.1; P<0.001), a result of more variable captures among days in traps in the field than among days in traps along the edge and in the forest. We suggest that dispersing flies were more subject to extremes of wind, humidity, and temperature in the field than along the edge or in the forest, where habitats are more climatically buffered.

Average number of flies captured per trap (+SE) along two types of edge (open and closed) at 50-m intervals from release sites at the Ordway Reserve (Florida). Open edges (n=5) are boundaries between fields and forest with sparse understory; closed edges (n=5) are similar except for a dense understory in the forest. Traps were placed in parallel transects in three habitats: a forest, b along edge, and c field. There were no edge traps at 250 m
The contingency table analysis also suggested a non-random pattern of dispersal (χ2=3.7; df=1; P=0.056). Twenty-three percent more flies were captured in closed sites than in open sites (501 vs. 408), in large part because capture rates were higher in traps along the closed than open edges. Specifically, of all flies caught in edge traps, 59% were from closed edges versus 41% from open edges. These results suggest that during dispersal, flies more often followed closed edges than open edges.
Open edges at the Ordway Preserve were statistically indistinguishable from the edges that flies encountered during the corridor experiment at the Savannah River Site (SRS) (median scores of 3 for both sites, χ2=0.44, df=1, P>0.50).
4 Discussion
Our results support both the Traditional Corridor hypothesis and the Drift Fence hypothesis. Flies were caught more often in patches with than without corridors, even when the corridors were blind-ended. Thus, corridors can function to increase dispersal among patches of habitat even when they don’t physically connect those patches—i.e., corridors can increase connectivity of patches without connecting them. This drift fence effect was in addition to the corridors’ ability to direct the movement of flies out of the source patch, down their expanse, and into connected receiver patches. Both the traditional and drift fence effects were directly attributable to the shape and placement of the corridors, not to the area added to patches by the corridors.
We attribute the observed effect of corridors to edge-following behavior of flies, as demonstrated in the edge experiment. The higher recapture rates of flies in winged and connected patches were likely due to flies changing direction when they encountered an edge, either staying in the open habitat and following patch edge, then corridor edge into a connected receiver patch, or entering the matrix and changing direction upon encountering a blind corridor associated with a winged patch. Such mechanisms of corridor function are notoriously difficult to document, except in situations where dispersing individuals can be tracked or followed (Schultz 1998; Bowne et al. 1999; Haddad 1999; Collinge 2000; Berggren et al. 2002). In our case, we cannot be certain of fly behavior in or along corridors because we could not observe the flies and did not place traps in the corridors (for fear of influencing fly dispersal behavior via odor plumes from the traps). However, a study in northern Florida concluded that stable flies used open habitat rights-of-way as dispersal corridors through wooded areas (Williams and Rogers 1976).
The degree to which corridors affect movement of flies is probably dependent on the type of boundary between patch and matrix habitat. In the corridor experiment, the boundary was relatively open. The second-growth of our patches abutted a forest with an open understory and trees spaced 3- to 4-m apart. Flies released in the source patch dispersed in all directions, with many traversing matrix habitat into unconnected patches. This result agrees with the pattern of dispersal found in the edge experiment—flies released along edges were captured frequently in both forest and field traps. However, when flies were released along closed edges (second growth abutting a forest with dense understory), they were more likely to follow those edges than to pass through them into the forest. This difference in behavior when encountering closed versus open edges implies that if edges in the corridor experiment had been more closed, dispersing flies would have been even more likely to have followed edges to connected and winged receiver patches. Conversely, if edges in the corridor experiment had been more open, the corridor effects would likely have been much less pronounced.
How a dispersing animal responds to habitat boundaries and corridors depends not only on external attributes such as edge type but also on internal attributes such as the animal’s perceptual range (Lima and Zollner 1996; Beier and Noss 1998; Tischendorf and Fahrig 2000). Some species are deterred from entering the matrix regardless of the contrast between patch and matrix habitats, some species are easily able to traverse even hostile matrix, and others are more likely to travel through open (or “soft”) than closed (or “hard”) boundaries (Yahner 1983; Stamps et al. 1987; Rosenberg et al. 1997; Dover and Fry 2001; Ricketts 2001; Berggren et al. 2002). House flies apparently belong to this latter group—they treated closed edges as barriers more so than open edges. Thus, edge “permeability” appears to be a key factor influencing the dispersal pattern of house flies. At least three other species of Diptera appear to perceive and react in similar ways to differences in edge type. These species, all hover flies (Syrphidae) that are common in farm landscapes, are more likely to cross boundaries that are low or sparsely vegetated than other types of boundaries (Harwood et al. 1994; Wratten et al. 2003). Tabanid flies in coastal habitat in Maryland follow vegetational barriers until they encounter a gap, at which point they will cross through the barrier (Morgan and Lee 1977). Likewise, the number of butterflies that crossed boundaries separating adjacent meadows was negatively correlated with boundary height and positively correlated with boundary openness (Fry and Robson 1994).
In conclusion, our study demonstrates that habitat corridors direct the movement of flies. Although corridors clearly do not affect animal movement universally, several recent reviews have concluded that they do so commonly (Rosenberg et al. 1997; Beier and Noss 1998; Debinski and Holt 2000; Haddad et al. 2003). All acknowledge, however, that landscape connectivity is an attribute of both the species and the landscape. Thus, generalities about corridor function will remain elusive until the focus of research shifts from its current emphasis on spatial ecology to one that better embraces the behavioral ecology of dispersing individuals (Lima and Zollner 1996; Tischendorf and Fahrig 2000).
References
Aars J, Ims RA (1999) The effect of habitat corridors on rates of transfer and interbreeding between vole demes. Ecology 80:1648–1655
Anderson GS, Danielson BJ (1997) The effects of landscape composition and physiognomy on metapopulation size: the role of corridors. Landscape Ecol 12:261–271
Arnold GW, Weeldenberg JR, Steven DE (1991) Distribution and abundance of two species of kangaroo in remnants of native vegetation in the central wheatbelt of Western Australia and the role of native vegetation along road verges and fencelines as linkages. In: Saunders DA, Hobbs RJ (eds) Nature conservation 2: the role of corridors. Surrey Beatty, Chipping Norton, pp 273–280
Beier P, Noss RF (1998) Do habitat corridors provide connectivity?. Conserv Biol 12:1241–1252
Berggren A, Birath B, Kindvall O (2002) Effect of corridors and habitat edges on dispersal behavior, movement rates, and movement angles in Roesel’s Bush-cricket (Metrioptera roeseli). Conserv Biol 16:1562–1569
Bowne DR, Peles JD, Barrett GW (1999) Effects of landscape spatial structure on movement patterns of the hispid cotton rat (Sigmodon hispidus). Landscape Ecol 14:53–65
Brown JH, Kodricbrown A (1977) Turnover rates in insular biogeography—effect of immigration on extinction. Ecology 58:445–449
Collinge SK (2000) Effects of grassland fragmentation on insect species loss, colonization, and movement patterns. Ecology 81:2211–2226
Danielson BJ, Hubbard MW (2000) The influence of corridors on the movement behavior of individual Peromyscus polionotus in experimental landscapes. Landscape Ecol 15:323–331
Date EM, Ford HA, Recher HF (1991) Frugivorous pigeons, stepping stones, and needs in northern New South Wales. In: Saunders DA, Hobbs RJ (eds) Nature conservation 2: the role of corridors. Surrey Beatty, Chipping Norton, New South Wales, pp 241–245
Debinski DM, Holt RD (2000) A survey and overview of habitat fragmentation experiments. Conserv Biol 14:342–355
Dover JW, Fry GLA (2001) Experimental simulation of some visual and physical components of a hedge and the effects on butterfly behaviour in an agricultural landscape. Entomol Exp Appl 100:221–233
Dunning JB Jr, Borgella RJ, Clements K, Meffe GK (1995) Patch isolation, corridor effects, and colonization by a resident sparrow in a managed pine woodlot. Conserv Biol 9:542–550
Fry GLA, Robson WJ (1994) The effects of field margins on butterfly movement. In: Boatman N (ed) Margins: integrating agriculture and conservation. British Crop Protection Monograph No. 58, Thornton Heath Surrey, pp 111–121
Gonzalez A, Lawton JH, Gilbert FS, Blackburn TM, Evans-Freke I (1998) Metapopulation dynamics, abundance, and distribution in a microecosystem. Science 281:2045–2047
Haddad NM (1999) Corridor use predicted from behaviors at habitat boundaries. Am Nat 153:215–227
Haddad NM, Baum KA (1999) An experimental test of corridor effects on butterfly densities. Ecol Appl 9:623–633
Haddad NM, Bowne DR, Cunningham A, Danielson BJ, Levey DJ, Sargent S, Spira T (2003) Corridor use by diverse taxa. Ecology 84:609–615
Harrison S, Bruna E (1999) Habitat fragmentation and large-scale conservation: what do we know for sure? Ecography 22:225–232
Harwood RJW, Hickman JM, MacLeod A, Sherratt T, Wratten SD (1994) Managing field margins for hover flies. In: Boatman N (ed) Margins: integrating agricultue and conservation. British Crop Protection Monograph No. 58, Thornton Heath Surrey, pp 147–152
Hogsette JA (1992) New diets for production of house flies and stable flies (Diptera: Muscidae) in the laboratory. J Econ Entomol 85:2291–2294
Holt RD, Debinski DM (2003) Reflections on landscape experiments and ecological theory: Tools for the study of habitat fragmentation. In: Bradshaw GA, Marquet PA (eds) How landscapes change. Springer, Berlin Heidelberg New York, pp 201–223
Lima SL, Zollner PZ (1996) Towards a behavioral ecology of ecological landscapes. Trends Ecol Evol 11:131–135
Mech SG, Hallett JG (2001) Evaluating the effectiveness of corridors: a genetic approach. Conserv Biol 15:467–474
Morgan NO, Lee RP (1977) Vegetative barriers influence flight direction of saltmarsh greenheads. Mosq News 37:263–267
Ricketts TT (2001) The matrix matters: Effective isolation in fragmented landscapes. Am Nat 158:87–99
Rosenberg DK, Noon BR, Meslow EC (1997) Biological corridors: Form, function, and efficacy. Bioscience 47:677–687
Rosenberg DK, Noon BK, Megahan JW, Meslow EC (1998) Compensatory behavior of Ensatina eschscholtzii in biological corridors: a field experiment. Can J Zool 76:117–133
Schultz CB (1998) Dispersal behavior and its implications for reserve design in a rare Oregon butterfly. Conserv Biol 12:284–292
Simberloff D, Farr JA, Cox J, Mehlman DW (1992) Movement corridors—conservation bargains or poor investments. Conserv Biol 6:493–504
Stamps JA, Buechner M, Krishnan VV (1987) The effects of edge permeability and habitat geometry on emigration from patches of habitat. Am Nat 129:533–552
Taylor RJ (1987) The geometry of colonization: 2. Peninsulas. Oikos 48:232–237
Tewksbury JJ, Levey DJ, Haddad NM, Sargent S, Orrock JL, Weldon A, Danielson BJ, Brinkerhoff J, Damschen EI, Townsend P (2002) Corridors affect plants, animals, and their interactions in fragmented landscapes. Proc Natl Acad Sci USA 99:12923–12926
Tischendorf L, Fahrig L (2000) How should we measure landscape connectivity? Landscape Ecol 15:633–641
Williams DF, Rogers AJ (1976) Vertical and lateral distribution of stable flies in northwestern Florida. J Med Entomol 13:95–98
Wratten SD, Bowie MH, Hickman JM, Evans AM, Sedcole JR, Tylianakis JM (2003) Field boundaries as barriers to movement of hover flies (Diptera: Syrphidae) in cultivated land. Oecologia 124:605–611
Yahner RH (1983) Population dynamics of small mammals in farmstead shelterbelts. J Mammal 64:380–386
Acknowledgements
Funding was provided by the National Science Foundation (REU Supplement to DEB-9815834) and the Department of Energy-Savannah River Operations Office through the US Forest Service-Savannah River, under Interagency Agreements DE-IA09-76SR00056 and DE-IA09-00SR22188. Hazen Brown helped raise flies. John Blake, Charles Kwit, and Josh Tewksbury provided logistical support and advice. Larry Winner helped with SAS code and interpretation; we thank George Casella (University of Florida, Department of Statistics) for providing the consulting service. William Warner of Farnam Companies, Inc. provided the Captivator traps and the Musca attractant. We are especially grateful to the US Forest Service’s Research, Fire, and Forest Products staffs for their role in creating and maintaining the experimental landscapes.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Craig W. Osenberg
Rights and permissions
About this article
Cite this article
Fried, J.H., Levey, D.J. & Hogsette, J.A. Habitat corridors function as both drift fences and movement conduits for dispersing flies. Oecologia 143, 645–651 (2005). https://doi.org/10.1007/s00442-005-0023-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-005-0023-6