
Abstract
The risk of infanticide may alter foraging decisions made by females, which otherwise would have been based on nutritional requirements and forage quality and availability. In systems where meat resources are spatially aggregated in late summer and fall, female brown bears ( Ursus arctos) would be faced with a trade-off situation. The need of reproductive females to accumulate adequate fat stores would likely result in a decision to frequent salmon streams and consume the protein- and lipid-rich spawning salmon. In contrast, aggregations of bears along salmon streams would create conditions of high risk of infanticide. We investigated consumption of salmon by brown bears on Admiralty and Chichagof Islands in Southeast Alaska from 1982 to 2000 using stable isotope analysis and radiotelemetry. While nearly all males (22 of 23) consumed relatively large amounts of salmon (i.e., >10% relative contribution to seasonal diet), not all females ( n =56) did so. Five of 26 females for which we had reproductive data, occupied home ranges that had no access to salmon and thus did not consume salmon when they were mated or accompanied by young. Of females that had access to salmon streams ( n =21), all mated individuals ( n =16) had δ15N values indicative of salmon consumption. In contrast, 4 out of 16 females with cubs avoided consuming salmon altogether, and of the other 12, 3 consumed less salmon than they did when they were mated. For 11 of 21 females with access to salmon streams we had data encompassing both reproductive states. Five of those altered foraging strategies and exhibited significantly lower values of δ15N when accompanied by young than when mated, while 6 did not. Radiotelemetry data indicated that females with spring cubs were found, on average, further away from streams during the spawning season compared with females with no young, but both did not differ from males and females with yearlings and 2-year-olds. Females with young that avoided salmon streams were significantly lighter indicating that female choice to avoid consumption of salmon carries a cost that may translate to lower female or cub survivorship. The role of the social hierarchy of males and females, mating history, and paternity in affecting the risk of infanticide and foraging decisions of female brown bears merit further investigation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The risk of predation may alter foraging decisions made by individuals (Cowlishaw 1997; Diehl et al. 2000; Grand 2002; Kie 1999; Lima 1998; Luttbeg and Schmitz 2000; Sweitzer 1996). Such alteration in foraging strategies may be most pronounced for reproductive females faced with a trade-off between meeting nutritional requirements and avoiding the risk of predation, in an attempt to maximize lifetime reproductive success (Stearns 1992). In many species, females accompanied by young select habitats of lower quality, where the risk of predation may be lower or escape terrain more accessible (Barten et al. 2001; Bowyer et al. 1998; Cowlishaw 1997; Lima 1998; Rachlow and Bowyer 1998). Although superficially similar to conditions of foraging under predation risk, avoiding the risk of infanticide is further complicated by dependency on intraspecific social conditions. While animals foraging under predation risk are likely to experience predation pressures related to predator densities and habitat characters (Grand 2002; Lima and Dill 1990), infanticide may also depend on male social hierarchy (Weilgus and Bunnel 1995, 2000), male coalition size (Packer and Pusey 1984), or male-female mating history (Bartos and Madlafousek 1994). Thus, foraging decisions made by females in species experiencing risk of infanticide may have evolved under fluctuating pressures, which may lead to high variability in individual responses and have large consequences for the dynamics of populations and ecosystem processes.
Although rarely observed, infanticide in brown bears ( Ursus arctos), black bears ( U. americanus), and polar bears ( U. maritimus) is a well-documented phenomenon (Craighead et al. 1995a; Derocher and Wiig 1999; Wielgus and Bunnell 1995). Most reports describe adult and sub adult males killing young-of-the-year or yearling cubs (Craighead et al. 1995a; Dean et al. 1986; Derocher and Wiig 1999; Olson 1993; Taylor et al. 1985). Several of the reports indicate the killing of the sow in defense of her cubs, while others document the killing of young by nonsire females (Dean et al. 1986; Hessing and Aumiller 1998). Although the victim was consumed in most instances, cannibalism could not be invoked as the cause of aggression in all cases (Dean et al. 1986; Hessing and Aumiller 1998). Thus, similar to other species such as African lions ( Panthera leo; Packer and Pusey 1984) and red deer ( Cervus elaphus; Bartos and Madlafousek 1994), infanticide in bears is more likely related to male reproductive success (Swenson et al. 1997).
Weilgus and Bunnell (1995) reported that in areas with high mortality of older males, where influx of immigrant, nonsire males was high and social hierarchy was unstable, female brown bears avoided preferred habitats to reduce risk of infanticide. In areas with low male mortality, females did not demonstrate such spatial segregation (Wielgus and Bunnell 1995), suggesting that risk of infanticide may play an important role in determining availability of high-quality resources for reproductive females. Similar observations of spatial segregation due to predation on cubs by nonsire bears were made for Yellowstone grizzly bears (Mattson and Reinhart 1995). Swenson et al. (1997) described lower cub survival in a hunted population of brown bears in Sweden compared with a non-hunted population. These authors attributed the lower survivorship and resulting reduction in population growth to killing of cubs by immigrating nonsire males (Swenson et al. 1997). Similarly, Weilgus and Bunnel (2000) determined that reproduction rates in three populations of bears were consistent with expectations based on infanticide by nonsire immigrant males. In contrast, other studies indicated that dominant females had higher reproductive success (Craighead et al. 1995a), suggesting that female rather than male dominance could influence risk of infanticide and thus foraging decisions made by reproductive females.
In brown bears, the combined costs of hibernation, gestation, and lactation place large energetic demands on reproductive females (Barboza et al. 1997; Farley and Robbins 1995; Hilderbrand et al. 1999a, 1999b, 2000; Watts and Jonkel 1988). Because gestation and lactation in bears occur while the female is sequestered in a den, energy requirements for supporting a female and her offspring are derived from energy-stores accumulated prior to denning (Barboza et al. 1997; Farley and Robbins 1995). Thus, the ability of females to obtain meat resources and accumulate adequate fat stores in late summer and early fall has a large influence on gestation and lactation through the winter dormancy and the associated fast (Barboza et al. 1997; Farley and Robbins 1995). Indeed, several studies documented that reproductive success in bears is directly related to body mass in the fall (Atkinson and Ramsey 1995; Rogers 1987; Samson and Hout 1995).
Systems in which meat resources are spatially aggregated in late summer and fall, such as salmon streams ( Onchorhynchus spp.) in the Pacific Northwest (Heard 1991; Salo 1991; Sandercock 1991), create conditions where an individual female may face a trade-off situation. The need of reproductive female brown bears to accumulate adequate fat stores in late summer and early fall would likely result in a decision to frequent salmon streams and consume the protein- and lipid-rich spawning salmon. This would be especially true for mated and lactating females (i.e., females accompanied by young-of-the-year and yearlings) that would face high energetic demands in the following winter. In contrast, aggregations of bears along salmon streams would create conditions of high risk of infanticide from both males and nonsire females. Thus, females accompanied by young-of-the-year and yearlings would likely avoid aggregations of other bears along salmon streams, a decision that will result in low consumption of salmon.
In this study, we investigated consumption of salmon by brown bears on Admiralty and Chichagof Islands in Southeast Alaska from 1982 to 2000. In these areas, studies on resource selection found different selection for some habitat types by males and females (Titus et al. 1999), although both genders made high use of riparian areas in association with spawning salmon during late summer (Titus and Beier 1999). We hypothesized that females with young offspring will consume less salmon and will be found further away from salmon streams than adult males and mated or non-breeding females because females with young offspring will be more at risk from infanticidal nonsire males and females. In addition, we predicted that the same individuals would use different foraging patterns when accompanied with young offspring compared with periods when cubs are absent. We also explored the effect of female size, as a surrogate for dominance, on levels of salmon consumption in an attempt to evaluate the potential effect of dominance on decision making in reproductive brown bears.
Materials and methods
Study areas
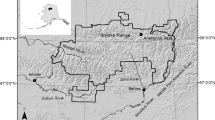
Admiralty and Chichagof Islands (57°52′N 135°18′W; Fig. 1), two of the three large northern islands of the Alexander Archipelago, are part of the Tongass National Forest. The archipelago has a maritime climate; summers are cool and wet and winters are characterized by deep snow (2,360 mm annual precipitation). The snow-free period extends from early May to early November at lower elevations. Vegetation at higher elevations is typically alpine tundra, and in lower elevations coastal, old-growth forest of Sitka spruce ( Picea sitchensis) and western hemlock ( Tsuga heterophylla) with a well-developed understory (mainly Oplopanax horridus, Vaccinium spp., Menziesia ferruginea , and Rubus spp.). New growth of grasses (Poaceae), sedges (Cyperaceae), beach peas ( Lathyrus japonicus), and skunk cabbage ( Lysichitum americanum) shoots occurs in May and June and lasts through October. Berries (blueberries, Vaccinium spp.; salmonberries, Rubus spectabilis; cloudberries, R. chamaemorus; stink current fruits, Ribes bracteosum; Pacific crab apple, Malus fusca; devil’s club fruits, Oplopanax horridus), and mushrooms ( Bolitus sp., Hygrophorus spp., Mycena spp., Omphalinea spp., and Russula spp.) become available from late July to late October. Our study area encompasses numerous streams that support an annual run of spawning Pacific salmon ( Onchorhynchus gorbuscha, O. keta , and O. kisutch) from the end of July to the end of November. Potential mammalian prey species for bears on the islands include long-tailed voles ( Microtus longicaudas) and Sitka black-tailed deer ( Odocoileus hemionus sitkensis). The intertidal zone has a rich fauna including crabs (such as Telmessus cheiragonous, Hemigrapsus oregonensis , H. nudus , Pugettia gracilis), blue mussels ( Mytilus trossulus), clams, and fishes (such as Oligocattus maculosus, Icelinus borealis , Leptocottus armatus).

Location of study areas ( shaded) on Admiralty and Chichagof Islands, southeast Alaska. USA
Brown bears in the Alexander Archipelago
Brown bears on Admiralty, Baranof, and Chichagof islands (ABC islands) of the Alexander Archipelago occur in high densities (Admiralty Island: 400–450 bears/1,000 km2; Chichagof Island: 320 bears/1,000 km2; Miller et al. 1997). Brown bears in our study areas usually den at high elevations with females entering their dens earlier (late October) and emerging later (mid-May) than males (Schoen et al. 1987). After emergence from dens bears travel to lower elevations in late April and May and begin feeding on new growth vegetation, often in estuaries, grass flats and muskegs. By mid-June many bears are found in subalpine and alpine habitats until mid-July following vegetation green-up. Mating often occurs during this period. From late-July through early September a high portion of the bear population is associated with salmon-spawning streams; some individual bears, however, do not visit salmon-spawning streams and remain at higher elevations throughout the year. By early September bears begin leaving the riparian areas, while spawning salmon remain abundant and easily accessible. Through the early fall, many of the bears are associated with avalanche chutes, a habitat type with abundant berry crops (Schoen and Beier 1990; Titus et al. 1999).
Capture of bears and collection of samples
We captured, immobilized with Telazol (7–10 mg/kg of estimated body weight; Fort Dodge Animal Health, Fort Dodge, Iowa, USA), and radio collared (Telonics, Mesa, Ariz., USA) more than 200 brown bears from 1982 through 2000. Subadult bears received surgical-tubing break-away radio collars. We captured most of the bears by darting from helicopters in rugged alpine habitats, mostly in June and early July. About 20% of the bears were captured with foot snares near a local landfill or on well-used trails along salmon-spawning streams. Capture and handling methods followed the Alaska Department of Fish and Game’s animal welfare policy to assure that bears received humane care and treatment. Over the 19-year study we believe we captured bears in representative habitats across the study areas and that there was no bias regarding capturing most bears in alpine habitats and subsequently assessing their riparian habitat use patterns. Cub survival was determined visually during re-sighting of females in years following the fitting of radio collars.
Measurements of body length, girth and neck circumference were obtained to the nearest 1 cm. Skull length and width were measured to the nearest 1 mm. Weight measurements were obtained for many of the captured bears using hanging scales to the nearest 5 kg. For bears that were not weighed, estimates were determined based on the morphometric measurements of the bear in comparison to other bears for which both weight and morphometrics were obtained and based on the experience of the tagging crew (L.R. Beier, J. Schoen, K. Titus). Blood and hair samples were collected from 56 individual female and 23 male bears.
Isotopic values of blood samples collected in June–July likely represent spring and early-summer diets, because sampling occurred 60–70 days after den emergence, a time which would allow for diet signatures to be incorporated into blood cells (Hilderbrand et al. 1996). Hair samples collected during the same period represent late summer and fall diets of the previous year because hair growth in bears occurs in late summer and fall (a period that lasts about 90 days) and hair growth is terminated when the animals enter hibernation in late October (C.T. Robbins, personal communications). Other studies of large mammals suggest that dietary isotopic values are fully expressed in hair after 10–12 weeks (Sponheimer et al. 2003).
For 22 of the females, multiple samples were obtained. These included both blood and hair samples from the same trapping occasion as well as samples from several different captures in different years. These allowed us to investigate changes in foraging patterns for the same individuals under different reproductive states. Females observed with young-of-the-year during the sampling session in spring/early-summer were defined as mated for the analysis of hair samples, because the hairs were replaced during the fall previous to capture. It is possible that these females had older offspring in toe but it is likely that these offspring did not den with the female that winter. Thus, we treated these females as if they were not accompanied by young offspring in our analysis. Females captured with yearlings and 2-year-old cubs in the spring/early summer session were defined as females with young offspring for the previous fall, which was represented in their hair samples.
Accessibility and distance to salmon streams
We located radio collared bears from a small aircraft (Kenward 1987) during daylight hours once or twice every 2 weeks throughout the year from 1990 to 2000. The location of each bear was determined with a global positioning system (GPS) unit and plotted on high-resolution, orthophoto maps (scale 1: 31,680) while circling in the aircraft above the location. Error was determined during recaptures of marked individuals and was estimated at 50–110 m. Aerial locations were digitized and transferred to geographic information system (GIS). Digital GIS maps of the study areas were obtained from the USDA Forest Service, Tongass National Forest. These maps demarcated stream segments used by anadromous fish.
Accessibility of salmon-spawning streams to each individual bear was assessed based on home range location (Minimum Convex Polygon, Kenward 1987) on the landscape. A home range that included a segment of an anadromous fish stream was considered to have salmon available. Distance to salmon stream was obtained by querying the GIS for the minimum distance to the nearest salmon stream for 38 males and 79 females ( n =117) during the spawning season in 1990 to 1995. For each bear, the average distance in each year was calculated to account for multiple locations and unequal sample sizes among bears. This resulted in a value of average distance per year. Several bears had multiple years of data (total of 181 data points), resulting in pseudo-replication. This was addressed in following analyses by blocking the analysis by bear (i.e., bear number was entered as a random factor).
Sampling food types
Samples for each season were collected for stable isotope analysis based on description of diets for brown bear in the Alexander Archipelago derived through fecal analysis (McCarthy 1989) and included: grasses; sedges; beach peas; skunk cabbage shoots and roots; berries (blueberries, salmonberries, cloudberries, stink current fruits, Pacific crab apple, devil’s club fruits); mushrooms; pink, chum, and coho salmon; long-tailed voles; Sitka black-tailed deer; crabs and blue mussels. Tissue samples of salmon and deer were obtained from encountered carcasses or obtained from fishermen and hunters (Ben-David et al. 1997a, 1997b). Vegetation and mushroom samples were collected at 100 m intervals along 11 vegetation transects (1,000 m long) from riparian to upland habitats (Ben-David et al. 1998). Additional samples of vegetation were collected at higher elevations and at beach fringe habitats. Muscle samples from long-tailed voles were collected from two companion studies (Ben-David et al. 1997a; Hanley and Barnard 1999). Each carcass was also prepared as a museum specimen (including frozen tissues) and archived at the University of Alaska Museum.
Analysis of stable isotope ratios
Tissues (clotted blood cells, hair, muscle samples, and vegetation samples) were kept frozen until preparation for determination of stable isotope ratios. Although lipid contents may deplete values of δ13C of tissues (DeNiro and Epstein 1978; Kelly 2000), samples were not defatted because clotted blood cells, hair, and vegetation contain low amounts of lipids and no difference was detected in δ13C in salmon fat and defatted muscle tissues (Ben-David 1996). Samples were dried at 60° to 70°C for 48 h and then ground to fine powder using a Wig—L Bug grinder (Crescent Dental, Chicago, Ill.). Samples of intertidal crabs were then dissolved in 95% hydrochloric acid solution to remove calcium carbonates and re-dried. Subsequently, a sub-sample (1–1.5 mg for animal tissues and 8–10 mg for plant tissues) was weighed into a miniature tin cup (4×6 mm) for combustion. We used a Europa 20/20 continuous flow isotope ratio mass spectrometer to obtain the stable isotope ratios. Each sample was analyzed in duplicate and results were accepted only if the variance between the duplicates did not exceed that of the peptone standard (δ13Cstd=−15.8, δ15Nstd=7.0, CV=0.1)
Data analysis
To determine whether male and female diets significantly differed between seasons we used 2-way MANOVA with δ13C and δ15N as the dependent variables, and season and sex as the factors as well as a sex by season interaction (Johnson and Wichern 1988). Because several samples were collected from the same individuals in different seasons and years we added individual ID as a random factor to control for pseudo-replication. Because values of δ13C in hair samples are enriched by 1–2‰ compared to muscle and blood in the same individual (Hilderbrand et al. 1996; Nakagawa et al. 1985; Tieszen and Fagre 1993), we corrected hair values relative to red blood cells by 1‰ (Hilderbrand et al 1996).
Using the program IsoSource (Phillips and Gregg 2003) we determined the cutoff point, which represented less than 10% relative contribution of salmon to the diet of bears. Although linear mixing models may be inadequate to determine relative contribution of food items to the diet of consumers (Ben-David and Schell 2001; Phillips 2001; Phillips and Koch 2002; Robbins et al. 2002), the cutoff point we derived is similar to that (6.1‰) obtained from a different model described by Hilderbrand et al. (1996; Robbins et al. 2002). To derive the cutoff point, we used diet-tissue enrichment of 2‰ for carbon when mammalian prey, avian prey, and berries were consumed, and 1‰ when salmon or invertebrates were consumed, based on results from feeding experiments in captivity on mink ( Mustela vison) and black bears (Ben-David 1996; Ben-David and Schell 2001; Hilderbrand et al. 1996). Also, based on the captive experiments we used fractionation values of 3‰ for nitrogen (Ben-David 1996; Hilderbrand et al. 1996). Before conducting the analysis, we verified that potential foods significantly differed from each other using a K nearest-neighbor randomization test (Rosing et al. 1998).
We further explored differences in diet between mated females and those accompanied by young-of-the-year and yearlings based on this cutoff point. This analysis was conducted while accounting for the availability of salmon streams based on the location of the home range on the landscape. We tested whether the isotopic values of females that adopted different foraging strategies differed between when they were mated and when accompanied by young using a Wilcoxon test of 2 related samples (Zar 1984).
We investigated the effects of sex and reproductive state on average distance to salmon streams with a one-way ANOVA with category (males, females with no cubs, females with cubs of the year, and females with yearlings and 2-year-olds) as the independent variable and distance as the dependent one with bear ID as a random factor to account for pseudo-replication. This analysis was followed by Scheffé multiple comparisons to identify the sex and reproductive states that significantly differed from each other.
We then explored whether the dominance of the female influenced her foraging patterns. Because we had no data on social interactions among our study animals we used age and size as surrogates to dominance and compared size of those females with young that consumed >10% salmon with females with young that consumed ≤10% using a Mann-Whitney test (Zar 1984). All analyses were conducted using SPSS for Windows and significance level was set at α level of 0.05.
Results
Stable isotope ratios of blood and hair samples collected from individual bears exhibited high variability (Fig. 2; n =56 females, 23 males, and overall 129 data points because of repeated sampling of several individuals). A significant change in isotope values occurred between the two seasons and genders (Fig. 2; 2-way MANOVA, overall model P <0.01, δ13C corrected, P <0.001, and δ15N, P =0.002). While no significant difference in isotopic ratios was detected between males and females in early summer (Fig. 2; 2-way MANOVA, gender P =0.08), such difference became significant in the fall (Fig. 2; 2-way MANOVA, gender P =0.01). This change was most evident from the increase in values of δ15N in samples collected from males, represented by the significant interaction between season and gender (Fig. 2; 2-way MANOVA, interaction δ13C corrected, P =0.2; and δ15N, P =0.05).

Values of δ13C and δ15N for male ( triangles) and female ( circles) brown bears on Admiralty and Chichagof Islands, southeast Alaska, between 1982 and 2000. Blood samples ( n =64 from 43 individuals) represent the spring early-summer season ( A) and hair samples ( n =65 from 37 individuals) represent late-summer fall season ( B). Mean values ± SE are given for possible food items in each season with sample sizes denoted in parentheses. Stable isotope ratios of grasses, sedges, beach pea, and roots of skunk cabbage did not significantly differ from each other ( K nearest-neighbor randomization test, P >0.3) and were pooled as one group—grasses/sedges. Similarly, voles and deer ( P =0.8), resulted in a single group, as did all species of berries ( P =0.2). Following these grouping stable isotope values of food items significantly differed from each other ( P <0.05). δ13C values of hair were corrected by 1‰ for tissue fractionation in all statistical analyses
Comparison of isotope values for mated females with those of females with young-offspring revealed that females in our study adopted three foraging strategies. Five of the 26 females for which we also had reproductive data, occupied home ranges that had no access to salmon and thus did not consume salmon when they were mated or accompanied by young (Fig. 3). Two of these females, sampled when they were neither mated nor accompanied by young, had depleted isotopic values indicative of lack of salmon in the diet regardless of reproductive state (no. 43, δ15N=1.82‰; no. 60, δ15N=2.38‰).

Values of δ13C and δ15N for female brown bears representing three foraging strategies on Admiralty and Chichagof Islands, southeast Alaska, between 1982 and 2000. Top panel ( A) represents females that consumed salmon when mated and when accompanied by young; middle panel ( B) females that changed foraging strategies; and ( C) females inhabiting home ranges without access to salmon streams. Open symbols represent data for mated females, whereas filled symbols represent females with young cubs. Circles represent individuals for which data is available in both reproductive states. Squares represent females with no repeated samples. A cutoff point for consumption of >10% of salmon corresponds with a δ15N value of 6.2‰
Of females that had access to salmon streams and reproductive information ( n =21), all mated individuals ( n =16) had δ15N values >6.2‰ (Fig. 3). In contrast, 4 out of 16 females with cubs avoided consuming salmon altogether, and of the other 12, 3 consumed less salmon than they did when they were mated (Fig. 3). For 11 of those 21 females with access to salmon streams we had data encompassing both reproductive states. Five of those altered foraging strategies and exhibited significantly lower values of δ15N when accompanied by young then when mated (Wilcoxon, P =0.043), while 6 did not (Wilcoxon, P =0.249). One individual (no. 141) had lower values of δ15N when accompanied by young than when mated in one reproductive event (1994), but no difference in another (2000; Fig. 3).
Radiotelemetry data indicated that females with spring cubs were found, on average, further away from streams during the spawning season compared with females with no young, but both did not differ from males and females with yearlings and 2-year-olds (Fig. 4; ANOVA, P =0.011). In addition, only 33% of females with young cubs were found within 1,000 m of salmon streams compared with 59% of females with no cubs (Fig. 4). Of the 79 females for which we had telemetry data, 7 were sampled both when they were mated and when they had young cubs. Six of those 7 females were found further away from salmon streams when accompanied by young-of-the-year.

Average distance to salmon stream in meters (± SE) (Top panel), and the proportion of animals found less than 1,000m from a stream (Bottom panel) for different sex and reproductive classes of brown bears on Chichagof Island, southeast Alaska, between 1990 and 1995. Different letters represent significant differences at the α=0.05 level among groups as determined by Scheffé multiple comparisons. Sample sizes are reported above bars
Cub survival to the following year was reliably established for only 6 of 21 females with access to salmon streams and foraging data. Of those 6, five consumed salmon when the cubs were young and one significantly reduced salmon consumption.
Analysis of effects of size on consumption of salmon for those females accompanied by young revealed that age, body length, girth, skull length, and skull width were not different between females with differing foraging patterns (Table 1). Circumference of neck, however, was significantly smaller in the females that consumed little salmon when accompanied by young (Table 1). Similarly body mass, measured in the following spring, was significantly lower in females that consumed little salmon when accompanied by young (Table 1).
Discussion
The upstream migration of spawning salmon was followed by a significant change in diet for brown bears in our study areas (Fig. 2). Several other studies using direct observations, fecal analysis, and stable isotope analysis documented similar patterns in coastal British Columbia and Alaska (Hilderbrand et al. 1999a, 1999b, 2000; Jacoby et al. 1999; McCarthy 1989; P. Hessing, Alaska Dept of Fish and Game, personal communication). That bears will consume large amounts of salmon when this nutrient-rich resource becomes available is not surprising in view of the high reliance of bears on stored lipids and protein during hibernation (Barboza et al. 1997; Farley and Robbins 1995). Farley and Robbins (1995) demonstrated that weight loss of adult non-breeding bears during hibernation was proportional to their metabolic body weight and ranged from less than 70 g/day to over 500 g/day. Such mass loss illustrates the importance of accumulation of fat stores prior to denning for all sex and reproductive categories of bears (Hilderbrand et al. 2000). Thus, it is unclear why several females in our study consistently avoided consumption of salmon. Whether the tendency of these females to select home ranges that do not encompass salmon streams is a learned behavior acquired from their mothers or a result of their social status is unclear, and merits further investigation.
The observation that all mated females with access to salmon streams consumed salmon was expected. Although milk consumption of captive bear cubs in the den constituted only 9% of the estimated annual milk intake, lactating female black and grizzly bears lost body mass throughout hibernation, but maintained body weight after they had resumed feeding (Farley and Robbins 1995). The rate of weight loss by denning, lactating females relative to non-lactating bears was 45% higher for black bears and 95% higher for grizzly bears (Farley and Robbins 1995). These data emphasize the high costs of gestation and lactation during hibernation for mated females and support our conclusion that mated lone females, in particular, would benefit from a diet including spawning salmon when this resource becomes available.
Of females with access to salmon streams, stable isotope values indicated that only half of those with young offspring consumed salmon, while the others significantly reduced the amounts they consumed. In addition, telemetry data indicated that females with young cubs were found significantly further away from salmon streams than females with no young. This observation is striking given that the former group likely included females with both foraging strategies. It is possible, however, that females with young offspring will have lower energetic requirements than mated females in fall. The high costs of hibernation without lactation (Barboza et al. 1997; Farley and Robbins 1995; Hilderbrand et al. 2000) preclude the possibility of nursing active, non-hibernating yearlings and 2-year-olds (C.T. Robbins, personal communications). Although no data on milk production during hibernation in those succeeding years is available, no occurrences of active mammary glands in adult hibernating bears with yearlings or 2-year-old cubs were reported (C.T. Robbins, personal communication), and lactating females stopped milk production when young-of-the-year reached 300 days of age prior to hibernation (Farley and Robbins 1995). Nonetheless, our observation that females with young that consumed small amounts of salmon had significantly lower body mass the following spring than those females with young that consumed large quantities of salmon suggests that reduction in consumption of salmon carries high energetic costs for females with young. Also, the observation that several females with young offspring made use of the available salmon and that the majority of males include large amounts of salmon in their diets, suggests the decision of females with young offspring to avoid salmon streams may be in response to factors other than nutritional requirements.
Can the risk of infanticide be implicated in the decision of female brown bears in our study to alter foraging strategies when accompanied by young? It is hard to imagine another scenario that will result in consumption of salmon by all mated females with access to salmon streams and reduction in use of this resource in half of those females when they had young. On the Kenai Peninsula, Alaska, where salmon runs are longer in duration than in our system, all females with young offspring consume large amounts of salmon (S.D. Farley and G.V. Hilderbrand, Alaska Dept of Fish and Game, personal communication). In that system, both spatial and temporal segregation appears to be the mechanism for reducing the risk of infanticide (S.D. Farley and G.V. Hilderbrand, Alaska Dept of Fish and Game, personal communication). The degree of the risk of infanticide may also differ between these two areas because the density of brown bears is higher on the ABC islands than on the Kenai Peninsula. The high density on the ABC islands combined with the fact that >60% of all brown bear radiotelemetry locations during August were <1,000 m from a salmon stream (Titus and Beier 1999) indicates that bears are highly concentrated along specific portions of streams and that the probability of encountering other bears on a regular basis is high. Finally, although we have never observed an event of infanticide in our population, on several occasions we encountered bear feces that contained hairs of brown bear cubs.
Moreover, recent studies demonstrated that in populations of bears inhabiting locations with abundant food resources, age at first reproduction and interval between litters are lower, and litter size is higher compared with those inhabiting low quality areas (Bunnell and Tait 1981; Hilderbrand et al. 1999a, 1999b; Rogers 1987; Stringham 1990a, 1990b). The high availability of salmon for brown bears on Admiralty and Chichagof Islands suggests that brown bears in our study areas should have relatively low age at first reproduction, low interval between successful litters, and high litter sizes compared with bears inhabiting low quality areas. For brown bears on the ABC islands, however, mean age at first reproduction was estimated at 8.1 years and mean litter size ranged from 1.8 to 1.9 depending on years and study area. Interval between successful litters (from weaning to weaning) was 3.9 years with a range from 3 to 6 years (Schoen and Beier 1990). In comparison, mean age at first reproduction for grizzly bears in the Yellowstone ecosystem was estimated at 6.3 years and mean litter size at 2.1 cubs per litter. Interval between successful litters was 2.9 years (Craighead et al. 1995a).
The lower than expected reproductive rates we observed in our populations may be related to lower body condition as a result of avoidance of salmon streams by some of the reproductive females. For those females, our study areas represent lower quality habitats despite the high availability of salmon, because alternative meat resources are scarce (only deer carcasses and voles). Thus, reproductive output for these females will likely be lower than that of females that consume salmon (Bunnell and Tait 1981; Hilderbrand et al. 1999a, 1999b; Rogers 1987; Stringham 1990a, 1990b). Alternatively, such low reproductive rates may be related to high densities of bears in our study areas and represent density-dependent effects. These density effects, however, are likely to result in high incidents of infanticide as encounters between females with young and nonsiring individuals will be high. In addition, our population is hunted and likely experiences lower stability of the male hierarchy and higher risk of infanticide (Weilgus and Bunnell 1995, 2000) because mostly large males are targeted. In comparison, the population in Yellowstone National Park that has a lower habitat quality but higher reproductive rates is protected. Whether the low reproductive rates in our populations are related to body condition of females as a result of avoidance of salmon streams, or a result of density effects on infanticide, or an interaction of the two merits further investigation. Determining survival of offspring of females that avoid salmon streams compared with offspring of females that frequent these habitats should be the first step in such studies. Unfortunately, our data on survivorship of cubs are meager, as most bears were not visually observed for several years after they were fitted with radio transmitters.
The factors influencing the decision of a female with young offspring to frequent or avoid salmon streams are still unclear. Weilgus and Bunnell (1995, 2000) suggested that the stability of the dominance hierarchy among males determined the spatial segregation of females with young offspring. That at least one individual in our sample adopted different foraging strategies in two separate breeding events may provide support for this hypothesis. Nonetheless, whether siring males will be less likely to kill young is unknown and merits further investigation. Analysis of paternity using DNA microsatellite analysis (Craighead et al. 1995b) concurrent with isotope analysis on consumption of salmon by the female will provide strong indirect evidence for the male dominance hypothesis.
Alternatively, it is possible that the dominance of the female determines whether she will frequent salmon streams. Our analysis of female size and age as surrogate to dominance was inconclusive potentially because of small sample sizes. Although neck circumference was significantly smaller in the females avoiding salmon streams, a parameter that may influence the females fighting ability, all other variables were not (Table 1). Smaller neck circumference and lower mass of the females avoiding salmon streams could have been a result of this decision rather than its cause. These measurements were obtained in the spring following hibernation, while the decision to avoid salmon streams was made prior to denning. Whether these females experienced smaller neck circumference and lower body mass at the time they decided to avoid salmon streams is unknown. Also, it is unclear whether size alone would accurately represent aggressiveness and thus dominance in female bears.
Our study demonstrates that the decision to consume salmon made by females foraging under the risk of infanticide may be complicated and relate not only to nutritional requirements, and quality and availability of resources, but also to intraspecific interactions. Our data suggest that females make different foraging decisions based on their reproductive state (mated or accompanied by young), as well as social conditions during specific reproductive attempts. In bears, such decisions might be made based on the stability of the male hierarchy, the female mating history, or her own social status. Alternatively, making a foraging decision could result in detrimental consequences to that reproductive effort. The role of the social hierarchy of males and females, mating history, and paternity in affecting the risk of infanticide and foraging decisions of female brown bears merit further investigation.
References
Atkinson SN, Ramsay MA (1995) The effect of prolonged fasting on the body composition and reproductive success of female polar bears ( Ursus maritimus). Funct Ecol 9:559–567
Barboza PS, Farley SD, Robbins CT (1997) Whole-body urea cycling and protein turnover during hyperphagia and dormancy in growing bears ( Ursus americanus and U. arctos). Can J Zool 75:2129–2136
Barten NL, Bowyer RT, Jenkins KJ (2001) Habitat use by female caribou: tradeoffs associated with parturition. J Wildl Manage 65:77–92
Bartos L, Madlafousek J (1994) Infanticide in a seasonal breeder: the case of red deer. Anim Behav 47:217–220
Ben-David M (1996) Seasonal diets of mink and martens: effects of spatial and temporal changes in resource abundance. PhD Thesis, University of Alaska Fairbanks. Fairbanks, Alaska
Ben-David M, Schell DM (2001) Mixing models in analyses of diet using multiple stable isotopes: a response. Oecologia 127:180–184
Ben-David M, Hanley TA, Klein DR, Schell DM (1997a) Seasonal changes in diets of coastal and riverine mink: the role of spawning Pacific salmon. Can J Zool 75:803–811
Ben-David M, Flynn RW, Schell DM (1997b) Annual and seasonal changes in diets of martens: evidence from stable isotope analysis. Oecologia 111:280–291
Ben-David M, Hanley TA, Schell DM (1998) Fertilization of terrestrial vegetation by spawning Pacific salmon: the role of flooding and predator activity. Oikos 83:47–55
Bowyer RT, Kie JG, Van Ballenberghe V (1998) Habitat selection by neonatal black-tailed deer: climate, forage or risk of predation? J Mammal 79:415–425
Bunnell FL, Tait DEN (1981) Population dynamics of bears—implications. In: Fowler CW, Smith TD (eds) Dynamics of large mammal populations. Wiley, New York, pp 75–98
Cowlishaw G (1997) Trade-offs between foraging and predation risk determine habitat use in a desert baboon population. Anim Behav 53:667–686
Craighead JJ, Sumner JS, Mitchell JA (1995a) The grizzly bears of Yellowstone: their ecology in the Yellowstone ecosystem, 1959–1992. Island, Washington, D.C.
Craighead L, Paetkau D, Reynolds HV, Vyse ER, Strobeck C (1995b) Microsatellite analysis of paternity and reproduction in Arctic grizzly bears. J Hered 86:255–261
Dean FC, Darling LM, Lierhaus AG (1986) Observations of intraspecific killing by brown bears, Ursus arctos. Can Field-Nat 100:208–211
DeNiro MJ, Epstein S (1978) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42:495–506
Derocher AE, Wiig O (1999) Infanticide and cannibalism of juvenile polar bears in Svalbard ( Ursus maritimus). Arctic 52:307–310
Diehl S, Cooper SD, Kratz KW, Nisbet RM, Roll SK, Wiseman SW, Jenkins TM Jr. (2000) Effects of multiple, predator-induced behaviors on short-term producer-grazer dynamics in open systems. Am Nat 156:293–314
Farley SD, Robbins CT (1995) Lactation, hibernation and weight dynamics of American black and grizzly bears. Can J Zool 73:2216–2222
Grand TC (2002) Alternative forms of competition and predation dramatically affect habitat selection under foraging-predation—risk trade-offs. Behav Ecol 13:280–290
Hanley TA, Barnard JC (1999) Food resources and diet composition in riparian and upland habitats for Sitka mice, Peromyscus sitkensis. Can Field-Nat 113:401–407
Heard WR (1991) Life history of pink salmon ( Oncorhynchus gorbuscha). In: Groot C, Margolis L (eds) Pacific salmon life histories. University of British Columbia Press, Vancouver, pp 118–230
Hessing P, Aumiller L (1994) Observations of conspecific predation by brown bears, Ursus arctos, in Alaska. Can Field-Nat 108:332–336
Hilderbrand GV, Farley SD, Robbins CT, Hanley TA, Titus K, Servheen C (1996) Use of stable isotopes to determine diets of living and extinct bears. Can J Zool 74:2080–2088
Hilderbrand GV, Schwartz CC, Robbins CT, Jacoby ME, Hanley TA, Arthur SM, Servheen C (1999a) The importance of meat, particularly salmon, to body size, population productivity, and conservation of North American Brown bears. Can J Zool 77:132–138
Hilderbrand GV, Jenkins SG, Schwartz CC, Hanley TA, Robbins CT (1999b) Effect of seasonal dietary meat intake on changes in body mass and composition in wild and captive brown bears. Can J Zool 77:1–8
Hilderbrand GV, Schwartz CC, Robbins CT, Hanley TA (2000) Effect of hibernation and reproductive status on body mass and condition of coastal brown bears. J Wildl Manage 64:178–183
Jacoby ME, Hilderbrand GV, Servheen C, Schwartz CC, Arthur SM, Hanley TA, Robbins CT, Michener R (1999) Trophic relationships of brown and black bears in several western North American ecosystems. J Wildl Manage 63:921–929
Johnson RA, Wichern DW (1988) Applied multivariate statistical analysis. Prentice-Hall, Englewood Cliffs
Kelly JF (2000) Stable isotopes of carbon and nitrogen in the study of avian and mammalian trophic ecology. Can J Zool 78:1–27
Kenward RE (1987) Wildlife radio tagging. Academic Press, London
Kie JG (1999) Optimal foraging and risk of predation: effects on behavior and social structure in ungulates. J Mammal 80:1114–1129
Lima SL (1998) Stress and decision-making under the risk of predation: recent developments from behavioral, reproductive, and ecological perspectives. Adv Study Behav 27:215–290
Lima SL, Dill LM (1990) Behavioral decisions made under the risk of predation: a review and prospectus. Can J Zool 68:619–640
Luttbeg B, Schmitz OJ (2000) Predator and prey models with flexible individual behavior and imperfect information. Am Nat 155:669–684
Mattson DJ, Reinhart DP (1995) Influence of cutthroat trout ( Oncorhynchus clarki) on behavior and reproduction of Yellowstone grizzly bears ( Ursus arctos), 1975–1989. Can J Zool 73:2072–2079
McCarthy TM (1989) Food habits of brown bears on northern Admiralty Island, Southeast Alaska. MSc Thesis, University of Alaska Fairbanks, Fairbanks, Alaska
Miller SD, White GC, Sellers RA, Reynolds HV, Schoen JW, Titus K, Barnes VG, Smith RB, Nelson RR, Ballard WB, Schwartz CC (1997) Brown and black bear density estimation in Alaska using radiotelemetry and replicated mark-resight techniques. Wildl Monogr 133:1–55
Nakagawa A, Kitagawa A, Asami M, Nakamura K, Schoeller DA, Slater R, Minagawa M, Kaplan IR (1985) Evaluation of isotope ratio (IR) mass spectrometry for the study of drug metabolism. Biomed Mass Spec 12:502–506
Olson T (1993) Infanticide in brown bears, Ursus arctos , at Brooks River, Alaska. Can Field-Nat 107:92–94
Packer C, Pusey AE (1984) Infanticide in carnivores. In: Hausfater G, Hrdy SB (eds) Infanticide: comparative and evolutionary perspectives. Conference on Infanticide in Animals and Man, Ithaca, 1982. Ithaca, New York, pp 31–42
Phillips DL (2001) Mixing models in analysis of diet using multiple stable isotopes: a critique. Oecologia 127:166–170
Phillips DL, Gregg JL (2003) Source partitioning using stable isotopes: coping with too many sources. Oecologia 136:261–269
Phillips DL, Koch PL (2002) Incorporating concentration dependence in stable isotope mixing models. Oecologia 130:114–125
Rachlow JL, Bowyer RT (1998) Habitat selection by Dall’s sheep: maternal tradeoffs. J Zool London 245:457–465
Robbins CT, Hilderbrand GV, Farley SD (2002) Incorporating concentration dependence in stable isotope mixing models: a response to Phillips and Koch (2002). Oecologia 133:10–13
Rogers LL (1987) Effects of food supply and kinship on social behavior, movements and population growth of black hears in northeastern Minnesota. Wildl Monogr 97:1–72
Rosing MN, Ben-David M, Barry RP (1998) Analysis of stable isotope data: a K nearest-neighbor randomization test. J Wildl Manage 62:380–388
Salo EO (1991) Life history of chum salmon ( Oncorhynchus keta) In: Groot C, Margolis L (eds) Pacific salmon life histories. University of British Columbia Press, Vancouver, pp 231–310
Samson C, Hout J (1995) Reproductive biology of female black bears in relation to body mass in early winter. J Mammal 76:68–77
Sandcrock FK (1991) Life history of coho salmon ( Oncorhynchus kisutch). In: Groot C, Margolis L (eds) Pacific salmon life histories. University of British Columbia Press, Vancouver, pp 395–446
Schoen J, Beier L (1990) Brown bear habitat preferences and brown bear logging and mining relationships in Southeast Alaska. Alaska Department of Fish and Game. Federal Aid in Wildlife Restoration Research Final Report. Study 4.17. Juneau, Alaska, USA
Schoen J, Beier LR, Lentfer JW, Johnson LJ (1987) Denning ecology of brown bears on Admiralty and Chichagof islands. Int Conf Bear Res Manage 7:293–304
Sponheimer M, Robinson T, Ayliffe L, Roeder B, Hammer J, Passey B, West A, Cerling T, Dearing D, Ehleringer J (2003) Nitrogen isotopes in mammalian herbivores: hair δ15 N values from a controlled feeding study. Int J Osteoarchaeol 13:80–87
Stearns SC (1992) The evolution of life histories. Oxford University Press, Oxford
Stringham SF (1990a) Black bear reproductive rate relative to body weight in hunted populations. In: Darling LM, Archibald WR (eds) Bears, their biology and management. Proceedings of the International Conference on Bear Management and Research. Victoria BC, February 1989. International Association for Bear Research and Management, Washington, D.C., pp 425–432
Stringham SF (1990b) Grizzly bear reproductive rate relative to body size. In: Darling LM, Archibald WR (eds) Bears, their biology and management. Proceedings of the International Conference on Bear Management and Research. Victoria BC, February 1989. International Association for Bear Research and Management, Washington, D.C., pp 433–443
Sweitzer RA (1996) Predation or starvation: consequences of foraging decisions by porcupines ( Erethizon dorsatum). J Mammal 77:1068–1077
Swenson JE, Sandegren F, Södreberg A, Bjärvall A, Franzén R, Wabakken P (1997) Infanticide caused by hunting of male bears. Nature 386:450–451
Taylor M, Larsen T, Schweinsburg RE (1985) Observations of intraspecific aggression and cannibalism in polar bears ( Ursus maritimus). Arctic 38:303–309
Tieszen LL, Fagre T (1993) Effects of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. In: Lambert JB, Grupe G (eds) Prehistoric human bone—archaeology at the molecular level. Springer, Berlin Heidelberg New York, pp 127–156
Titus K, Beier LR (1999) Suitability of stream buffers and riparian habitats for brown bears. Ursus 11:149–156
Titus K, Beier LR, Faro JR (1996) Population and habitat ecology of brown bears on Admiralty and Chichagof islands. Alaska Department of Fish and Game. Federal Aid in Wildlife Restoration Research Progress Report. Study 4.22. Juneau, Alaska, USA
Titus K, Flynn RW, Pendelton GW, Beier LR (1999) Population and habitat ecology of brown bears on Admiralty and Chichagof islands. Alaska Department of Fish and Game. Federal Aid in Wildlife Restoration Research Progress Report. Study 4.26. Juneau, Alaska, USA
Watts PD, Jonkel C (1988) Energetic cost of winter dormancy in grizzly bears. J Wildl Manage 52:654–656
Wielgus RB, Bunnell FL (1995) Tests of hypotheses for sexual segregation in grizzly bears. J Wildl Manage 59:552–560
Wielgus RB, Bunnell FL (2000) Possible negative effects of adult male mortality on female grizzly bear reproduction. Biol Conserv 93:145–154
Zar JH (1984) Biostatistical analysis. Prentice-Hall, Englewood Cliffs
Acknowledgements
This paper is dedicated to the memory of Dr. Malcolm Ramsay. Malcolm Ramsay (Professor at the University of Saskatchewan, Saskatoon) spent decades studying the ecology and behavior of polar bears in the Canadian high Arctic. We had the pleasure of discussing our ideas on trade-offs encountered by bears on numerous occasions. Malcolm provided insightful comments and constructive criticism to our work. On 20 May 2000, Malcolm Ramsay died in a helicopter crash during research out of Resolute Bay, Nunavut. The Southeast Alaska brown bear research program was initiated by J. Schoen. His foresight in collecting and archiving samples during the early years of the project made it possible to follow females through different reproductive phases and deserves our highest appreciation and gratitude. We appreciate the assistance of numerous ADF&G staff with field-work. L. Bennett and other fixed-wing pilots provided safe flying, and the long-term skilled assistance of helicopter pilot B. Englebrecht was instrumental in obtaining many of the samples used in this paper. G.V. Hilderbrand, S.D. Farley, C.T. Robbins, G.M. Blundell, and R.W. Flynn provided insightful discussions on bear ecology and stable isotope analysis. R.W. Flynn and T.V. Schumacher assisted in GIS analyses. We thank C. Restrepo for assistance in sample preparation in the laboratory. N. Haubenstock, P. Rivera, and T. Howe performed the stable isotope analysis. G.V. Hilderbrand and D. Esler, M. Festa-Bianchet and 3 anonymous reviewer provided insightful comments on earlier versions of the manuscript. This study was funded primarily by the Alaska Department of Fish and Game (ADF&G) and the Federal Aid in Wildlife Restoration Program (Studies 4.17 and 4.26). The Alaska Region of the U.S. Forest Service provided financial assistance from 1989 to 93 for the Chichagof Island portion of the study and we appreciate the support from the Hoonah Ranger District.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ben-David, M., Titus, K. & Beier, L.R. Consumption of salmon by Alaskan brown bears: a trade-off between nutritional requirements and the risk of infanticide?. Oecologia 138, 465–474 (2004). https://doi.org/10.1007/s00442-003-1442-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-003-1442-x