
Abstract
Many heat shock proteins are chaperones that help refold or degrade misfolded proteins and battle apoptosis. Because of their capacity to protect against protein misfolding, they may help keep diseases of aging at bay. A few reports have examined heat shock proteins (eg. Hsp25, Hsp60, Hsp70, and heat shock cognate 70 or Hsc70) as a function of age in the striatum and nigra. In the present study, we examined the impact of aging on Hsp25, heme oxygenase 1 (HO1 or Hsp32), Hsp40, Hsp60, Hsc70, Hsc/Hsp70 interacting protein (Hip), 78 kDa glucose-regulated protein (GRP78), Hsp90, and ubiquitinated proteins in the nigra and striatum of the female rat by infrared immunoblotting. Female animals are not typically examined in aging studies, adding further to the novelty of our study. Striatal HO1 and Hsp40 were both higher in middle-aged females than in the oldest group. Hsp60 levels were also highest in middle age in the nigra, but were highest in the oldest animals in the striatum. Striatal levels of Hsc70 and the co-chaperone Hip were lower in the oldest group relative to the youngest animals. In contrast, Hsp25 rose with advancing age in both regions. Hsp25 was also colocalized with tyrosine hydroxylase in nigral neurons. Ubiquitinated proteins exhibited a trend to rise in the oldest animals in both regions, and K48 linkage-specific ubiquitin rose significantly from 4–6 to 16–19 months in the striatum. Our study reveals a complex array of age-related changes in heat shock proteins. Furthermore, the age-related rises in some proteins, such as Hsp25, may reflect endogenous adaptations to cellular stress.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Neurodegenerative diseases are primarily diseases of protein misfolding. For this reason, they are often called proteinopathies (Morimoto 2008; Dickson 2009; Uversky 2009). Excessive protein misfolding is known as proteotoxicity and can lead to oxidative stress, protein aggregations, and cell death. However, cells have evolved an extensive network of chaperone proteins to blunt proteotoxic stress. The best-studied examples of chaperones are the heat shock proteins, which are expressed constitutively or induced by protein denaturation (Mayer and Bukau 2005; Morimoto 2008; Lanneau et al. 2010). In addition to their well-studied role in protein folding, chaperones also escort irreparably damaged proteins to the proteasome and lysosome for removal, or activate the unfolded protein response in the endoplasmic reticulum (Szegezdi et al. 2006; Kalia et al. 2010; Lanneau et al. 2010; Aridon et al. 2011; Gorbatyuk and Gorbatyuk 2013). For example, heat shock cognate 70 (Hsc70) recognizes and guides damaged proteins with a consensus motif to the lysosome for degradation by chaperone-mediated autophagy (Arias and Cuervo 2011). In addition, some heat shock proteins, such as heat shock protein 70 (Hsp70) and Hsp25 (Hsp27 in humans), are known to reduce caspase-mediated apoptosis (Arya et al. 2007; Acunzo et al. 2012). Hsp70 is highly inducible upon protein denaturation and forms one of several complexes with other chaperones such as Hsp90, which has numerous client proteins that it stabilizes, folds, activates, and assembles (Pearl and Prodromou 2006; Johnson and Brown 2009; Aridon et al. 2011). Hsp40 stimulates the ATPase and refolding activities of Hsc70 (Hohfeld 1998; Ballinger et al. 1999), and cooperates with α-synuclein to promote neurotransmitter release (Witt 2013). The Hsp60 chaperonin helps convert partially folded proteins into native proteins, but mostly acts as a ‘loner’ with few interacting partners (Clare and Saibil 2013; Saibil 2013).
Chaperones often work in conjunction with co-chaperones that help with protein triage decisions, such as the Hsc/Hsp70-interacting protein Hip (Mayer and Bukau 2005; Morimoto 2008; Kalia et al. 2010; Lanneau et al. 2010; Aridon et al. 2011). Hip stabilizes the ADP-bound state of Hsc/Hsp70, thereby promoting chaperone/substrate interactions (Mayer and Bukau 2005). Chaperones in the endoplasmic reticulum, such as glucose-regulated protein 78 (GRP78), are also integral to protein homeostasis. In unstressed cells, GRP78 associates with protein kinase RNA-like endoplasmic reticulum kinase (PERK), activating transcription factor 6 (ATF6) and inositol-requiring enzyme 1 (IRE1). Following endoplasmic reticulum stress, GRP78 dissociates from these proteins, leading to the activation of the unfolded protein response (Szegezdi et al. 2006; Walter and Ron 2011). The heat shock protein heme oxygenase 1 (HO1 or Hsp32) is also induced by cellular stress and breaks down toxic heme into the antioxidant biliverdin and the prosurvival molecule carbon monoxide (Schipper 2000; Schipper et al. 2009; Aztatzi-Santillan et al. 2010; Wu et al. 2011; Grochot-Przeczek et al. 2012). Thus, some stress-induced heat shock proteins are also important for defense against oxidative stress, which can propel and propagate proteotoxicity if left unchecked.
It is important to study heat shock proteins in the brain because they may offer considerable protection against proteinopathies and cellular stress in general (Muchowski and Wacker 2005). Indeed, levels of some heat shock proteins, including ones involved in antioxidant defense, are raised in neurodegenerative conditions, perhaps reflecting an endogenous adaptation against proteotoxic and oxidative damage. For example, levels of HO1 are increased in hippocampal and cortical neurons and astrocytes in Alzheimer’s victims (Schipper 2000; Schipper et al. 2006). HO1 is also increased in astrocytes in Parkinson’s disease and is present in Lewy bodies (Schipper et al. 1998). Some have argued that these changes are pathogenic because HO1 also breaks down heme into ferrous iron, increasing the risk for Fenton chemistry and iron toxicity (Schipper 2011). Although heme-derived iron and carbon monoxide may increase oxidative stress (Zhang and Piantadosi 1992; Desmard et al. 2007), co-induction of apoferritin synthesis is thought to limit HO1 toxicity (Dennery 2000; Ryter and Tyrrell 2000). Furthermore, the vast majority of experimental studies of HO1 manipulation demonstrate that HO1 is protective (Calabrese et al. 2009; Jazwa and Cuadrado 2010; Zhang et al. 2013). Similar to the increases in HO1, patients with mild cognitive impairment exhibit increased Hsp70 and Hsp27 levels in the inferior parietal lobule (Di Domenico et al. 2010). Hsp27 levels are also raised in the nigrostriatal pathway of Parkinson’s victims (Zhang et al. 2005). Similarly, Hsp90 levels are higher in the brain in Parkinson’s disease (Uryu et al. 2006), and Hsp27, Hsp40, Hsp60, Hsp70, and Hsp90 are all major components of Lewy bodies (Auluck et al. 2002; McLean et al. 2002; Klucken et al. 2004; Kalia et al. 2010). One might speculate that at least some of these endogenous increases in heat shock proteins help delay the onset of neurodegenerative diseases or slow their progression once they have commenced. However, such defenses may collapse with aging in end-stage disease states. Furthermore, other heat shock proteins are actually decreased in neurodegenerative disorders. For example, it has been demonstrated that Hsc70 levels are reduced within nigral neurons in Parkinson’s disease (Chu et al. 2009). In short, the heat shock protein response to neurodegenerative disorders is complex and likely to vary with the age of the individual, disease stage, oxidative stress levels, and many other variables.
Aging is the major risk factor for neurodegenerative diseases and is thought to lead to increased proteotoxic and oxidative stress. For example, the activity of the ubiquitin–proteasome and autophagic systems both decline with aging (Conconi et al. 1996; Keller et al. 2000b, c, 2002; Cuervo et al. 2005; Zeng et al. 2005). Furthermore, aging increases lipofuscin content in the brain, suggestive of increased autophagic stress (Goyal 1982b, a; Sulzer et al. 2008). Signs of oxidative damage are especially abundant in aged tissue and have helped inspire the free radical theory of aging (Sohal and Allen 1990; Harman 2006; Muller et al. 2007). Furthermore, aging is thought to exacerbate the damage associated with proteinopathies. Relative to age-matched controls, Alzheimer’s and Parkinson’s patients exhibit reductions in proteasomal activity in select brain regions (Keller et al. 2000a; McNaught and Jenner 2001; McNaught et al. 2003) and other signs of increased proteotoxicity such as hallmark protein inclusions (Braak and Braak 1995; Braak et al. 2003a, b; Thal et al. 2004). Parkinson’s and Alzheimer’s diseases are also strongly associated with oxidative stress (Alam et al. 1997; Gabbita et al. 1998; Jenner 2003; Zhu et al. 2005; Lovell and Markesbery 2007; Butterfield et al. 2010).
Although it is not clear why the aged brain exhibits increased proteotoxic and oxidative stress, heat shock proteins may be able to slow down age-related toxicity. If heat shock proteins that are reduced with aging can be identified, such molecules would be rational targets for gene therapies against age-related neurodegenerative disorders. Alternatively, if heat shock proteins are raised in some brain regions with aging, this may reflect endogenous defenses against proteotoxic stress that should be boosted by gene- or pharmacotherapies. The goal of the present study was to study the impact of aging on heat shock proteins in the ventral mesencephalon and striatum, because the dense dopaminergic projection from the substantia nigra to the caudoputamen is known to degenerate in Parkinson’s disease (Damier et al. 1999). We examined multiple heat shock proteins and co-chaperones in female rats ranging in age from 2 to 22 months. Female rats are not often used for aging studies, adding to the novelty of the present study. In our assessments, we relied on infrared immunoblotting, as this technique is highly sensitive and offers much higher resolution than X-ray film (Mathews et al. 2009; Aguilar et al. 2010; Jinwal and Dickey 2011). The infrared Odyssey imager has a bit-depth of 16 (216 or 65,536 shades of infrared) and a 4.5 log dynamic range. In contrast, X-ray film can only resolve 150 shades of gray. Noise is also low in the infrared wavelengths, thereby leading to higher signal-to-noise ratios (Patonay and Antoine 1991).
Methods
Tissue collection
Animal use was approved by the Duquesne University Institutional Animal Care and Use Committee and is in compliance with the principles outlined in the NIH Guide for the Care and Use of Laboratory Animals. We used female Sprague Dawley rats (Charles River, Wilmington, MA, USA) that were part of an in-house breeding colony to generate rat pup tissue for primary neuron cultures. As they did not survive well past the 22-month mark, all rats were sacrificed by this age. Animals were anesthetized with 5 % isoflurane immediately prior to decapitation and tissue microdissection. The boundaries of the substantia nigra and the striatum were defined according to the Paxinos rat atlas (Paxinos and Watson 1998). All animals were sacrificed within 48 h of each other and processed together. In addition, three young female rats (2–4 months old) were perfused with 4 % paraformaldehyde for immunofluorescent staining of heat shock proteins. Fixed brains were cryoprotected in 30 % sucrose in phosphate-buffered saline and cut at 40 μm on a freezing microtome (Microm HM450; Thermo Scientific, Pittsburgh, PA, USA).
Western blot analyses
Tissue samples were weighed and sonicated in 1× Cell Lysis Buffer (20 μL/mg tissue; Cat. no. 9803; Cell Signaling, Danvers, MA, USA) supplemented with protease inhibitor cocktail (Cat. no. P8340; Sigma-Aldrich, St. Louis, MO, USA) and 10 mM sodium fluoride (Leak et al. 2010). Laemmli buffer was added to this solution prior to heating. The final lysis buffer contained 1 % Triton and 0.7 % sodium dodecyl sulfate (SDS). Equal amounts of protein were separated by SDS-PAGE electrophoresis and transferred to Immobilon-FL polyvinylidene fluoride membranes or nitrocellulose membranes (EMD Millipore). Membranes were blocked with 5 % nonfat dry milk in Tris-buffered saline (TBS) or with Odyssey Block (Cat. no. 927-40000; LI-COR Bioscience, Lincoln, NE, USA) diluted 1:1 with phosphate buffered saline (PBS). Primary and secondary antibodies were prepared in 5 % bovine serum albumin (Cat. no. A30075; Research Products International, Mt. Prospect, IL, USA) in TBS with 0.1 % Tween 20 (Cat. no. BP337; Fisher Scientific, Pittsburgh, PA, USA) or Odyssey Block and PBS (1:1 ratio), also supplemented with 0.1 % Tween 20. Immunolabeled blots were visualized on an infrared Odyssey Imager and quantified with Odyssey Software (v.3; LI-COR Bioscience). All primary and secondary antibodies are listed in Tables 1 and 2. Depending upon the species of primary antibody and molecular weight of the heat shock protein, β-actin, α-tubulin, or GAPDH were used as protein loading controls. Several blots were probed for more than one protein; the original protein loading control was used for normalization in these instances.
Immunofluorescence
Free-floating coronal sections through the midbrain and striatum were stained for Hsp25, Hsp60, and HO1 in conjunction with the dopaminergic phenotypic marker tyrosine hydroxylase. Briefly, all sections were blocked in a 50 % solution of Odyssey Block diluted in PBS for 1 h. Sections were then incubated in primary antibodies overnight at 4 °C. The following day, sections were incubated in fluorescent secondary antibodies for 1 h at room temperature after a series of washes in PBS. Sections were then washed again and mounted on glass slides, coverslipped with Fluoromount-G (Cat. no. 0100-01; Southern Biotech, Birmingham, AL, USA), and viewed with confocal microscopy (Leica TCS SP2; Wetzlar, Germany). All primary and secondary antibodies used for immunofluorescence are listed in Tables 1 and 2. For each antibody, we tested multiple dilutions in order to optimize the signal-to-noise ratio, centered around dilutions recommended by the manufacturer. For every immunostaining experiment, we included the following controls: (1) exposure to all solutions except both primary antibodies and (2) exposure to one primary antibody at a time, followed by exposure to both secondary antibodies. The latter control allowed us to determine whether there was cross-reaction between the secondary and primary antibodies.
Statistical analyses
Data are presented as the mean ± standard error of the mean from 3–6 animals per age group. Animals were excluded from analysis when air bubbles during the transfer or highly fluorescent lint interfered with the western blot signal. Statistical significance was determined by one-way ANOVA followed by the least significant difference (LSD) post hoc correction (IBM SPSS Statistics, v.20; Armonk, NY, USA) when p ≤ 0.05. Trends toward statistical significance were defined as 0.05 < p ≤ 0.10.
Results
Striatum
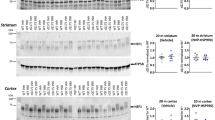
Hsp25 levels were higher in 19- to 22-month-old animals than at 2–4 months of age. There was also a trend for Hsp25 levels to be higher in 19- to 22-month-old rats than at 4–6 months of age (Fig. 1a; p = 0.072). HO1 levels were higher in 8- to 9-month-old middle-aged animals than at 2–4 months of age and were significantly lower in 16- to 19- and 19- to 22-month-old animals than at middle age (Fig. 1b). HO1 levels were also lower in the 16- to 19-month-old animals than in the 2- to 4- and 4- 6-month-old animals. Hsp40 levels in 8- to 9-month-old animals were higher than at 4–6 months of age and striatal Hsp40 expression in 19- to 22-month-old animals was lower than in all other groups except the 4- to 6-month-old rats (Fig. 1c). Hip fell from 2–4 to 4–6 months of age and showed a trend to rise in 8- to 9-month-old animals relative to the 4- to 6-month group (Fig. 1d; p = 0.059). There was also a trend towards a decrease in Hip levels from 2–4 to 16–19 months of age (p = 0.083) and Hip levels in the oldest group were significantly lower than the 2–- to 4-month-group. Hsp60 levels in 19- to 22-month-old animals were higher than all age groups except for the 4- to 6-month-old rats (Fig. 1e). Hsc70 levels were lower in 19- to 22-month-old animals than in the youngest group (Fig. 1f). Grp78 (Fig. 1g) and Hsp90 (Fig. 1i) levels were not affected by aging in this model. Ubiquitinated proteins exhibited a trend to rise in 19- to 22-month-old animals relative to the 8- to 9-month-group (Fig. 1h; p = 0.058). This antibody recognizes all ubiquitinated proteins, including ones not destined for proteasomal removal. A second antibody that specifically recognizes ubiquitin linked at Lys48 was used to measure levels of proteins targeted for the proteasomal barrel (Ciechanover and Brundin 2003; Elsasser and Finley 2005). K48 linkage-specific ubiquitinated protein levels were significantly higher at 16–19 months of age relative to the 4- to 6-month-old group. In addition, there was a statistical trend towards a difference between the 4- to 6- and 19- to 22-month-old groups (Fig. 1j; p = 0.10).

Impact of aging on heat shock proteins and ubiquitinated proteins in the caudoputamen. Whole tissue lysates of striatal tissue from female Sprague–Dawley rats were probed by infrared Western blots for the indicated proteins. GAPDH, α-tubulin, or β-actin was used as a protein loading control. Infrared signal was pseudocolored red or green. *p ≤ 0.05 versus 2–4 months old, + p ≤ 0.05 versus 4–6 months old, ∼ p ≤ 0.05 versus 8–9 months old, ^ p ≤ 0.05 versus 16–19 months old
Substantia nigra
In the substantia nigra, Hsp25 levels were higher in the 16- to 19- and 19- to 22-month old animals than in the youngest group and the 8- to 9-month-old animals (Fig. 2a). In addition, Hsp25 levels in 19- to 22-month-old animals were higher than at 4–6 months of age. There was only a trend towards a difference in Hsp25 levels between 4- to 6-month-old animals and 16- to 19-month-old animals (p = 0.074). HO1 levels exhibited a transient drop at 16–19 months of age relative to the youngest group (Fig. 2b). Hsp40 exhibited trends towards a rise in 8- to 9-month-old animals relative to the youngest group (Fig. 2c; p = 0.082) and relative to the oldest group (p = 0.054). Hip did not change significantly with aging (Fig. 2d). Hsp60 showed a rise in 8- to 9-month-old animals relative to the youngest group, and Hsp60 levels at 16–19 months and 19–22 months were lower than at 8–9 months (Fig. 2e). Hsc70 (Fig. 2f), GRP78 (Fig. 2g), and Hsp90 (Fig. 2i) levels were not impacted by aging in the nigra. Ubiquitinated proteins exhibited a trend to rise in 19- to 22-month-old animals relative to the 2- to 4-month group (Fig. 2h; p = 0.078). However, K48 linkage-specific ubiquitinated proteins were not affected by aging in the nigra (Fig. 2j).

Impact of aging on heat shock proteins and ubiquitinated proteins in the ventral midbrain. Whole tissue lysates of substantia nigra tissue from female Sprague–Dawley rats were probed by infrared Western blots for the indicated proteins. GAPDH, α-tubulin, or β-actin was used as a protein loading control. Infrared signal was pseudocolored red or green. *p ≤ 0.05 versus 2–4 months old, + p ≤ 0.05 versus 4–6 months old, ∼ p ≤ 0.05 versus 8–9 months old, ^ p ≤ 0.05 versus 16–19 months old
Although we probed for Hsp70 with multiple antibodies (see Table 1), levels of this protein were undetectable in the striatum and nigra. As a positive control, we ran cortical lysates for Hsp70, and found detectable levels in this brain region. As a result, we do not report any data on Hsp70. We also probed for the co-chaperone Hop and for several proteins related to the unfolded protein response. However, Hop was undetectable in striatum and nigra (but detectable in cortex), and PERK and phospho-PERK antibodies either led to many non-specific bands or exhibited lack of binding. Similarly, an antibody for the endoplasmic stress indicator C/EBP homologous protein (CHOP) was highly non-specific. A recent review of methods to detect the unfolded protein response also reports non-specificity in many antibodies for CHOP (Cawley et al. 2011). Indeed, the authors make the case that antibodies against unfolded protein response-related proteins are difficult to work with because of non-specificity and low abundance of these proteins, especially IRE1 and ATF6. GRP78 is more abundantly expressed and, after purchasing three separate antibodies, we finally discovered one antibody to be specific and are therefore able to report GRP78 levels (see above).
Dual immunofluorescence
Using dual-label immunofluorescence and confocal microscopy, we attempted to stain tyrosine hydroxylase+ dopamine neurons with markers against the three heat shock proteins (Hsp25, Hsp60, and HO1) that were affected by aging in the substantia nigra. Hsp60 and HO1 levels in the nigra and striatum were below the threshold for immunofluorescent detection in fixed tissue. Hsp25 immunostaining was absent from the striatum but was present at low levels in the ventral mesencephalon. Figure 3a–c illustrates Hsp25+ signal within tyrosine hydroxylase+ dopamine neurons in the substantia nigra, pars compacta of a young female rat (see arrows for examples). Omission of the primary antibodies for tyrosine hydroxylase and Hsp25, either simultaneously or one at a time, led to loss of signal. The omission of one primary antibody at a time, followed by incubation in both secondary antibodies, helped ensure that there was no cross-reaction between the primary and secondary antibodies. For example, Fig. 3d–f shows a negative control nigral section incubated only in the mouse anti-tyrosine hydroxylase primary antibody, followed by incubation in both donkey anti-mouse 488 nm and donkey anti-goat 546 nm secondary antibodies. Although it tripled the amount of work, these were essential controls because we initially used sheep and goat primary antibodies for tyrosine hydroxylase and Hsp25, respectively, and found that secondary staining in the red channel was still present even when we omitted primary antibodies against Hsp25 and that it looked exactly the same as the tyrosine hydroxylase staining in the green channel (see Tables 1, 2). This false positive appearance of ‘dual-labeling’ was the result of cross-reaction between the donkey anti-goat 546 secondary antibody with the sheep anti-tyrosine hydroxylase primary antibody, perhaps due to phylogenetic similarities between sheep and goat IgGs. Switching to mouse anti-TH antibodies eliminated the problem.

Hsp25 expression in the ventral mesencephalon. Confocal analysis of Hsp25 (red) and TH (green) immunostaining in the substantia nigra, pars compacta of a young female rat (a–c). Examples of dual-labeled neurons are indicated by white arrows. The dual-labeled neurons do not appear yellow in the merged image on the right because the Hsp25 staining was weak and the images are not overly manipulated. d–f An adjacent midbrain section that was only incubated in primary antibodies against TH, followed by exposure to the same secondary antibody solutions as in (a–c). Omission of both primary antibodies at once, or one primary antibody at a time, led to loss of fluorescent signal
Discussion
Ours is the first report of changes in striatal and nigral heat shock proteins as a function of age in female rats. In the present study, we found that middle-aged animals expressed higher levels of HO1 and Hsp40 in the striatum than old animals, and higher levels of Hsp60 in the nigra. There was a trend for Hsp40 to rise with middle age in the nigra, reflecting an inverted U-shaped curve. With advanced aging, there was a loss in HO1, Hip, Hsp40, and Hsc70 in the striatum and a loss in HO1 and Hsp60 in the nigra. In both the nigra and the striatum, Hsp25 levels were highest in the oldest group sacrificed. Taken together, these results reveal dynamic changes in heat shock proteins in response to aging. Previous studies of changes in striatal and nigral heat shock proteins as a function of age are discussed below, as are experimental caveats of the present report.
We found a striking increase in nigral and striatal Hsp25 in the oldest animals examined. Because age-related changes in Hsp25 were apparent in both the striatum and nigra, we suspected that the changes in this protein occurred within dopamine neurons. Consistent with this interpretation, confocal analyses suggest that Hsp25 was expressed within dopamine neurons of the substantia nigra, pars compacta. In addition, Hsp25 was also expressed in non-dopaminergic cells of the ventral mesencephalon. Similar to our findings in female rats, previous studies have reported age-related increases in Hsp25 in both the striatum and substantia nigra of male Fischer 433/Brown Norway rats (Gupte et al. 2010) and in whole brain extracts from mice (Dickey et al. 2009). Dickey and colleagues examined male and female mice together when they reported age-related changes in Hsp70 and Hsp25 in whole brain extracts (Dickey et al. 2009). In primates, Hsp27 is known to rise in pallidonigral spheroids with aging (Schultz et al. 2001). The precise nature of the stimulus for this age-related rise in Hsp25/27 is not known, although we speculate that it reflects a compensatory response against cellular stress. Hsp25 is an important prosurvival protein as it enhances the catalytic activity of the proteasome particle and battles apoptosis (Lanneau et al. 2010; Acunzo et al. 2012). The natural rise in Hsp25 that we and others have observed may therefore slow down age-related declines in proteasomal function and blunt stress-induced proteotoxicity. Notably, Hsp27 levels are also raised in the nigrostriatal pathway in Parkinson’s disease (Zhang et al. 2005). This response to the disease is therefore likely to be superimposed upon the age-related increase in Hsp25 that we and others have observed.
We found trends toward slight increases in ubiquitinated proteins in the striatum and nigra of aged female rats. These findings are consistent with several other reports. First, ubiquitin expression has been shown to rise in the nigra with aging in Asian Indian populations (Alladi et al. 2010). Ubiquitin-positive degenerative dendrites are also known to rise with aging in the canine substantia nigra (Uchida et al. 2003). Further, ubiquitin is known to rise in pallidonigral spheroids of aged rhesus monkeys (Schultz et al. 2001). Finally, ubiquitin-positive axonal spheroids are common in the substantia nigra of normal elderly humans (Dickson et al. 1990). As the first antibody recognizes all ubiquitinated proteins, including ones not destined for proteasomal degradation, we also measured levels of K48 linkage-specific ubiquitin. Polyubiquitin chains linked through the Lys48 residue are most often associated with proteasomal degradation, whereas polyubiquitination through Lys63 regulates signal transduction, endocytosis, kinases, and tolerance of DNA damage (Ciechanover and Brundin 2003; Elsasser and Finley 2005, 2009; Sadowski and Sarcevic 2010). The results with the K48 linkage-specific antibody revealed no net change in the midbrain, but an increase in the striatum from 4–6 months to 16–19 months and a trend towards an increase in the oldest group. Although there seemed to be a decrease from 2–4 to 4–6 months in the striatum, this change was not significant. It is possible that levels of K48 linkage-specific ubiquitinated proteins were not lowest in the youngest group because of still-ongoing developmental events requiring proteasomal processing. Many previous studies have shown that proteasomal and autophagic activities decline with aging (Conconi et al. 1996; Keller et al. 2000b, c, 2002; Cuervo et al. 2005; Zeng et al. 2005) and that lipofuscin pigments, a sign of autophagic stress, increase in the aged brain (Goyal 1982b, a, Sulzer et al. 2008). Furthermore, there is also abundant evidence of enhanced oxidative stress with aging (Sohal and Allen 1990; Harman 2006; Muller et al. 2007). Taking into account this considerable body of literature and the well-established association of heat shock proteins with cellular stress, the present findings likely reflect age-related changes in proteotoxic and/or oxidative stress levels.
We found an age-related loss of Hsc70 in the striatum of female rats. In contrast to our observations, Calabrese and colleagues have found an age-related increase in Hsc70 in the striatum and nigra of male Wistar rats (Calabrese et al. 2004). Similarly, Unno and colleagues have found an increase in Hsc70 in the striatum in 24-month-old male Wistar rats (Unno et al. 2000). However, Gupte and colleagues reported that Hsc70 was not affected by age in either the striatum or nigra of male Fisher rats (Gupte et al. 2010). In addition, we found an age-related increase in striatal Hsp60 and a rise in nigral Hsp60 in middle-aged animals. These results also conflict with data from male rats collected by Gupte and colleageus, where no change in Hsp60 in either striatum or nigra was observed (Gupte et al. 2010). All these discrepancies may reflect differences in gender or rat strain.
Similar to our study of the striatum and nigra, Dickey and colleagues did not find an age-related increase in Hsp90 in whole-brain extracts (Dickey et al. 2009). However, we observed a rise in HO1 in the striatum of middle-aged animals and a loss in the striatum and nigra in the 16- to 19-month group. Previous studies have also shown age-related declines in HO1 in the substantia nigra of male rats (Ewing and Maines 2006). Our findings of a rise in striatal HO1 in middle age contrast with a previous study by Arumugam showing a drop in HO1 in middle-aged male mice (Arumugam et al. 2010). The discrepancies between the ventral midbrain and the striatum in our study suggest that the HO1 changes may not be occurring within dopamine neurons. Unfortunately, levels of HO1 were below the threshold for immunofluorescent detection in the ventral mesencephalon, as shown by two independent antibodies raised against HO1. Hsp60 was similarly undetectable by confocal analysis. Only the more sensitive infrared immunoblotting experiments were able to detect basal levels of these two proteins. These findings may reflect low levels of Hsp60 and HO1 in the nigrostriatal pathway under non-injured, physiological conditions or that the antibodies were not suited for antigen detection on fixed tissue.
Allawi and colleagues have shown that GRP78 protein levels rise with aging in the substantia nigra of Asian Indians (Alladi et al. 2010). The authors interpret this as evidence of endoplasmic reticulum stress. However, the authors did not distinguish between male and female subjects in their analyses. We did not observe any evidence of such changes in female rodents. Many heat shock proteins other than GRP78, such as Hsp40, Hsp25, Hsp60, Hsp70, and Hsc70, have also been shown to rise in the aged central nervous system (Lee et al. 2000; Lu et al. 2004), perhaps as a defensive measure against cumulative age-related damage. As far as we can tell, there are no previous reports of the impact of aging on striatal and nigral Hip, Hsp40, and Hsp90 in either male or female mammals. Hsp40 levels in the striatum rose in middle age relative to 4- to 6-month-old rats, and dropped in old age, in an inverted, roughly U-shaped pattern. The pattern in the nigra was similarly biphasic. Furthermore, Hsp60 levels rose in middle age in the nigra in a U-shaped pattern. Biphasic responses to stress are defined as hormesis, in which there are favorable reactions to low-level stress and negative reactions to high stress (Mattson 2008; Calabrese 2010). Given the stress-inducible nature of heat shock proteins, we interpret these natural rises in heat shock proteins in middle age to reflect mild stress on protein homeostasis. Although levels of proteotoxicity probably continue to rise into old age, Hsp40 and Hsp60 defenses may eventually fail because of overwhelming stress. This dualistic response to mild versus severe stress is the fundamental basis of U-shaped stress response curves (Mattson 2008; Calabrese 2010). Raising Hsp40 and Hsp60 with gene- or pharmacotherapies may therefore be useful to ameliorate age-related declines in proteasomal function. In contrast to these U-shaped patterns, Hsp25 continued to rise into old age. The reason for these variations in heat shock protein responses to aging remain to be understood. It seems likely that the stimulus thresholds for their induction are different.
One weakness of the present set of experiments is that we did not ascertain protein activity levels. One can imagine that heat shock protein levels go up with aging in response to an age-related decline in activity. Another weakness is that, with the exception of Hsp25, we do not know the phenotype of the cells in which many of the heat shock proteins were expressed. The latter concern is actually a considerable weakness of all immunoblotting experiments, especially on heterogenous tissues such as the brain. Furthermore, the magnitude of changes within specific cell types can be masked by the presence of other abundant types of cells in which there is no change. Although one can usually identify the phenotype of cells by immunofluorescent staining, using this method on aged tissue is often confounded by the high autofluorescence of lipofuscin, a wear-and-tear pigment residue of failed lysosomal digestion (Brunk and Terman 2002; Sulzer et al. 2008). In addition, immunofluorescence on fixed tissue is anyway not a quantitative method anyway and is therefore not suitable for measuring age-related changes in heat shock proteins. Future studies using in situ hybridization for heat shock protein mRNA in phenotypically defined cellular populations are warranted to overcome some of these obstacles.
In conclusion, aging has a complex impact on heat shock protein defenses in female rats, leading to U-shaped response curves for some proteins, increases for some proteins, and declines in others. Future therapies against age-related proteotoxicity and oxidative stress should take endogenous changes in heat shock proteins into consideration.
References
Acunzo J, Katsogiannou M, Rocchi P (2012) Small heat shock proteins HSP27 (HspB1), alphaB-crystallin (HspB5) and HSP22 (HspB8) as regulators of cell death. Int J Biochem Cell Biol 44:1622–1631
Aguilar HN, Zielnik B, Tracey CN, Mitchell BF (2010) Quantification of rapid Myosin regulatory light chain phosphorylation using high-throughput in-cell Western assays: comparison to Western immunoblots. PLoS ONE 5:e9965
Alam ZI, Jenner A, Daniel SE, Lees AJ, Cairns N, Marsden CD, Jenner P, Halliwell B (1997) Oxidative DNA damage in the parkinsonian brain: an apparent selective increase in 8-hydroxyguanine levels in substantia nigra. J Neurochem 69:1196–1203
Alladi PA, Mahadevan A, Vijayalakshmi K, Muthane U, Shankar SK, Raju TR (2010) Ageing enhances alpha-synuclein, ubiquitin and endoplasmic reticular stress protein expression in the nigral neurons of Asian Indians. Neurochem Int 57:530–539
Arias E, Cuervo AM (2011) Chaperone-mediated autophagy in protein quality control. Curr Opin Cell Biol 23:184–189
Aridon P, Geraci F, Turturici G, D’Amelio M, Savettieri G, Sconzo G (2011) Protective role of heat shock proteins in Parkinson’s disease. Neurodegener Dis 8:155–168
Arumugam TV, Phillips TM, Cheng A, Morrell CH, Mattson MP, Wan R (2010) Age and energy intake interact to modify cell stress pathways and stroke outcome. Ann Neurol 67:41–52
Arya R, Mallik M, Lakhotia SC (2007) Heat shock genes - integrating cell survival and death. J Biosci 32:595–610
Auluck PK, Chan HY, Trojanowski JQ, Lee VM, Bonini NM (2002) Chaperone suppression of alpha-synuclein toxicity in a Drosophila model for Parkinson’s disease. Science 295:865–868
Aztatzi-Santillan E, Nares-Lopez FE, Marquez-Valadez B, Aguilera P, Chanez-Cardenas ME (2010) The protective role of heme oxygenase-1 in cerebral ischemia. Cent Nerv Syst Agents Med Chem 10:310–316
Ballinger CA, Connell P, Wu Y, Hu Z, Thompson LJ, Yin LY, Patterson C (1999) Identification of CHIP, a novel tetratricopeptide repeat-containing protein that interacts with heat shock proteins and negatively regulates chaperone functions. Mol Cell Biol 19:4535–4545
Braak H, Braak E (1995) Staging of Alzheimer’s disease-related neurofibrillary changes. Neurobiol Aging 16:271–278, discussion 278-284
Braak H, Del Tredici K, Rub U, de Vos RA, Jansen Steur EN, Braak E (2003a) Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging 24:197–211
Braak H, Rub U, Gai WP, Del Tredici K (2003b) Idiopathic Parkinson’s disease: possible routes by which vulnerable neuronal types may be subject to neuroinvasion by an unknown pathogen. J Neural Transm 110:517–536
Brunk UT, Terman A (2002) Lipofuscin: mechanisms of age-related accumulation and influence on cell function. Free Radic Biol Med 33:611–619
Butterfield DA, Bader Lange ML, Sultana R (2010) Involvements of the lipid peroxidation product, HNE, in the pathogenesis and progression of Alzheimer’s disease. Biochim Biophys Acta 1801:924–929
Calabrese EJ (2010) Hormesis is central to toxicology, pharmacology and risk assessment. Hum Exp Toxicol 29:249–261
Calabrese V, Scapagnini G, Ravagna A, Colombrita C, Spadaro F, Butterfield DA, Giuffrida Stella AM (2004) Increased expression of heat shock proteins in rat brain during aging: relationship with mitochondrial function and glutathione redox state. Mech Ageing Dev 125:325–335
Calabrese V, Cornelius C, Mancuso C, Barone E, Calafato S, Bates T, Rizzarelli E, Kostova AT (2009) Vitagenes, dietary antioxidants and neuroprotection in neurodegenerative diseases. Front Biosci (Elite Ed) 14:376–397
Cawley K, Deegan S, Samali A, Gupta S (2011) Assays for detecting the unfolded protein response. Methods Enzymol 490:31–51
Chu Y, Dodiya H, Aebischer P, Olanow CW, Kordower JH (2009) Alterations in lysosomal and proteasomal markers in Parkinson’s disease: relationship to alpha-synuclein inclusions. Neurobiol Dis 35:385–398
Ciechanover A, Brundin P (2003) The ubiquitin proteasome system in neurodegenerative diseases: sometimes the chicken, sometimes the egg. Neuron 40:427–446
Clare DK, Saibil HR (2013) ATP-driven molecular chaperone machines. Biopolymers 99:846–859
Conconi M, Szweda LI, Levine RL, Stadtman ER, Friguet B (1996) Age-related decline of rat liver multicatalytic proteinase activity and protection from oxidative inactivation by heat-shock protein 90. Arch Biochem Biophys 331:232–240
Cuervo AM, Bergamini E, Brunk UT, Droge W, Ffrench M, Terman A (2005) Autophagy and aging: the importance of maintaining "clean" cells. Autophagy 1:131–140
Damier P, Hirsch EC, Agid Y, Graybiel AM (1999) The substantia nigra of the human brain. II. Patterns of loss of dopamine-containing neurons in Parkinson’s disease. Brain 122(Pt 8):1437–1448
Dennery PA (2000) Regulation and role of heme oxygenase in oxidative injury. Curr Top Cell Regul 36:181–199
Desmard M, Boczkowski J, Poderoso J, Motterlini R (2007) Mitochondrial and cellular heme-dependent proteins as targets for the bioactive function of the heme oxygenase/carbon monoxide system. Antioxid Redox Signal 9:2139–2155
Di Domenico F, Sultana R, Tiu GF, Scheff NN, Perluigi M, Cini C, Butterfield DA (2010) Protein levels of heat shock proteins 27, 32, 60, 70, 90 and thioredoxin-1 in amnestic mild cognitive impairment: an investigation on the role of cellular stress response in the progression of Alzheimer disease. Brain Res 1333:72–81
Dickey C, Kraft C, Jinwal U, Koren J, Johnson A, Anderson L, Lebson L, Lee D, Dickson D, de Silva R, Binder LI, Morgan D, Lewis J (2009) Aging analysis reveals slowed tau turnover and enhanced stress response in a mouse model of tauopathy. Am J Pathol 174:228–238
Dickson DW (2009) Neuropathology of non-Alzheimer degenerative disorders. Int J Clin Exp Pathol 3:1–23
Dickson DW, Wertkin A, Kress Y, Ksiezak-Reding H, Yen SH (1990) Ubiquitin immunoreactive structures in normal human brains. Distribution and developmental aspects. Lab Invest 63:87–99
Elsasser S, Finley D (2005) Delivery of ubiquitinated substrates to protein-unfolding machines. Nat Cell Biol 7:742–749
Ewing JF, Maines MD (2006) Regulation and expression of heme oxygenase enzymes in aged-rat brain: age related depression in HO-1 and HO-2 expression and altered stress-response. J Neural Transm 113:439–454
Finley D (2009) Recognition and processing of ubiquitin-protein conjugates by the proteasome. Annu Rev Biochem 78:477–513
Gabbita SP, Lovell MA, Markesbery WR (1998) Increased nuclear DNA oxidation in the brain in Alzheimer’s disease. J Neurochem 71:2034–2040
Gorbatyuk MS, Gorbatyuk OS (2013) The molecular chaperone GRP78/BiP as a therapeutic target for neurodegenerative disorders: a mini review. J Genet Syndr Gene Ther 4
Goyal VK (1982a) Lipofuscin pigment accumulation in human brain during aging. Exp Gerontol 17:481–487
Goyal VK (1982b) Lipofuscin pigment accumulation in the central nervous system of the mouse during aging. Exp Gerontol 17:89–94
Grochot-Przeczek A, Dulak J, Jozkowicz A (2012) Haem oxygenase-1: non-canonical roles in physiology and pathology. Clin Sci (Lond) 122:93–103
Gupte AA, Morris JK, Zhang H, Bomhoff GL, Geiger PC, Stanford JA (2010) Age-related changes in HSP25 expression in basal ganglia and cortex of F344/BN rats. Neurosci Lett 472:90–93
Harman D (2006) Alzheimer’s disease pathogenesis: role of aging. Ann NY Acad Sci 1067:454–460
Hohfeld J (1998) Regulation of the heat shock conjugate Hsc70 in the mammalian cell: the characterization of the anti-apoptotic protein BAG-1 provides novel insights. Biol Chem 379:269–274
Jazwa A, Cuadrado A (2010) Targeting heme oxygenase-1 for neuroprotection and neuroinflammation in neurodegenerative diseases. Curr Drug Targets 11:1517–1531
Jenner P (2003) Oxidative stress in Parkinson’s disease. Ann Neurol 53(Suppl 3):S26–S36, discussion S36-28
Jinwal UK, Dickey CA (2011) Cell-based assays for regulators of tau biology. Methods Mol Biol 670:93–108
Johnson JL, Brown C (2009) Plasticity of the Hsp90 chaperone machine in divergent eukaryotic organisms. Cell Stress Chaperones 14:83–94
Kalia SK, Kalia LV, McLean PJ (2010) Molecular chaperones as rational drug targets for Parkinson’s disease therapeutics. CNS Neurol Disord Drug Targets 9:741–753
Keller JN, Hanni KB, Markesbery WR (2000a) Impaired proteasome function in Alzheimer’s disease. J Neurochem 75:436–439
Keller JN, Hanni KB, Markesbery WR (2000b) Possible involvement of proteasome inhibition in aging: implications for oxidative stress. Mech Ageing Dev 113:61–70
Keller JN, Huang FF, Markesbery WR (2000c) Decreased levels of proteasome activity and proteasome expression in aging spinal cord. Neuroscience 98:149–156
Keller JN, Gee J, Ding Q (2002) The proteasome in brain aging. Ageing Res Rev 1:279–293
Klucken J, Shin Y, Masliah E, Hyman BT, McLean PJ (2004) Hsp70 reduces alpha-synuclein aggregation and toxicity. J Biol Chem 279:25497–25502
Lanneau D, Wettstein G, Bonniaud P, Garrido C (2010) Heat shock proteins: cell protection through protein triage. ScientificWorldJournal 10:1543–1552
Leak RK, Castro SL, Jaumotte JD, Smith AD, Zigmond MJ (2010) Assaying multiple biochemical variables from the same tissue sample. J Neurosci Methods 191:234–238
Lee CK, Weindruch R, Prolla TA (2000) Gene-expression profile of the ageing brain in mice. Nat Genet 25:294–297
Lovell MA, Markesbery WR (2007) Oxidative DNA damage in mild cognitive impairment and late-stage Alzheimer’s disease. Nucleic Acids Res 35:7497–7504
Lu T, Pan Y, Kao SY, Li C, Kohane I, Chan J, Yankner BA (2004) Gene regulation and DNA damage in the ageing human brain. Nature 429:883–891
Mathews ST, Plaisance EP, Kim T (2009) Imaging systems for westerns: chemiluminescence vs. infrared detection. Methods Mol Biol 536:499–513
Mattson MP (2008) Hormesis defined. Ageing Res Rev 7:1–7
Mayer MP, Bukau B (2005) Hsp70 chaperones: cellular functions and molecular mechanism. Cell Mol Life Sci 62:670–684
McLean PJ, Kawamata H, Shariff S, Hewett J, Sharma N, Ueda K, Breakefield XO, Hyman BT (2002) TorsinA and heat shock proteins act as molecular chaperones: suppression of alpha-synuclein aggregation. J Neurochem 83:846–854
McNaught KS, Jenner P (2001) Proteasomal function is impaired in substantia nigra in Parkinson’s disease. Neurosci Lett 297:191–194
McNaught KS, Belizaire R, Isacson O, Jenner P, Olanow CW (2003) Altered proteasomal function in sporadic Parkinson’s disease. Exp Neurol 179:38–46
Morimoto RI (2008) Proteotoxic stress and inducible chaperone networks in neurodegenerative disease and aging. Genes Dev 22:1427–1438
Muchowski PJ, Wacker JL (2005) Modulation of neurodegeneration by molecular chaperones. Nature reviews. Neuroscience 6:11–22
Muller FL, Lustgarten MS, Jang Y, Richardson A, Van Remmen H (2007) Trends in oxidative aging theories. Free Radic Biol Med 43:477–503
Patonay G, Antoine M (1991) Near-infrared fluorogenic labels: new approach to an old problem. Anal Chem 63:321A–327A
Paxinos G, Watson C (1998) The rat brain in stereotaxic coordinates. Academic, San Diego
Pearl LH, Prodromou C (2006) Structure and mechanism of the Hsp90 molecular chaperone machinery. Annu Rev Biochem 75:271–294
Ryter SW, Tyrrell RM (2000) The heme synthesis and degradation pathways: role in oxidant sensitivity. Heme oxygenase has both pro- and antioxidant properties. Free Radic Biol Med 28:289–309
Sadowski M, Sarcevic B (2010) Mechanisms of mono- and poly-ubiquitination: ubiquitination specificity depends on compatibility between the E2 catalytic core and amino acid residues proximal to the lysine. Cell Div 5:19
Saibil H (2013) Chaperone machines for protein folding, unfolding and disaggregation. Nature reviews. Mol Cell Biol 14:630–642
Schipper HM (2000) Heme oxygenase-1: role in brain aging and neurodegeneration. Exp Gerontol 35:821–830
Schipper HM (2011) Heme oxygenase-1 in Alzheimer disease: a tribute to Moussa Youdim. J Neural Transm 118:381–387
Schipper HM, Liberman A, Stopa EG (1998) Neural heme oxygenase-1 expression in idiopathic Parkinson’s disease. Exp Neurol 150:60–68
Schipper HM, Bennett DA, Liberman A, Bienias JL, Schneider JA, Kelly J, Arvanitakis Z (2006) Glial heme oxygenase-1 expression in Alzheimer disease and mild cognitive impairment. Neurobiol Aging 27:252–261
Schipper HM, Song W, Zukor H, Hascalovici JR, Zeligman D (2009) Heme oxygenase-1 and neurodegeneration: expanding frontiers of engagement. J Neurochem 110:469–485
Schultz C, Dick EJ, Cox AB, Hubbard GB, Braak E, Braak H (2001) Expression of stress proteins alpha B-crystallin, ubiquitin, and hsp27 in pallido-nigral spheroids of aged rhesus monkeys. Neurobiol Aging 22:677–682
Sohal RS, Allen RG (1990) Oxidative stress as a causal factor in differentiation and aging: a unifying hypothesis. Exp Gerontol 25:499–522
Sulzer D, Mosharov E, Talloczy Z, Zucca FA, Simon JD, Zecca L (2008) Neuronal pigmented autophagic vacuoles: lipofuscin, neuromelanin, and ceroid as macroautophagic responses during aging and disease. J Neurochem 106:24–36
Szegezdi E, Logue SE, Gorman AM, Samali A (2006) Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep 7:880–885
Thal DR, Del Tredici K, Braak H (2004) Neurodegeneration in normal brain aging and disease. Sci Aging Knowl Environ 2004:pe26
Uchida K, Kihara N, Hashimoto K, Nakayama H, Yamaguchi R, Tateyama S (2003) Age-related histological changes in the canine substantia nigra. J Vet Med Sci Jpn Soc Vet Sci 65:179–185
Unno K, Asakura H, Shibuya Y, Kaiho M, Okada S, Oku N (2000) Increase in basal level of Hsp70, consisting chiefly of constitutively expressed Hsp70 (Hsc70) in aged rat brain. J Gerontol A 55:B329–B335
Uryu K, Richter-Landsberg C, Welch W, Sun E, Goldbaum O, Norris EH, Pham CT, Yazawa I, Hilburger K, Micsenyi M, Giasson BI, Bonini NM, Lee VM, Trojanowski JQ (2006) Convergence of heat shock protein 90 with ubiquitin in filamentous alpha-synuclein inclusions of alpha-synucleinopathies. Am J Pathol 168:947–961
Uversky VN (2009) Intrinsic disorder in proteins associated with neurodegenerative diseases. Front Biosci J Virtual Libr 14:5188–5238
Walter P, Ron D (2011) The unfolded protein response: from stress pathway to homeostatic regulation. Science 334:1081–1086
Witt SN (2013) Molecular chaperones, alpha-synuclein, and neurodegeneration. Mol Neurobiol 47:552–560
Wu ML, Ho YC, Yet SF (2011) A central role of heme oxygenase-1 in cardiovascular protection. Antioxid Redox Signal 15:1835–1846
Zeng BY, Medhurst AD, Jackson M, Rose S, Jenner P (2005) Proteasomal activity in brain differs between species and brain regions and changes with age. Mech Ageing Dev 126:760–766
Zhang J, Piantadosi CA (1992) Mitochondrial oxidative stress after carbon monoxide hypoxia in the rat brain. J Clin Invest 90:1193–1199
Zhang Y, James M, Middleton FA, Davis RL (2005) Transcriptional analysis of multiple brain regions in Parkinson’s disease supports the involvement of specific protein processing, energy metabolism, and signaling pathways, and suggests novel disease mechanisms. Am J Med Genet B 137B:5–16
Zhang M, An C, Gao Y, Leak RK, Chen J, Zhang F (2013) Emerging roles of Nrf2 and phase II antioxidant enzymes in neuroprotection. Prog Neurobiol 100:30–47
Zhu X, Lee HG, Casadesus G, Avila J, Drew K, Perry G, Smith MA (2005) Oxidative imbalance in Alzheimer’s disease. Mol Neurobiol 31:205–217
Acknowledgments
R.K.L wrote the manuscript with A.M.G. A.M.G., S.H., D.B.P., and J.M.P. performed the experiments and analyzed the data. We are grateful to Deb Willson, Mary Caruso, and Jackie Farrer for outstanding administrative support and to Dr. John Pollock for use of his confocal microscope. Funding was provided by the Mylan School of Pharmacy at Duquesne University.
Author information
Authors and Affiliations
Corresponding author
Additional information
A.M. Gleixner and S.H. Pulugulla did equivalent work.
Rights and permissions
About this article
Cite this article
Gleixner, A.M., Pulugulla, S.H., Pant, D.B. et al. Impact of aging on heat shock protein expression in the substantia nigra and striatum of the female rat. Cell Tissue Res 357, 43–54 (2014). https://doi.org/10.1007/s00441-014-1852-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-014-1852-6