
Abstract
Free-living amoebae of the genus Acanthamoeba are causative agents of Acanthamoeba keratitis and amoebic encephalitis in humans, both of which are serious infections. The ability to produce proteases is one of the factors involved in the pathogenesis of Acanthamoeba infections. The aim of this study was to evaluate the secreted proteases of six Acanthamoeba strains from distinct genotypes (T1, T2, T4 and T11) maintained in prolonged axenic culture and following three successive passages in Madin-Darby Canine Kidney (MDCK) cells. Conditioned medium was obtained from cultures before and after interaction with the MDCK monolayers, resolved in SDS-PAGE containing gelatine, then subjected to quantitative azocasein assays. Zymography profiles varied between the strains, with the predominant proteases found to be serine-type proteases from 49 to 128 kDa. A T1 genotype strain isolated from dust showed quantitatively higher protease secretion compared to the other strains. No changes were detected in the zymography profiles of MDCK-interacted cultures compared to long-term axenic cultures. Two strains presented lower proteolytic activity post-MDCK interaction, while the remaining strains presented similar values before and after MDCK passages. In conclusion, this study confirms the predominance of serine-type protease secretion by Acanthamoeba, with distinct profiles presented by the different strains and genotypes studied. Also, interaction of trophozoites with MDCK cells did not alter the zymography pattern.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Amoebae of the genus Acanthamoeba are free-living organisms that are ubiquitously distributed in nature and are therefore in frequent contact with humans. In most cases this contact is innocuous; however, Acanthamoeba species can cause potentially blinding or fatal infections. A painful, progressive corneal disease caused by this protozoan, known as Acanthamoeba keratitis (AK), is generally associated with contact lens use, usually due to poor hygiene or exposure to amoebas while swimming or showering while wearing contact lenses (Page and Mathers 2013). Other clinical presentations of acanthamoebiasis include skin lesions and lung infections that progress to involvement of the central nervous system, culminating in granulomatous amoebic encephalitis (GAE). GAE is a severe disease with a high mortality rate (higher than 90%) that occurs predominantly in patients presenting chronic or immunosuppressive diseases (Khan 2009; Marciano-Cabral and Cabral 2003).
Proteases are considered an important virulence factor of many pathogens, including Acanthamoeba (Khan 2006). Adhesion to host cells through glycoprotein-lectin interactions constitutes the first step in AK, which is followed by protease secretion and a subsequent cytopathic effect (Hurt et al. 2003; Clarke and Niederkorn 2006). In GAE, proteases play an important role in the disruption of the blood-brain barrier (Alsam et al. 2005). Acanthamoeba predominantly produces serine proteases but can also secrete cysteine and metalloproteases, which present variable patterns between distinct isolates (Alfieri et al. 2000; Khan et al. 2000; Khan 2006; Serrano-Luna et al. 2006). Some studies have reported that clinical isolates release higher quantities of extracellular proteases than environmental isolates (Khan et al. 2000; Kim et al. 2006; Lorenzo-Morales et al. 2015). A study by Koehsler et al. (2009) demonstrated that a strain which was previously interacted with HEp-2 cell monolayers presented higher protease activity than the corresponding culture under prolonged axenic conditions. Another report indicated that the production of a serine protease known as MIP133 is stimulated after contact of the amoeba with Corynebacterium xerosis, a mannose-rich bacterium that is present in the ocular flora (Alizadeh et al. 2005). Thus, host-related factors seem to be involved in the modulation of protease secretion by Acanthamoeba.
Among the 20 genotypes of Acanthamoeba that have been identified (T1 to T20) (Stothard et al. 1998; Nuprasert et al. 2010; Corsaro et al. 2015), the T4 genotype is most commonly associated with GAE and AK (Maciver et al. 2013). Most previous studies that have focused on protease characterisation have evaluated strains belonging to the T4 genotype (Hurt et al. 2003; Khan et al. 2002; Koehsler et al. 2009; Mahdavi Poor et al. 2017). Protease characterisation of genotypes other than T4 is less common. Thus, the aim of this study was to compare the qualitative and quantitative profiles of secreted proteases of six Acanthamoeba strains of distinct genotypes (T1, T2, T4 and T11). In addition, trophozoites in prolonged axenic culture were compared to trophozoites interacted with MDCK monolayers to determine whether contact with host cells can alter the protease profile.
Materials and Methods
Acanthamoeba strains
Six Acanthamoeba strains belonging to the genotypes T1, T2, T4 and T11 were used. Two of them (LG and ALX) were isolated from AK cases, and four (AP4, R2P5, AR14 and AR15) had an environmental origin. General details of the isolates, including their pathogenic characteristics, are outlined in Table1. Acanthamoeba cultures were maintained axenically in PYG medium (20-g/L proteose peptone, 1-g/L yeast extract, 0.1-M glucose, 4-mM MgSO4, 0.4-mM CaCl2, 3.4 -mM sodium citrate, 0.05-mM Fe(NH4)2(SO4)2 and 2.5 mM of each of Na2HPO4 and KH2PO4) supplemented with 10% foetal bovine serum (FBS; Cultilab, Campinas, SP, Brazil) and 20-μg/L enrofloxacin (Baytril, Bayer) at 32 °C.
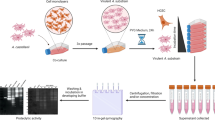
Acanthamoeba passages in MDCK monolayers
Monolayers of the established Madin-Darby Canine Kidney (MDCK, ATCC CCL34) cell line were cultured in 25-cm2 cell culture flasks (Sarstedt, Nümbrecht, Germany) in Dulbecco’s modified Eagle’s medium (DMEM; Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% FBS (Cultilab, Campinas, SP, Brazil) and antibiotics (10-U/mL penicillin, 10-μg/mL streptomycin, 0.025-mg/mL fungizone) in a 5% CO2 atmosphere at 37 °C. The cells were trypsinized and quantified in a Neubauer chamber. Cells were seeded into a 24-well plate (Sarstedt, Nümbrecht, Germany) at a density of 2 × 105 cells per well along with 0.5 mL of complete DMEM. The plate was incubated for 24 h in a 5% CO2 atmosphere at 37 °C to allow cells to adhere. Acanthamoeba trophozoites in the exponential growth phase were washed by centrifugation in phosphate-buffered saline, quantified and then suspended in complete DMEM. Then, 500 μL of each sample (containing 1 × 105 trophozoites) was added into each well (2:1; cell:trophozoite ratio), and the plate was incubated at 37 °C in 5% CO2 atmosphere for 24 h. The DMEM medium from each well was gently aspirated and released three times by pipetting to detach the trophozoites, and then they were transferred to 1.5-mL microtubes. After centrifugation at 400 × g for 5 min, the supernatant was removed, and the sediment with trophozoites was transferred to a plate with fresh MDCK cells, prepared under the same conditions as previously described. This process was repeated one more time for a total of three cocultivations. After the last passage, trophozoites were transferred to 25-cm2 flasks containing complete PYG medium (with antibiotic and FBS) and incubated at 32 °C.
Conditioned medium
To obtain conditioned medium for zymography and azocasein assays, trophozoites from prolonged axenic cultures and those from MDCK-interacted cultures were prepared after no more than four passages in PYG. Trophozoites in the exponential growth phase were quantified in a Neubauer chamber and centrifuged at 500 × g for 10 min, and then 1 × 107 trophozoites were transferred to culture flasks (Sarstedt, Nümbrecht, Germany) with a final volume of 5 mL of PYG medium without serum. The flasks were maintained at 32 °C for 24 h, and then the supernatant was collected and centrifuged for 10 min at 500 × g. The new supernatant was filtered through a sterile 0.22-μm membrane (Millex®; Merck Millipore, Darmstadt, Germany) and stored in a freezer at −80 °C.
Zymography assays
Sodium dodecyl sulphate-polyacrylamide gel electrophoresis containing 1-mg/mL gelatine was used to obtain the profile of extracellular proteases from Acanthamoeba before and after interaction with MDCK cell monolayers. The procedures were based on a previously described protocol (Alfieri et al. 2000). Briefly, 5 μL of conditioned medium was diluted (1:2) with electrophoresis sample loading buffer and applied to the gels. After electrophoresis, gels were soaked in 2% Triton X-100 solution for 60 min, incubated overnight in developing buffer (50 mM Tris with 10 mM CaCl2, pH 7.4) at 37 °C, and then stained with Coomassie Brilliant Blue. To partially classify the proteases, samples of conditioned medium were pretreated with 10-mM ethylenediaminetetraacetic acid (EDTA, a metalloprotease inhibitor), 1-mM phenylmethylsulfonylfluoride (PMSF, a serine proteases inhibitor) and 10-μM E64 (a cysteine proteinase inhibitor) for 30 min before the run.
Azocasein assay
Protease activity in the conditioned medium was determined by a colorimetric method (Khan et al. 2000). Briefly, 50 μL of 2% azocasein was incubated with 25 μL of conditioned medium in pH 7.5 buffer for 3 h. The reaction was stopped with 10% trichloroacetic acid (TCA). After 15 min, the mixture was centrifuged at 10,000 × g for 15 min. The supernatant was transferred to a 96-well plate containing 100 μL of 1 M NaOH, and absorbance was determined at 440 nm. The values were converted to units of protease activity (U), as previously described (Alfieri et al. 2000). One-way ANOVA was used to compare protease activities of the strains. The differences between the values before and after MDCK interaction were evaluated with a Mann-Whitney test (non-normally data) or a paired t-test (normally distributed data). A P value of <0.05 indicated that the differences were statistically significant.
Results
Conditioned medium from all strains of Acanthamoeba following prolonged cultivation presented a protease pattern characterised by one to three bands between 100 and 150 kDa and one to three smaller bands from 49 to 65 kDa (Fig.1, left column gels, channel 1). R2P5 samples exhibited higher activity than other isolates, needing to be diluted to obtain visually defined bands (Fig.1, channels 1a and 1b). A similar zymographic pattern was seen in strains R2P5, ALX and AR15, which included bands at 47, 59 and 108 kDa. The other strains showed unique band patterns, with AP4 presenting main bands at 47, 108, 123 and 150 kDa; LG with bands at 47, 50, 59, 108 and 123 kDa; and AR14 with bands at 47, 59, 100 and 130 kDa.

Proteolytic activity of conditioned medium from six Acanthamoeba isolates under conditions of prolonged cultivation (gels along the left side) and after interaction with MDCK cells (gels along the right side). Channels indicated by number 1 represent original conditioned medium samples, with 1a and 1b corresponding to R2P5-conditioned medium diluted 1:10 and 1:20. Channels indicated by numbers 2, 3 and 4 correspond to samples previously treated with the inhibitors EDTA (10 mM), PMSF (1 mM) and E64 (50 mM), respectively. The molecular weight in kDa is indicated on the edge of each gel
Proteases were totally or strongly inhibited by PMSF (Fig.1, left column gels, channel 3), while E64 had no effect on the original band profile (channel 4). The treatment of samples with EDTA (channels 2) did not alter the number of bands, but seemed to induce faster migration of proteases with smaller molecular weights (49 to 65 kDa).
Conditioned medium from MDCK-interacted cultures showed similar protease profiles to those from prolonged axenic cultures (Fig.1, right column gels, channel 1). All of them were also totally or strongly inhibited by PMSF (channel 3), as shown in gels corresponding to prolonged cultures. No modifications in EDTA or E64 inhibition profiles were seen in relation to gels corresponding to prolonged cultures (channels 2 and 4, respectively).
The azocasein quantitative assay indicated activities between 0.39 and 2.44 U/mL (Fig.2). Prior to MDCK cell interaction, the R2P5 strain (genotype T1) had a protease activity about two to six times higher than other strains. Following R2P5, strains ALX and AR14 exhibited the second highest values of protease activity. However, ALX activity was significantly higher only than LG and AR15, while AR14 was higher only than LG. After interaction with MDCK cells, ALX and R2P5 exhibited significantly lower activity than their corresponding long-term cultures, while the other strains showed similar protease activities before and after MDCK cell interaction (Fig.2).

Quantification of the proteolytic activity by azocasein colorimetric assay of conditioned medium obtained from six strains of Acanthamoeba. Significant differences (P < 0.05) indicating higher protease activity as follows: (a) R2P5 versus the other five strains, (b) ALX versus LG and AR15, (c) AR14 versus LG and (d) prolonged culture versus MDCK-interacted culture
Discussion
All Acanthamoeba species, regardless of their supposed pathogenicity, produce proteases as a biological process to degrade food in the environment (Khan 2006). However, Acanthamoeba proteases have been identified as an important determinant of pathogenicity, as clinically relevant samples show higher protease production than those that do not cause infection (Khan et al. 2000; Khan 2006; Koehsler et al. 2009). Several studies have characterised Acanthamoeba proteases; however, most have focused on T4 genotype strains (Alves et al. 2016; Magliano et al. 2009; Mahdavi et al. 2017) or strains with an unknown genotype (de Souza Carvalho et al. 2011; Khan et al. 2000; Serrano-Luna et al. 2006). In the present study, strains from clinical and environmental sources belonging to distinct genotypes were evaluated to assess the profile of secreted proteases. The protease profiles varied among the strains, and partial characterisation indicated serine protease activity, which was confirmed to be the predominant type in the repertoire of proteolytic enzymes described for Acanthamoeba (Dudley et al. 2008; Khan 2006; Serrano-Luna et al. 2006; Sissons et al. 2005). Faster migration of lower molecular weight bands was observed in samples treated with EDTA, potentially due to conformational changes induced by metal ion chelation, which could affect enzyme structure (Szajn and Csopak 1977).
A protease molecular weight of approximately 107 kDa, previously reported as a possible pathogenicity marker (Khan et al. 2000), was observed in all strains including AP4 (T2) and AR15 (T11), which are considered to have low pathogenicity (Da Rocha-Azevedo and Costa e Silva-Filho 2007; Duarte et al. 2013). Also, a previous observation that pathogenic Acanthamoeba presents quantitatively higher protease activity (Khan et al. 2000; Lorenzo-Morales et al. 2005; Mattana et al. 2002) was not confirmed under the present conditions, as the supposedly pathogenic LG, a T4 genotype strain obtained from a keratitis case, did not present significantly higher protease activity than the low pathogenicity T2 and T11 strains. These observations confirmed the complexity of protease profiles among Acanthamoeba strains, as previously mentioned by Serrano-Luna et al. (2006), making it difficult to determine a pathogenicity criterion for protease assays.
Nevertheless, remarkable protease activity seems to be a feature of some strains, as observed for R2P5 (genotype T1). For this strain, the conditioned medium needed to be diluted in order to visualise separated bands in the zymography assay. This strain was isolated from an environmental source, is tolerant to 40 °C and was found to have a cytopathic effect on MDCK cells, suggesting higher pathogenicity (Possamai et al. 2018). The rare genotype T1 has been found few times in nature (Lorenzo-Morales et al. 2005, 2006; Huang and Hsu 2010). It has also been isolated from the oral cavity of an immunocompromised patient (Niyyati et al. 2017). When detected in infections, the T1 genotype has been associated with GAE but not AK (Alsam et al. 2005; Booton et al. 2005; Sissons et al. 2006). The mechanism underlying the ability of T1 strains to invade the brain but not the other organs remains unknown (Booton et al. 2005). Further experiments, such as in vivo assays to induce infection, could determine if the higher protease activity of R2P5 could be associated with significant virulence, especially for GAE, as previously reported for the T1 genotype.
In protozoan parasites, the loss of virulence properties due to prolonged cultivation is a recognised event. Such properties can be restored after passage in host animal models, as observed for Leishmania (Katakura and Kobayashi 1985), Entamoeba histolytica (Kairalia et al. 1978) and the fish parasite Neoparamoeba (Bridle et al. 2015). Concerning Acanthamoeba, studies employing this approach indicated higher virulence of cultures recovered from experimentally infected animals when compared to the corresponding long-term cultures (Veríssimo et al. 2013). Moreover, other properties related to virulence, such as encystment rate and protease activity, tend to augment after in vitro interaction with culture cells, as demonstrated by a study with a T4 genotype strain interacted with HEp-2 cells (Koehsler et al. 2009). In the present study, strains from distinct genotypes maintained under prolonged axenic cultivation were interacted with MDCK cells, similar to that described by Koehsler et al. (2009), to evaluate possible changes in the secreted protease profile and protease quantities. MDCK cell monolayers have been successfully used to study the interaction between amoebae and target cells, which are susceptible to the cytopathic effect of Acanthamoeba trophozoites by either contact-independent or contact-dependent mechanisms (Omaña-Molina et al. 2013; González-Robles et al. 2014). In the present study, Acanthamoeba trophozoites also caused detachment areas in MDCK monolayers, indicating a cytopathic effect (data not shown). As observed in a study by Koehsler et al. (2009), the zymography profiles of all tested strains showed no changes after cell interaction. However, the increase in extracellular protease activity in post-interaction trophozoites observed by Koehsler et al. (2009) was not observed in the current study. In contrast, the strains presented similar activity before and after MDCK interaction (AP4, AR15, LG and AR14 strains) or showed lower protease activity after the interaction (ALX and R2P5 strains). These distinct findings could be related to the type of cell used in the interaction, as HEp-2 passage could stimulate protease production by mechanisms that were not triggered by MDCK cells. Indeed, a previous study indicated that contact with mannose-rich bacteria Corynebacterium xerosis induces the release of protease MIP133 by Acanthamoeba and exacerbates its pathogenic behaviour, while bacteria that express small quantities of mannose do not induce MIP133 production (Alizadeh et al. 2005).
In summary, Acanthamoeba of different genotypes exhibit different serine protease profiles at pH 7.4, which were unchanged after interaction with MDCK cells. The higher secretion of proteases by the environmentally isolated T1 genotype strain highlights its potential to cause infections.
References
Alfieri SC, Correia CE, Motegi SA, Pral EM (2000) Proteinase activities in total extracts and in medium conditioned by Acanthamoeba polyphaga trophozoites. J Parasitol 86:220–227. https://doi.org/10.1645/0022-3395(2000)086[0220:PAITEA]2.0.CO;2
Alizadeh H, Neelam S, Hurt M, Niederkorn JY (2005) Role of contact lens wear, bacterial flora, and mannose-induced pathogenic protease in the pathogenesis of amoebic keratitis. Infect Immun 73:1061–1068. https://doi.org/10.1128/IAI.73.2.1061-1068.2005
Alsam S, Sissons J, Jayasekera S, Khan NA (2005) Extracellular proteases of Acanthamoeba castellanii (encephalitis isolate belonging to T1 genotype) contribute to increased permeability in an in vitro model of the human blood-brain barrier. J Infect 51:150–156. https://doi.org/10.1016/j.jinf.2004.09.001
Alves D d SMM, Moraes AS, Alves LM et al (2016) Experimental infection of T4 Acanthamoeba genotype determines the pathogenic potential. Parasitol Res. 115:3435–3440. https://doi.org/10.1007/s00436-016-5105-3
Booton GC, Visvesvara GS, Byers TJ, Kelly DJ, Fuerst PA (2005) Identification and distribution of Acanthamoeba species genotypes associated with nonkeratitis infections. J Clin Microbiol 43:1689–1693. https://doi.org/10.1128/JCM.43.4.1689-1693.2005
Bridle AR, Davenport DL, Crosbie PBB, Polinski M, Nowak BF (2015) Neoparamoeba perurans loses virulence during clonal culture. Int J Parasitol 45:575–578. https://doi.org/10.1016/J.IJPARA.2015.04.005
Clarke DW, Niederkorn JY (2006) The pathophysiology of Acanthamoeba keratitis. Trends Parasitol 22:175–180. https://doi.org/10.1016/j.pt.2006.02.004
Corsaro D, Walochnik J, Köhsler M, Rott MB (2015) Acanthamoeba misidentification and multiple labels: redefining genotypes T16, T19, and T20 and proposal for Acanthamoeba micheli sp. nov. (genotype T19). Parasitol Res 114:2481–2490. https://doi.org/10.1007/s00436-015-4445-8
Da Rocha-Azevedo B, Costa E, Silva-Filho F (2007) Biological characterization of a clinical and an environmental isolate of Acanthamoeba polyphaga: analysis of relevant parameters to decode pathogenicity. Arch Microbiol 188:441–449. https://doi.org/10.1007/s00203-007-0264-3
De Souza Carvalho FR, Carrijo-Carvalho LC, Chudzinski-Tavassi AM et al (2011) Serine-like proteolytic enzymes correlated with differential pathogenicity in patients with acute Acanthamoeba keratitis. Clin Microbiol Infect 17:603–609. https://doi.org/10.1111/j.1469-0691.2010.03252.x
Duarte JL, Furst C, Klisiowicz DR, Klassen G, Costa AO (2013) Morphological, genotypic, and physiological characterization of Acanthamoeba isolates from keratitis patients and the domestic environment in Vitoria, Espírito Santo, Brazil. Exp Parasitol 135:9–14. https://doi.org/10.1016/j.exppara.2013.05.013
Dudley R, Alsam S, Khan NA (2008) The role of proteases in the differentiation of Acanthamoeba castellanii. FEMS Microbiol Lett 286:9–15. https://doi.org/10.1111/j.1574-6968.2008.01249.x
González-Robles A, Salazar-Villatoro L, Omaña-Molina M et al (2014) Morphological features and in vitro cytopathic effect of Acanthamoeba griffini trophozoites isolated from a clinical case. J Parasitol Res 2014:256310. https://doi.org/10.1155/2014/256310
Huang S-W, Hsu B-M (2010) Isolation and identification of Acanthamoeba from Taiwan spring recreation areas using culture enrichment combined with PCR. Acta Trop 115:282–287. https://doi.org/10.1016/j.actatropica.2010.04.012
Hurt M, Niederkorn J, Alizadeh H (2003) Effects of mannose on Acanthamoeba castellanii proliferation and cytolytic ability to corneal epithelial cells. Investig Ophthalmol Vis Sci 44:3424–3431. https://doi.org/10.1167/iovs.03-0019
Kairalia AB, Lushbaugh WB, Pittman FE, Loadholt CB (1978) Effect of hamster liver passage on the virulence of axenically cultivated Entamoeba histolytica. Am J Trop Med Hyg 27:248–254. https://doi.org/10.4269/ajtmh.1978.27.248
Katakura K, Kobayashi A (1985) Enhancement of infectivity of Leishmania donovani promastigotes by serial mouse passages. J Parasitol 71:393–394
Khan NA (2006) Acanthamoeba : biology and increasing importance in human health. FEMS Microbiol Rev 30:564–595. https://doi.org/10.1111/j.1574-6976.2006.00023.x
Khan NA (2009) Acanthamoeba : biology and pathogenesis. Caister Academic
Khan NA, Jarroll EL, Panjwani N, Cao Z, Paget TA (2000) Proteases as markers for differentiation of pathogenic and nonpathogenic species of Acanthamoeba. J Clin Microbiol 38:2858–2861
Khan NA, Jarroll EL, Paget TA (2002) Molecular and physiological differentiation between pathogenic and nonpathogenic Acanthamoeba. Curr Microbiol 45:197–202. https://doi.org/10.1007/s00284-001-0108-3
Kim WT, Kong HH, Ha YR, Hong YC, Jeong HJ, Yu HS, Chung DI (2006) Comparison of specific activity and cytopathic effects of purified 33 kDa serine proteinase from Acanthamoeba strains with different degree of virulence. Korean J Parasitol 44:321–330. https://doi.org/10.3347/kjp.2006.44.4.321
Koehsler M, Leitsch D, Duchêne M, Nagl M, Walochnik J (2009) Acanthamoeba castellanii : growth on human cell layers reactivates attenuated properties after prolonged axenic culture. FEMS Microbiol Lett 299:121–127. https://doi.org/10.1111/j.1574-6968.2009.01680.x
Lorenzo-Morales J, Ortega-Rivas A, Foronda P, Martínez E, Valladares B (2005) Isolation and identification of pathogenic Acanthamoeba strains in Tenerife, Canary Islands, Spain from water sources. Parasitol Res 95:273–277. https://doi.org/10.1007/s00436-005-1301-2
Lorenzo-Morales J, Ortega-Rivas A, Martinez E et al (2006) Acanthamoeba isolates belonging to T1, T2, T3, T4 and T7 genotypes from environmental freshwater samples in the Nile Delta region, Egypt. Acta Trop 100:63–69. https://doi.org/10.1016/j.actatropica.2006.09.008
Lorenzo-Morales J, Khan NA, Walochnik J (2015) An update on Acanthamoeba keratitis: diagnosis, pathogenesis and treatment. Parasite 22:10. https://doi.org/10.1051/parasite/2015010
Maciver SK, Asif M, Simmen MW, Lorenzo-Morales J (2013) A systematic analysis of Acanthamoeba genotype frequency correlated with source and pathogenicity: T4 is confirmed as a pathogen-rich genotype. Eur J Protistol 49:217–221. https://doi.org/10.1016/j.ejop.2012.11.004
Magliano ACM, da Silva FM, Teixeira MMG, Alfieri SC (2009) Genotyping, physiological features and proteolytic activities of a potentially pathogenic Acanthamoeba sp. isolated from tap water in Brazil. Exp Parasitol 123:231–235. https://doi.org/10.1016/j.exppara.2009.07.006
Mahdavi Poor B, Dalimi A, Ghafarifar F, Khoshzaban F, Abdolalizadeh J (2017) Characterization of extracellular proteases of Acanthamoeba genotype T4 isolated from different sources in Iran. Parasitol Res 116:3373–3380. https://doi.org/10.1007/s00436-017-5656-y
Marciano-Cabral F, Cabral G (2003) Acanthamoeba spp. as agents of disease in humans. Clin Microbiol Rev 16:273–307. https://doi.org/10.1128/CMR.16.2.273-307.2003
Mattana A, Cappai V, Alberti L, Serra C, Fiori PL, Cappuccinelli P (2002) ADP and other metabolites released from Acanthamoeba castellanii lead to human monocytic cell death through apoptosis and stimulate the secretion of proinflammatory cytokines. Infect Immun 70:4424–4432. https://doi.org/10.1128/iai.70.8.4424-4432.2002
Niyyati M, Arab-Mazar Z, Lasjerdi Z, Lorenzo-Morales J, Espotin A, Yadegarynia D, Gachkar L, Rahmati Roodsari S (2017) Molecular characterization of Acanthamoeba strains isolated from the oral cavity of hemodialysis patients in Iran. Parasitol Res 116:2965–2969. https://doi.org/10.1007/s00436-017-5605-9
Nuprasert W, Putaporntip C, Pariyakanok L, Jongwutiwes S (2010) Identification of a novel T17 genotype of Acanthamoeba from environmental isolates and T10 genotype causing keratitis in Thailand. J Clin Microbiol 48:4636–4640. https://doi.org/10.1128/JCM.01090-10
Omaña-Molina M, González-Robles A, Iliana Salazar-Villatoro L, Lorenzo-Morales J, Cristóbal-Ramos AR, Hernández-Ramírez VI, Talamás-Rohana P, Méndez Cruz AR, Martínez-Palomo A (2013) Reevaluating the role of Acanthamoeba proteases in tissue invasion: observation of cytopathogenic mechanisms on MDCK cell monolayers and hamster corneal cells. Biomed Res Int. 2013:461329. https://doi.org/10.1155/2013/461329
Page MA, Mathers WD (2013) Acanthamoeba keratitis: a 12-year experience covering a wide spectrum of presentations, diagnoses, and outcomes. J Ophthalmol 2013:670242. https://doi.org/10.1155/2013/670242
Possamai CO, Loss AC, Costa AO, Falqueto A, Furst C (2018) Acanthamoeba of three morphological groups and distinct genotypes exhibit variable and weakly inter-related physiological properties. Parasitol Res 117:1389–1400. https://doi.org/10.1007/s00436-018-5824-8
Serrano-Luna JDJ, Cervantes-Sandoval I, Calderón J et al (2006) Protease activities of Acanthamoeba polyphaga and Acanthamoeba castellanii. Can J Microbiol 52:16–23. https://doi.org/10.1139/W05-114
Sissons J, Kim KS, Stins M, Jayasekera S, Alsam S, Khan NA (2005) Acanthamoeba castellanii induces host cell death via a phosphatidylinositol 3-kinase-dependent mechanism. Infect Immun 73:2704–2708. https://doi.org/10.1128/IAI.73.5.2704-2708.2005
Sissons J, Alsam S, Goldsworthy G, Lightfoot M, Jarroll EL, Khan NA (2006) Identification and properties of proteases from an Acanthamoeba isolate capable of producing granulomatous encephalitis. BMC Microbiol 6:42. https://doi.org/10.1186/1471-2180-6-42
Stothard DR, Schroeder-Diedrich JM, Awwad MH, Gast RJ, Ledee DR, Rodriguez-Zaragoza S, Dean CL, Fuerst PA, Byers TJ (1998) The evolutionary history of the genus Acanthamoeba and the identification of eight new 18S rRNA gene sequence types. J Eukaryot Microbiol 45:45–54. https://doi.org/10.1111/j.1550-7408.1998.tb05068.x
Szajn H, Csopak H (1977) Metal ion-induced conformational changes in Escherichia coli alkaline phosphatase. Biochim Biophys Acta 480:143–153. https://doi.org/10.1016/0005-2744(77)90329-1
Veríssimo CDM, Maschio VJ, Correa APF et al (2013) Infection in a rat model reactivates attenuated virulence after long-term axenic culture of Acanthamoeba spp. Memórias do Inst Oswaldo Cruz 108:832–835. https://doi.org/10.1590/0074-0276130099
Funding
This study received financial support from Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG APQ-01100-14), Fundação Estadual de Amparo à Pesquisa do Estado do Espírito Santo (FAPES; grant number 68854315/14) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES grant number AUXPE 1526/2011), an entity of the Brazilian government for the qualification of human resources and financial aid for educational or research projects.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Handling Editor: Julia Walochnik
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Cirelli, C., Mesquita, E.I.S., Chagas, I.A.R. et al. Extracellular protease profile of Acanthamoeba after prolonged axenic culture and after interaction with MDCK cells. Parasitol Res 119, 659–666 (2020). https://doi.org/10.1007/s00436-019-06562-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-019-06562-w