
Abstract
Despite the wide distribution of wild boar populations in Italy and the increase of its diffusion in urbanized areas, only one case report has described the occurrence of Echinococcus granulosus s.l. in a wild boar from Marche (Central Italy). The present study investigated the presence of E. granulosus sensu lato with an epidemiological survey on wild boars from central Italy that had been killed during hunting season. Seven hundred sixty-five (765) adult wild boars were examined during the 2016–2017 hunting season. Of these animals, 1.0% (8/765) were positive to E. granulosus s.l. with a fertility of 0.3% (2/765), and 2.9% animals (22/765) were positive for the metacestode stage of Taenia hydatigena (Cysticercus tenuicollis), while 0.5% (4/765) showed mixed infection (E. granulosus s.l. + T. hydatigena). Sixteen hydatids were found, of which 12.5% were fertile, 37.5% were sterile, 31.3% were caseous, and 18.8% were calcified. Eight hydatids (two fertile and six sterile cysts) were molecularly characterized by analysis of the mitochondrial gene, cytochrome c oxidase subunit 1 (cox1), and the NADH dehydrogenase subunit 1 (ND1). Hydatids found in wild boars were characterized as E. granulosus sensu stricto (G1 genotype). The present survey represents the first epidemiological study on cystic echinococcosis in wild boar in Italy which highlights the need for more extensive epidemiological investigations to determine the causal factors, economic impact, and public health importance of the disease in this livestock-wildlife setting.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cystic echinococcosis (CE) is a zoonosis caused by the larval stage of Echinococcus granulosus sensu lato (s.l.) species complex. The disease represents a major public health concern worldwide and is currently considered among the five most common zoonoses in the Mediterranean area (Varcasia et al. 2011). The genetic diversity of E. granulosus s.l. species complex is considerably high and has been divided into genotypes (genotype groups) G1-G10 (Bowles et al. 1992, 1994; Lavikainen et al. 2003). The evidence of various morphological, host range, and life cycle differences between genotypes has led to several of these genotypes being considered as separate species: E. granulosus sensu stricto (s.s.; G1, G2, and G3), E. equinus (G4), E. ortleppi (G5), and E. canadensis (G6–G10) (Thompson and McManus 2002; Lymbery 2017; Yanagida et al. 2017; Kinkar et al. 2018). Recently, it was even suggested that G6/G7 and G8/G10 could be considered two distinct species (Laurimäe et al. 2018).
Wild boars are intermediate hosts of E. granulosus s.l. and are likely to be involved in the epidemiology of E. granulosus in its sylvatic cycle (Di Paolo et al. 2017). To date, E. granulosus sensu stricto (s.s.; G1, G2, and G3) and E. canadensis (G6-G10) have been recorded in wild boars in various parts of the world (Martín-Hernando et al. 2008; Senlik et al. 2011; Onac et al. 2013; Umhang et al. 2014; Kinkar et al. 2016). These two species have different life cycles as well as different genetic and biochemical characteristics (Eckert and Deplazes 2004).
Despite the wide distribution of wild boar populations in Italy (Toso and Pedrotti 2001) and the increasing spread of this wild ungulate in urbanized areas (Di Nicola et al. 2015), only one case report has been described on the occurrence of E. granulosus s.l. in a wild boar from Marche (Central Italy) (Di Paolo et al. 2017). The present study evaluated the presence of E. granulosus s.l. in wild boars from the Central Italy with an epidemiological survey on animals killed during the hunting season.
Materials and methods
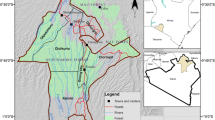
The study was carried out in the National Park of the Sibillini Mountains and Gran Sasso Mountains of Laga National Park, in Marche, Italy (42.8554018° N, 13.5749771° E). Seven hundred and sixty-five (765) adult wild boars were examined during the hunting season in 2016–2017 at the slaughterhouse (A.S.U.R. Ascoli Piceno, Italy). At necropsy, the viscera of animal were carefully examined for the presence and location of hydatids by visual inspection, palpation, and systematic dissection. All infected organs were removed and transported in individual bags as soon as possible to the Faculty of Veterinary Medicine of Teramo (Italy).
For each organ, the number and the location of the cysts were recorded. Hydatid cysts, when found, were ranked as fertile, sterile, or calcified/caseous, as described elsewhere (Varcasia et al. 2006). Hydatid fertility was assessed after dissection and collection of the germinal layer to verify the presence and vitality of protoscoleces through flame cell activity and motility by light microscopy without staining (Scala et al. 2006).
DNA was then extracted from the germinal layer and protoscoleces from single fertile and sterile cysts, using a genomic DNA extraction kit (QIAamp DNA Mini Kit, QIAGEN GmbH, Germany) according to manufacturer’s instructions. About 10–20 ng of template DNA was used to amplify a fragment within the cytochrome c oxidase subunit 1 (cox1) and the NADH dehydrogenase subunit 1 (ND1) mitochondrial genes following protocols published by Bowles et al. (1992) and Bowles and McManus (1993). PCR products were commercially sequenced by MWG-Biotech (Ebersberg, Germany). The nucleotide sequences generated were edited in Proseq 3.5 (Filatov 2002) and compared with those available in the National Center for Biotechnology Information (NCBI) database using the basic local alignment search tool (BLAST).
Results
The survey showed an overall prevalence of 3.9% for metacestodes in the hunted animals examined. Of these animals, 1.0% (8/765) were positive to E. granulosus s.l., with a fertility of 0.3% (2/765 examined; 2/8 positive to CE). The survey identified 2.9% animals (22/765) positive to larval forms of Taenia hydatigena (Cysticercus tenuicollis), while 0.5% (4/765) showed mixed infection (E. granulosus s.l. + T. hydatigena).
A total of 16 hydatids were found, of which 12.5% were fertile, 37.5% were sterile, 31.3% were caseous, and 18.8% were calcified. Regarding cyst location, 56.3% of the hydatids, including all the fertile cysts, were found in the lungs, and 43.8% in the livers of positive animals. The viability of fertile cysts was 0.3% (2/765) in examined animals and 25% in positive animals. Fifty-two C. tenuicollis were found in the examined animals, of which 71.7% were in the liver, 15.2% were in the lungs, 10.9% in the omentum, and 2.2% in the thoracic diaphragm. All cysticerci found were viable with their characteristic bottleneck morphology. Eight hydatids (two fertile and six sterile cysts) were molecularly characterized as E. granulosus sensu stricto, showing 100% identity with GenBank accession numbers KT2002231.1 and KY76689.1 from E. granulosus sensu stricto (former genotype G1).
Discussion and conclusions
The present survey represents the first epidemiological study on cystic echinococcosis in wild boar in Italy after the first case report of the parasite by Di Paolo et al. (2017). Authors believe that a full sylvatic lifecycle of E. granulosus in the region could be improbable even wolves are present and widespread in the same region (Scotti et al. 2012). Anyway, the unlikely, though possible predation of wolves on adult wild boars and low prevalence found for CE in wild boars most likely suggest that wolves and wild boar could interact in a semi-domestic lifecycle, in which wild ungulates can be infected by wolves and shepard and stray dog fecal contamination wild carnivores, could be infected by preying on sheep and other domestic animals in the nearby farms where cystic echinococcosis has been already described (Garippa and Manfredi 2009). The distance between the area where positive wild boars have been found and the nearest positive sheep farms (maximum 5-km radium) is consistent with the daily movement capacities of wild boar (from 1 to 16 km) (Lemel et al. 2003). Most likely, the environmental contamination due to common home slaughtering of sheep, incorrect disposal of offal, and uncontrolled dog movements may contribute to the transmission of these parasites to wild boars (Varcasia et al. 2011). This speculation is consistent with the finding of E. granulosus sensu stricto in the present survey which is endemic in Italy and in the islands, with a typical sheep-dog lifecycle (Dore et al. 2014).
Previous surveys on CE in wild boar have reported the G1 and G7 strains; in particular Kedra et al. (2000) reported for the first time the G7 in a wild boar from the Sumy region, Ukraine, while Daniel Mwambete et al. (2004) reported the G7 in wild boar in Spain from fertile cysts and G1 in sterile cysts. Some years later, Martin-Hernando et al. (2008) reported a massive infection in a wild boar from Spain caused by G1 strain of E. granulosus. Similar observations were also reported in a study of wild boar in south western Iran, where the hydatids found were fertile, and molecular investigations confirmed the presence of the sheep strain, G1 genotype (Sarkari et al. 2015).
Onac et al. (2013) reported a prevalence of 12.3% for hydatid cysts in wild boar in Romania, with a mixed infection by G1 and G7 in 45.5% and 39.4% of cases, respectively.
In a recent survey Umhang et al. (2014) recorded the G6/7 genotype of E. canadensis in both pigs and wild boar from Corsica.
Fertility rates appear to be limited (0.3%) compared with the rates found in domestic pigs from organic farms (e.g., 7.6%) (Varcasia et al. 2007). The percentage of viable hydatids in positive animals (25%) was high, considering that pigs and wild boar are not primary target host of E. granulosus sensu stricto. Anyway, the fertility rates found in wild boar were similar to the prevalence of E. granulosus sensu stricto found in other hosts, such as cattle (Poglayen et al. 2017).
Larval stages of T. hydatigena (Cysticercus tenuicollis) have been found in a wide range of ruminants and also pigs (Troncy 1989) and in several countries (De-la-Muela et al. 2001; Senlik et al. 2011; Boufana et al. 2015; Mansouri et al. 2016). The importance of T. hydatigena metacestodosis is due to the economic damage caused by larvae migrations due to massive contamination, especially in young ruminants (Scala et al. 2015, 2016); however, little information is available on wild boar. In the present study, T. hydatigena metacestodes were found with a higher prevalence compared with cystic echinococcosis.
Our findings shown that wild boars examined in the present work roam in areas where domestic and wild carnivores (shepard dogs and wolves) have access to raw offal of infected sheep and/or other animals. T. hydatigena and E. granulosus s.l. have a different biotic potential (i.e., number of proglottids and eggs released) (Torgerson et al. 1998; Gemmell et al. 1987) but the same lifecycle. This suggests that wild boars positive to cysticercosis living in contaminated areas could have a greater risk of exposure to CE compared with other animals, also bearing in mind that CE grows slowly and thus may not be clearly visible especially in young animals (Scala et al. 2016; Varcasia et al. 2004). Moreover, the expansion of wild boars towards urban areas and agricultural lands (Di Nicola et al. 2015) could facilitate the interaction between wild boars, domestic animals, definitive hosts, and their parasites.
More surveys are thus necessary to better define the role of wild hosts in the transmission and epidemiology of cystic echinococcosis and other metacestodosis, as well as to verify the existence of alternative wild lifecycles of these parasites in Italy.
References
Boufana B, Scala A, Lahmar S, Pointing S, Craig PS, Dessì G, Zidda A, Pipia AP, Varcasia A (2015) A preliminary investigation into the genetic variation and population structure of Taenia hydatigena from Sardinia, Italy. Vet Parasitol 214:67–74
Bowles J, McManus DP (1993) NADH dehydrogenase 1 gene sequences compared for species and strains of the genus Echinococcus. Int J Parasitol 23:969–972
Bowles J, Blair D, McManus DP (1992) Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol Biochem Parasitol 54:165–173
Bowles J, Blair D, McManus DP (1994) Molecular genetic characterization of the cervid strain (‘northern form’) of Echinococcus granulosus. Parasitology 109:215–221
Daniel Mwambete K, Ponce-Gordo F, Cuesta-Bandera C (2004) Genetic identification and host range of the Spanish strains of Echinococcus granulosus. Acta Trop 91:87–93
De-la-Muela N, Hernandez-de-Lujan S, Ferre I (2001) Helminths of wild boar in Spain. J Wild Dis 37:840–843
Di Nicola U, Scacchia M, Marruchella G (2015) Pathological and serological findings in wild boars (Sus scrofa) from Gran Sasso and Monti della Laga National Park (Central Italy). Large Anim Rev 21:167–171
Di Paolo A, Piseddu T, Sebastianelli M, Manuali E, Corneli S, Paniccià M, Papa P, Viali S, Mazzone P (2017) Detection of Echinococcus granulosus G3 in a wild boar (Sus scrofa) in Central Italy using PCR and sequencing. J Wildl Dis 53:399–401
Dore F, Varcasia A, Pipia AP, Sanna G, Pinna Parpaglia ML, Corda A, Romig T, Scala A (2014) Ultrasound as a monitoring tool for cystic echinococcosis in sheep. Vet Parasitol 203:59–64
Eckert J, Deplazes P (2004) Biological, epidemiological, and clinical aspects of Echinococcosis, a zoonosis of increasing concern. Clin Microbiol Rev 17:107–135
Filatov DA (2002) PROSEQ: a software for preparation and evolutionary analysis of DNA sequence data sets. Mol Ecol Notes 2:621–624
Garippa G, Manfredi MT (2009) Cystic Echinococcosis in Europe and in Italy. Vet Res Commun 33:35–39
Gemmell MA, Lawson JR, Roberts M (1987) Population dynamics in echinococcosis and cysticercosis: evaluation of the biological parameters of Taenia hydatigena and T. ovis and comparison with those of Echinococcus granulosus. Parasitology 94(1):161–180
Kedra AH, Tkach VV, Swiderski ZP, Pawlowski Z, Emets A, Pawlowski J (2000) Molecular characterisation of Echinococcus granulosus from a wild boar. Acta Parasitol 45(2):121–122
Kinkar L, Laurimaë T, Simsek S, Balkaya I, Casulli A, Manfredi MT, Ponce-Gordo F, Varcasia A, Lavikainen A, González LM, Rehbein S, Van Der Giessen J, Sprong H, Saarma U (2016) High-resolution phylogeography of zoonotic tapeworm Echinococcus granulosus sensu stricto genotype G1 with an emphasis on its distribution in Turkey, Italy and Spain. Parasitology 143(13):1790–1801
Kinkar L, Laurimäe T, Balkaya I, Casulli A, Zait H, Irshadullah M, Sharbatkhori M, Mirhendi H, Rostami-Nejad M, Ponce-Gordo F, Rehbein S, Kia EB, Simsek S, Šnábel V, Umhang G, Varcasia A, Saarma U (2018) Genetic diversity and phylogeography of the elusive, but epidemiologically important Echinococcus granulosus sensu stricto genotype G3. Parasitology:1–10 Article in Press
Laurimäe T, Kinkar L, Moks E, Romig T, Omer RA, Casulli A, Umhang G, Bagrade G, Irshadullah M, Sharbatkhori M, Mirhendi H, Ponce-Gordo F, Soriano SV, Varcasia A, Rostami-Nejad M, Andresiuk V, Saarma U (2018) Molecular phylogeny based on six nuclear genes suggests that Echinococcus granulosus sensu lato genotypes G6/G7 and G8/G10 can be regarded as two distinct species. Parasitology, in press:1–9
Lavikainen A, Lehtinen MJ, Meri T, Hirvelä-Koski V, Meri S (2003) Molecular genetic characterization of the Fennoscandian cervid strain, a new genotypic group (G10) of Echinococcus granulosus. Parasitology 127:207–215
Lemel J, Truvé J, Söderberg B (2003) Variation in ranging and activity behaviour of European wild boar Sus scrofa in Sweden. Wildlife Biol 9:29–36
Lymbery AJ (2017) Phylogenetic pattern, evolutionary processes and species delimitation in the genus Echinococcus. Adv Parasitol 95:111–145
Mansouri M, Sarkari B, Mowlavi GR (2016) Helminth parasites of wild boars, Sus scrofa, in Bushehr Province, Southwestern Iran. Iran J Parasitol 11:377–382
Martín-Hernando MP, González LM, Ruiz-Fons F, Garate T, Gortazar C (2008) Massive presence of Echinococcus granulosus (Cestoda, Taeniidae) cysts in a wild boar (Sus scrofa) from Spain. Parasitol Res 103:705–707
Onac D, Győrke A, Oltean M, Gavrea R, Cozma V (2013) First detection of Echinococcus granulosus G1 and G7 in wild boars (Sus scrofa) and red deer (Cervus elaphus) in Romania using PCR and PCR-RFLP techniques. Vet Parasitol 193:289–291
Poglayen G, Varcasia A, Pipia AP, Tamponi C, Parigi M, Marchesi B, Morandi B, Benfenati V, Scala A (2017) Retrospective study on Cystic Echinococcosis in cattle of Italy. J Infect Dev Ctries 11:11719–11726. https://doi.org/10.3855/jidc.9433
Sarkari B, Mansouri M, Khabisi SA, Mowlavi G (2015) Molecular characterization and seroprevalence of Echinococcus granulosus in wild boars (Sus scrofa) in South-Western Iran. Ann Parasitol 61:269–273
Scala A, Garippa G, Varcasia A, Tranquillo VM, Genchi C (2006) Cystic Echinococcosis in slaughtered sheep in Sardinia (Italy). Vet Parasitol 135:33–38
Scala A, Pipia AP, Dore F, Sanna G, Tamponi C, Marrosu R, Bandino E, Carmona C, Boufana B, Varcasia A (2015) Epidemiological updates and economic losses due to Taenia hydatigena in sheep from Sardinia, Italy. Parasitol Res 114:3137–3143
Scala A, Urrai G, Varcasia A, Nicolussi P, Mulas M, Goddi L, Pipia AP, Sanna G, Genchi M, Bandino E (2016) Acute visceral cysticercosis by Taenia hydatigena in lambs and treatment with praziquantel. J Helminthol 90:113–116
Scotti M, Burattini R, Dell’Orso M, Di Massimo S, Forconi P, Gazzola A, Giacchini P, Marini G, Marini S, Piccinetti F, Randi E, Riganelli N, Salvi P, Soriani G, Vedova A, Zabaglia C (2012) Indagine conoscitiva sulla presenza del lupo Canis lupus nella regione Marche. Hystrix It J Mamm 181 (In Italian)
Senlik B, Cirak VY, Girisgin O, Akyol CV (2011) Helminth infections of wild boars (Sus scrofa) in the Bursa province of Turkey. J Helminthol 85:404–408
Thompson RC, McManus DP (2002) Towards a taxonomic revision of the genus Echinococcus. Trends Parasitol 18:452–457
Torgerson PR, Williams DH, Abo-Shehada MN (1998) Modelling the prevalence of Echinococcus and Taenia species in small ruminants of different ages in northern Jordan. Vet Parasitol 79(1):35–51
Toso S, Pedrotti L (2001) Linee guida per la gestione del cinghiale (Sus scrofa) nelle aree protette Quad. Cons.Natura, 2, Min. Ambiente – Ist. Naz. Fauna Selvatica (In Italian)
Troncy PM (1989) Helminth parasites of livestock and poultry in tropical Africa. In: Manual of Tropical Veterinary Parasitology, Translated by M. Shah-Fischer and R. R. Say, C.A.B. International, Wallingford, UK 7–164
Umhang G, Richomme C, Hormaz V, Boucher JM, Boué F (2014) Pigs and wild boar in Corsica harbor Echinococcus canadensis G6/7 at levels of concern for public health and local economy. Acta Trop 133:64–68
Varcasia A, Garippa G, Scala A (2004) The diagnosis of Echinococcus granulosus in dogs. Parassitologia 46:409–412
Varcasia A, Canu S, Lightowlers MW, Scala A, Garippa G (2006) Molecular characterization of Echinococcus granulosus strains in Sardinia. Parasitol Res 98:273–277
Varcasia A,. Tosciri G, Pedes T, Pipia AP, Marrosu R, Scala A, Garippa G (2007). Cystic Echinococcosis in pigs and wild boars of Sardinia (Italy), Proceedings of the 6th International Symposium on the Mediterranean Pig - Capo d’Orlando (ME), Italy, ISSN 2035-4088
Varcasia A, Tanda B, Giobbe M, Solinas C, Pipia AP, Malgor R, Carmona C, Garippa G, Scala A (2011) Cystic Echinococcosis in Sardinia: farmers’ knowledge and dog infection in sheep farms. Vet Parasitol 181:335–340
Yanagida T, Lavikainen A, Hoberg EP, Konyaev S, Ito A, Sato MO, Zaikov VA, Beckmen K, Nakao M (2017) Specific status of Echinococcus canadensis (Cestoda: Taeniidae) inferred from nuclear and mitochondrial gene sequences. Int J Parasitol 47:971–979
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical statement
Animals included in the present study were examined during post mortem mandatory inspection visit by official veterinaries physicians of the Zooprofilattico Sperimentale of Umbria and the Marche Institute, and ASUR Sanitary Units (Ministry of Health, Italy) and according to a specific agreement between Parco Nazionale of the Sibillini Mountains for sanitary monitoring of hunted animals and prevention of zoonosis (Parco Nazionale Monti Sibillini: Prot. N° 3516 – Cl. 13.4.2; Sanitary Units of Ministry of Health N° 0074901 of 13/10/2017).
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Section Editor: Bruno Gottstein
Rights and permissions
About this article

Cite this article
Paoletti, B., Della Salda, L., Di Cesare, A. et al. Epidemiological survey on cystic echinococcosis in wild boar from Central Italy. Parasitol Res 118, 43–46 (2019). https://doi.org/10.1007/s00436-018-6112-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-018-6112-3