
Abstract
Opisthorchis viverrini sensu lato is a food-borne trematode which is classified as a class 1 carcinogen, with infection potentially leading to cholangiocarcinoma. Snails of the genus Bithynia act as the first intermediate hosts and an amplifying point in the parasite life cycle. In order to investigate seasonal effect on transmission dynamics of O. viverrini in Bithynia siamensis goniomphalos, cercarial emergence and output profiles were monitored at different season. A total of 4533 snails originating from Sakon Nakhon Province, Thailand, collected during the three main seasons, were analyzed for O. viverrini s.l. infection. Emergence of O. viverrini s.l. cercariae from snails was monitored daily from 06:00 to 18:00 h for seven consecutive days. The prevalence of infection in the snails was highest in the hot-dry season and declined in the rainy and cool-dry seasons. Peak cercarial emergence occurred between 08:00 and 10:00 h during the rainy and cool-dry seasons and between 10:00 and 12:00 h during the hot-dry season. The cercarial output was highest in the hot-dry season, similar to a previous study from Lao People’s Democratic Republic (PDR). Average cercarial output/snail in Thailand was higher than in Lao PDR. The number of cercariae emerging from the snails was strongly related to snail size, but the relationship between prevalence of infection and snail size differed between seasons. Observed discrepancies in the emergence patterns and per capita cercarial release may reflect differences in environmental, snail, and/or parasite factors particularly biological characteristics between the cryptic species of O. viverrini s.l. and B. s. goniomphalos from Thailand and Lao PDR.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The carcinogenic liver fluke Opisthorchis viverrini sensu lato is a major medical problem endemic in Southeast Asian countries bordering the Mekong River, including Thailand and the Lao People’s Democratic Republic (Lao PDR) (Andrews et al. 2008; Sithithaworn et al. 2012). This fluke is classified as a group 1 carcinogen and is the major causative agent for cholangiocarcinoma (CCA) or bile duct cancer in the area (IARC 2011). The life cycle involves freshwater snails as first intermediate hosts, cyprinid fish as second intermediate hosts, humans as the definitive hosts and cats, and, to a lesser extent, dogs as reservoir hosts (Sithithaworn et al. 2012). Infection in humans is caused by eating raw or undercooked fish containing with metacercariae of O. viverrini. Fish hosts usually have a very high prevalence of infection (Saijuntha et al. 2014), while in the snail hosts, very low infection rates occur (Kiatsopit et al. 2012; Petney et al. 2012).
After the eggs hatch in the snail, the parasite multiplies asexually and produces relatively large numbers of cercariae. The biological characteristics of the cercariae, together with the environmental conditions, play a major role in trematode transmission (Anderson and May 1991; Haas 1994; Poulin 2006). Bithynia snails are the critical amplifying hosts of O. viverrini s.l., and they play a major role in transmission. The free-swimming cercarial stage invades cyprinid fish as the next intermediate host in the life cycle. The natural infection of O. viverrini s.l. surveyed in these snails ranges from 0.22 to 6.93 % (mean of 3.03 %) in Thailand and 0.37 to 8.37 % (mean of 2.01 %) in the Lao PDR (Kiatsopit et al. 2012).
Evidence has shown that O. viverrini s.l. is a species complex with distinct genetic groups in different wetlands (Saijuntha et al. 2007). Furthermore, Saijuntha et al. (2007) provided evidence that different genetic groups of Bithynia siamensis goniomphalos are associated with the different O. viverrini s.l. genetic groups within specific wetlands. Snails could be divided into two distinct genetic groups that corresponded with the delineation of O. viverrini s.l. into two cryptic species, one in Thailand and the other in Lao PDR. Subsequent results showed that snails from Sakon Nakhon and Nakhon Phanom provinces in the Songkram River wetland in northeast Thailand form a genetic cluster with fixed genetic differences ranging between 52 and 81 % from snails from other wetlands in Thailand, while they cluster more closely with snails from the Nam Ngum River wetland, Lao PDR, although 29–38 % fixed differences are still present (Kiatsopit et al. 2013). These high levels of fixed genetic differences suggest that a third cryptic species of snails exists within the Songkram River wetland from Sakon Nakhon, which is a tributary to the Mekong River. More recently, B. s. goniomphalos has been found to be a species complex containing potentially nine cryptic species associated with defined wetlands that also correspond to the distribution of O. viverrini s.l. cryptic species (Kiatsopit et al. 2013).
Additional evidence from an animal model supports the hypothesis that a morphologically, biologically, and genetically distinct species of O. viverrini s.l. exists within the Songkram River wetland, as well as the genetically distinct species of O. viverrini s.l. in Lao PDR (Laoprom et al. 2009). Although these differences in both O. viverrini s.l. and Bithynia snails have been found between different wetland systems in Thailand and Lao PDR using both molecular and biological characteristics, very little is known about the dynamics of O. viverrini s.l. cercarial release in B. s. goniomphalos.
Two recent studies on O. viverrini s.l. cercariae in snails, one from Lao PDR (Kiatsopit et al. 2014) and one from the Phang Khon district of Sakon Nakhon Province in Thailand (Namsanor et al. 2015), revealed that the infection characteristics of infected snails varied seasonally, with the highest prevalences occurring in the cool-dry season. Peak cercarial emergence occurred between 08:00 and 10:00 h during the hot-dry season and between 12:00 and 14:00 h during the rainy and cool-dry seasons (Kiatsopit et al. 2014). The cercarial output was highest in the hot-dry season. The prevalence of infection and the output of snails releasing cercariae were strongly dependent on snail size (Kiatsopit et al. 2014). For comparative purposes, we examined the patterns of cercarial emergence and output of cercariae released from B. s. goniomphalos originating from the Songkram River wetland in Thailand at different seasons.
Materials and methods
Sample collection
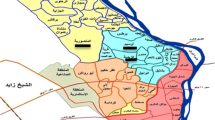
A total of 4533 B. s. goniomphalos were collected from Sakon Nakhon Province, Thailand (georeference coordinates, 17.348163, 103.570602; Fig. 1) on three sampling dates in 2012: the first collection was made in April (hot-dry season, average temperature = 28.21 °C; average rainfall = 4.81 mm/day, sunlight = 7.68 h/day, n = 303), the second in October (rainy season, average temperature = 27.78 °C; average rainfall = 6.54 mm/day, sunlight = 5.64 h/day, n = 1623), and the third from November (cool-dry season, average temperature = 25.04 °C; average rainfall = 0.48 mm/day, sunlight = 8.50 h/day, n = 2607). Snails were collected in paddy fields by handpicking and dredging the sediment with a scoop over an area of approximately 1500 m2 for 2 h. The snails were then cleaned, dried, placed into plastic bags, and transported to the laboratory at Khon Kaen University where they were identified according to standard morphological criteria of Brandt (1974), Chitramvong (1992), and Upatham et al. (1983). The shell size (length and width) of each infected snail was determined using digital Vernier calipers. Shell lengths were measured from the tip of the apex to the lower edge of the aperture. The width was measured in a straight line through the widest part of the shell, from the side of the body whorl to the outermost side of the aperture.

Map of sampling location of Bithynia siamensis goniomphalos from Sakon Nakhon Province, Thailand
Screening of O. viverrini s.l. infection in B. s. goniomphalos
Before the experiments, all snail samples were kept under laboratory conditions for 1 day. To determine the O. viverrini s.l. infections, collected snails were examined by the cercarial shedding method within 2 days to avoid the effects of laboratory maintenance on cercarial emergence. Individual snails were placed separately into a small plastic container (3 cm in diameter, 2.5 cm in height) filled with 5 ml dechlorinated tap water. The containers were covered with a lid studded with pins to prevent the snail from leaving. The snails were then exposed to 1200 lx artificial light placed 30 cm above each container for 5 h during the day at room temperature (25 ± 2 °C) to induce shedding. O. viverrini s.l. cercariae were identified morphologically using light microscopy (Kaewkes 2003). To confirm this morphological identification, PCR analyses were conducted using an O. viverrini s.l. specific primer (Wongratanacheewin et al. 2001). Moreover, O. viverrini s.l. cercariae and other pleurolophocercous cercariae were confirmed by sequencing data using internal transcribed spacer 2 (ITS2) and 28S ribosomal RNA (28s rRNA) primers (Kiatsopit et al., unpublished data).
Emergence of O. viverrini s.l. cercariae
O. viverrini-positive snails were maintained for 1 day in the dark until the experiment started. Snails were placed individually into a plastic container (3 cm in diameter, 2.5 cm height) filled with 5 ml dechlorinated tap water and exposed to light (1200 lx of artificial light placed 30 cm above each container) from 06:00 to 18:00 h at room temperature (25 ± 2 °C). Every 2 h, the snails were removed to a new container and the cercariae released from each snail over the 2-h interval were counted using a dissecting microscope after staining with 1 % Lugol’s iodine solution. At the end of the 12-h period, the snails were returned to the dark for 12 h and fed with leaves of the ivy gourd, Coccinia grandis. They were then exposed to light for 12 h the next day, and this pattern continued for seven consecutive days.
Statistical test
Raw data were entered into a Microsoft Excel spreadsheet, and descriptive statistics were calculated using SPSS software v. 19.0 (IBM Software Company, USA). Statistical comparisons of cercarial release between different times calculated daily for 7 days and seasons were done using the two-related-sample-test followed by Wilcoxon signed ranks tests. Chi-square tests were used to assess the association of size factors with the prevalence of cercariae. Statistical comparisons of the daily cercarial emergence were done using the Mann–Whitney U test. The results were considered significant when P was <0.05.
Results
The prevalence of O. viverrini s.l. cercarial infections in snails was 6.93 % in the hot-dry, 2.65 % in the rainy, and 2.42 % in the cool-dry seasons. Based on morphological characteristics, nine additional types of cercariae were found in snails from Sakon Nakhon, i.e., xiphidiocercariae 1, xiphidiocercariae 2, xiphidiocercariae 3, monostome, mutabile, cystophorous, parapleurolophocercous, furcocercous, and longifurcate-pharyngeate (see in Kiatsopit et al. 2015). A peak in cercarial emergence was observed for the 08.00–10.00-h period in the rainy (mean number of cercariae (±SD) = 889 ± 314 cercariae/snail/2 h/day) and cool-dry (mean number of cercariae = 515 ± 261 cercariae/snail/2 h/day) seasons, whereas in the hot-dry season, this was at 10:00–12:00 h with the average number of cercariae = 1307 ± 889 cercariae/snail/2 h/day (Fig. 2a). In addition, the snails collected in all seasons showed an increase in shedding in the morning followed by a decline until evening (Fig. 2a). The cercarial emergence rates of O. viverrini calculated every 2 h from 06:00 until 18:00 h were significantly different between different times and seasons (P < 0.05). The mean number of cercariae per snail per day was 789 ± 248 cercariae/snail/day for 15 snails in the hot-dry season, 487 ± 51 cercariae/snail/day for 40 snails in the rainy season, and 284 ± 87 cercariae/snail/day for 63 snails in the cool-dry season. The daily cercarial count in the hot-dry season was low until the second day after emergence, increased over the next 2 days, decreased on the fifth, and increased again on the sixth, while on the seventh, it decreased again. In the rainy season, the number decreased until the third day, increased on the fourth, while on the fifth and sixth, it decreased again and increased again on last day. In the cool-dry season, the daily cercarial count was low until the second day after emergence and increased during the subsequent 3 days, while on the sixth, it decreased and the decreased on the seventh day (Fig. 2b). Number of cercariae emerging calculated daily for 7 days was significantly different between different days and seasons (P < 0.05).

Cercarial emergence profiles of Opisthorchis viverrini infecting Bithynia siamensis goniomphalos. Cercarial emergence (a) calculated every 2 h from 06:00 until 18:00 h for 7 days (mean ± SE) and (b) calculated daily for 7 days (mean ± SE) from snails collected in Sakon Nakhon Province, Thailand. (n = no. of positive snails)
The investigations on the relationship between the prevalence of cercariae and the shell size of B. s. goniomphalos show that high prevalences occur in small snails from Sakon Nakhon collected in both rainy and cool-dry seasons (Fig. 3a). In the rainy season, significant differences in the prevalence of cercariae and shell size were found between small and medium snails (χ 2 2 = 12.435, P < 0.001), between small and large snails (χ 2 2 = 24.92, P < 0.001), and between medium and large snails (χ 2 2 = 7.39, P < 0.01) for length. For width, significant differences in cercarial prevalences were found between small and large snails (χ 2 2 = 29.19, P < 0.001) and between medium and large snails (χ 2 2 = 16.86, P < 0.001). In the cool-dry season, small snails had a significantly higher cercarial prevalence than large snails (χ 2 2 = 6.037, P < 0.05) and between medium and large snails (χ 2 2 = 4.34, P < 0.05) for length. For width, a significantly higher cercarial prevalence was found between small and large snails (χ 2 2 = 12.56, P < 0.001) and between medium and large snails (χ 2 2 = 10.79, P < 0.01). In contrast, the relationship between the prevalence of cercariae and shell size of B. s. goniomphalos in hot-dry season shows that high prevalence occurs in large snails with respect to both length and width. For width, significant differences in prevalence of cercariae and shell size were found between small and large snails (χ 2 2 = 10.84, P < 0.001) and between medium and large snails (χ 2 2 = 21.08, P < 0.001). Although the number of cercariae output was higher in larger snails (Fig. 3b), this was not significant (Mann–Whitney U test, P > 0.05).

Relationship between the prevalence (a) and output (b) of Opisthorchis viverrini cercariae and shell size of Bithynia siamensis goniomphalos. Mean values (±95 % CI) are shown. Black bars represent size class based on shell length (small = <8.0 mm, medium = 8.1–10.0 mm, large = >10.0 mm), and white bars represent size classes based on shell width (small = <5.0 mm, medium = 5.1–6.0 mm, large = >6.0). *P < 0.05, **P < 0.01, ***P < 0.001. (n = no. of positive snails)
Discussion
O. viverrini s.l. is a typical trematode undergoing asexual reproduction within snail hosts, releasing large numbers of cercariae into the water bodies that subsequently infect cyprinid fish the second intermediate hosts. The production of cercariae from their snail host is a fundamental component of the transmission success in trematodes (Poulin 2006). The infection rate of snails and the rate of cercarial development show variation related to the geographic origin of the parasite (Boray 1978).
The number of O. viverrini s.l. cercariae was highest in the hot-dry season. Interestingly, the average number of cercariae produced per snail in Thailand was higher than that per snail from Lao PDR in all seasons. The average number of cercariae in snails from Thailand in the hot-dry season (789 ± 248 cercariae/snail/2 h/day) was 3 times higher than from Lao PDR (263 ± 55 cercariae/snail/2 h/day), 5 times in the rainy season (487 ± 51 cercariae/snail/2 h/day from Thailand and 98 ± 76 cercariae/snail/2 h/day from Lao PDR), and 3.1 times in the cool-dry season (284 ± 86 cercariae/snail/2 h/day from Thailand and 91 ± 68 cercariae/snail/2 h/day from Lao PDR). The observed differences may reflect differences between cryptic species of B. s. goniomphalos and/or O. viverrini s.l. (Kiatsopit et al. 2013; Saijuntha et al. 2007). Indeed, a recent molecular genetics study showed substantial differences between the Lao PDR population and that from Sakon Nakhon (Pitaksakulrat et al., unpublished data).
B. s. goniomphalos is a natural intermediate host for O. viverrini in Thailand, which may reflect a greater adaptation to snail hosts in Thailand at each amplification stage following ingestion of eggs, namely, from miracidia to sporocysts to rediae and finally to cercariae. Similarly, Dar et al. (2003) show that the mean number of Fasciola gigantica cercariae shed from Galba truncatula in the Egyptian group was significantly greater than that shed from the Chinese group (a mean of 275.5 per cercaria-shedding per snail compared with 29.0).
Until recently, O. viverrini was considered to be a single species; however, molecular genetic analyses have now shown that it is a species complex containing at least two cryptic species one in Thailand and the other in the Lao PDR (Saijuntha et al. 2007). Furthermore, O. viverrini populations can be divided into six distinct genetic groups that are associated with five different river wetland systems. O. viverrini from the Songkram River wetland in Thailand clustered with O. viverrini from the Num Ngum River wetland, Lao PDR, while both are separated from other wetlands in Thailand. Similarly, molecular data on B. s. goniomphalos snails from the Songkram River wetland (Nakhon Phanom and Sakon Nakhon) show that they also cluster with those from the Num Ngum River wetland, Lao PDR (Saijuntha et al. 2007). A more recent, comprehensive genetic analysis has demonstrated that B. s. goniomphalos is a species complex containing at least nine cryptic species that have specific associations with defined wetlands in Thailand and Lao PDR (Kiatsopit et al. 2013). Importantly, Kiatsopit et al. (2013) also found that cryptic species of B. s. goniomphalos were associated with cryptic species and/or genetic groups of O. viverrini from the same wetlands in Thailand and Lao PDR. A study of O. viverrini from Songkram River, Sakon Nakhon, Thailand showed separation from all other isolates from Thailand and Lao PDR. This suggests that O. viverrini sensu lato from Sakon Nakhon may be another cryptic species within Thailand (Pitaksakulrat et al., unpublished data).
Analysis of the biological characteristics of O. viverrini from localities in Thailand and Lao PDR has provided substantial evidence that O. viverrini from Nakhon Phanom and Sakon Nakhon, Songkram River wetland, northeast Thailand represents a biologically different species based on body size, fecundity, and infectivity compared to O. viverrini from other localities in Thailand and Lao PDR (Laoprom et al. 2009). The separation of O. viverrini into possibly three sibling species is supported by evidence from other molecular and biological data. Fecundity in the Songkram River wetland is lower than in other wetland systems both in Thailand and Lao PDR (Laoprom et al. 2009). A comparison of the prevalence of O. viverrini infection in B. s. goniomphalos showed that there was a higher prevalence in the Songkram River wetland than in the Num Ngum River wetland (Kiatsopit et al. 2012). Fecundity in definitive and animal reservoir hosts may influence the possibility of snail infection; however, host–parasite interactions also have a great impact on the infection outcome.
Previous evidence also shows that peak emergence varies according to geographical locality. For instance, analyses of the cercarial shedding patterns of S. japonicum from two contrasting geographical regions within the Anhui Province of China demonstrated chronobiological polymorphism of cercarial emergence with a significant difference between marshland and hill regions (Lu et al. 2009). A late afternoon shedding peak was observed in all snails from the hill region, while a morning shedding pattern was observed in the majority of the snails from the marshland area. An experimental study of the rhythm of cercarial emergence was conducted to compare two geographical strains of S chistosoma intercalatum from Cameroon and Zaire. For the Cameroon strain, the peak emergence time was between 13:00 and 14:00, and the mean emergence time was 13:59. For the Zaire strain, the peak emergence time was between 12:00 and 13:00, and the mean emergence time was 12:13. These results were shown to be significantly different. Thus, the chronobiology of the emergence rhythms of cercariae seems to be an additional marker to estimate variability and to characterize the two strains (Pages and Theron 1990).
After eggs of O. viverrini are ingested by B. s. goniomphalos, the miracidia hatch and develop further through the stages of sporocysts, rediae, and cercariae about 2 months (Harinasuta and Harinasuta 1984). Our results show that the prevalences of O. viverrini varied with season and the large snails had peaked prevalence in the hot-dry season, while the small size snails had high prevalence in the cool-dry and rainy season. The studies by Chanawong and Waikagul (1991) and Prasopdee et al. (2015) suggested that the small-sized snails (immature stage) had higher infection rates than larger snails. However, other survey studies found that the large-sized snails (mature stage) have high prevalence of O. viverrini infection (Brockelman et al. 1986; Ngern-klun et al. 2006; Upatham and Sukhapanth 1980). Although more study is needed to explain the consequence of O. viverrini infection in different size of B. s. goniomphalos, it is possible that trematode infections are harmful to the mollusk host and can increase snail mortality (Fredensborg et al. 2005; Sorensen and Minchella 1998).
In this study, the peaked prevalence of O. viverrini occurred in the hot-dry season, but in a recent study in the same area, the highest prevalence was found in the cool-dry season (Namsanor et al. 2015). A yearly seasonal variation between years (2012 vs. 2013) is probably the major external factor associated with prevalence, particularly temperature and amount of rainfall, both of which play an important role in the complex interplay between host and parasite (Brockelman et al. 1986). Seasonal changes in temperature, rainfall, and the duration of sunlight cause marked fluctuations in the transmission rates of diseases and parasites (Mouritsen and Poulin 2002; Cattadori et al. 2005; Kim et al. 2005; Altizer et al. 2006).
Seasonal factors can affect the prevalence and number of cercarial produced. Seasonal differences in the number of Schistosoma mansoni cercariae produced per Biomphalaria alexandrina in the Nile Delta were highest from June to September and declined to zero in February and March (Chu and Dawood 1970). Similarly, for both Rhipidocozyle fennica in clams and Diplostomun spathaceum in Lymnaea stagnalis in northern temperate climates, the number of cercariae shed was highest in late summer (Karvonen et al. 2006; Taskinen 1998). O. viverrini cercariae show a circadian rhythm in the emergence of cercariae from snail hosts, but the maximum peak of emergence differs according to seasonal factors and geographical region. The temporal profile of cercariae of O. viverrini s.l. from Sakon Nakhon in the rainy and cool-dry seasons showed that maximum output occurred 2 h earlier than during the hot-dry seasons. The emergence maximum of cercariae is in the morning for Thailand in all seasons, whereas it occurs in the afternoon for Lao PDR except in the hot-dry season (Kiatsopit et al. 2014).
These observed differences reflect an adaptation of the parasite to ecological conditions associated with transmission which are related to geographical origin. The adaptive importance of a rhythm in cercarial emergence is probably related to the facilitation of cercarial transmission to vertebrate host behavior that would make shedding at different times advantageous. The time of shedding is likely to be associated with maximum availability of the second intermediate fish host (Combes et al. 2002). This is particularly relevant if one considers the short lifespan and the decreasing infectivity of cercariae over time (Combes et al. 1994). Such a temporal pattern promotes a high degree of synchrony between parasites and vertebrate hosts. However, there are many factors, age and size of snails, light conditions, temperature, depth of water, and food, that can influence the prevalence and intensity of digenetic trematode infections in the snail intermediate hosts (Fingerut et al. 2003; Graham 2003; Schmidt and Fried 1996; Smyth and Halton 1983).
Cercarial production may be enhanced at higher temperatures as a simple consequence of increased host metabolic activity and the greater energy available to the parasite. The rate of physiological processes increases markedly with increasing temperatures in the ectothermic animals, such as snails (Mas-Coma et al. 2009; Poulin 2006). Galaktionov and Dobrovolskij (2003) showed that temperature and light are the predominant controlling influences. Echinostoma trivolvis cercariae released from naturally infected Helisoma trivolvis snails maintained under different laboratory conditions revealed that temperature was the only significant factor in emergence (Schmidt and Fried 1996). Kaewkes et al. (2012) investigated optimum light intensity with 1000 lx being the most important stimulus on the quantity of O. viverrini cercariae shed from naturally infected B. s. goniomphalos snails in the laboratory.
The rate of development of the trematode in its snail host is dependent on access to ample supplies of food (Kendall 1949). The feeding of snails accelerates parasite maturity so that at a given time many more mature cercariae are produced with well-fed larger snails producing many more cercariae than smaller ones (Belfaiza et al. 2004; Boray 1969; Kendall and Ollerenshaw 1963; Rondelaud et al. 2002, 2004; Seppala et al. 2008). This is in contrast to the finding of Schmidt and Fried (1996), who studied cercarial release of E. trivolvis cercariae from naturally infected H. trivolvis snails maintained under different laboratory conditions. These authors found that food did not affect cercarial release. However, other factors such as the size of the snails can also affect cercarial production (Kiatsopit et al. 2012).
The prevalence of O. viverrini in small snails from Thailand was significantly higher than from medium and large snails in the rainy and cool-dry seasons. This was similar in snails from Lao PDR. This finding could be a result of more susceptibility to infection of smaller snails or also to parasite-induced mortality in snails (Fredensborg et al. 2005; Sorensen and Minchella 1998). The survival of large Lymnaea peregra infected with Echinoparyphium recurvatum was shorter than for infected small snails (Morley et al. 2010). However, the prevalence of O. viverrini cercariae and shell size in the hot-dry season from Thailand is significantly higher in large snails than from medium and small snails. This shows that additional factors such as land use for rice paddy such as irrigation may also influence the relationship between the prevalence of O. viverrini cercariae and shell size (Ngern-klun et al. 2006; Upatham and Sukhapanth 1980).
For the current study, a positive correlation was observed between the size of the snails and the intensity of O. viverrini cercarial infection in all seasons. This indicates that larger snails had the heaviest intensities of infection. Several previous studies have also observed a positive correlation between size with prevalence and intensity of infection (Graham 2003; Kiatsopit et al. 2012). This result is probably explained by the fact that larger snails provide more volume of soft tissue and greater energy resources for the miracidiae or a longer time for sporocyst and cercarial development (Loker 1983; Niemann and Lewis 1990).
The biological evidence of different cercarial emergence patterns from field-collected snail intermediate hosts originating from two geographically separated wetlands from Sakon Nakhon, Thailand and from Vientiane, Lao PDR provides substantial additional evidence that the systematics of B. s. goniomphalos and/or O. viverrini s.l. is much more complex than previously thought. Differences observed may be due to biological differences specific to cryptic species of snails and O. viverrini and/or environmental factors.
References
Altizer S, Dobson A, Hosseini P, Hudson P, Pascual M, Rohani P (2006) Seasonality and the dynamics of infectious diseases. Ecol Lett 9(4):467–484
Anderson RM, May RM (1991) Infectious diseases of humans, dynamics and control. Oxford University Press, New York
Andrews RH, Sithithaworn P, Petney TN (2008) Opisthorchis viverrini: an underestimated parasite in world health. Trends Parasitol 24(11):497–501
Belfaiza M, Rondelaud D, Moncef M, Dreyfuss G (2004) Fasciola hepatica: the effect of food quality on the development of redial generations in Galba truncatula infected with allopatric miracidia. Parasitol Res 92(1):12–17
Boray JC (1969) Experimental fascioliasis in Australia. Adv Parasitol 7:95–210
Boray JC (1978) The potential impact of exotic Lymnaea spp. on fascioliasis in Australasia. Vet Parasitol 4:127–141
Brandt RAM (1974) The non-marine aquatic Mollusca of Thailand. Arch Molluskenkd 105:1–423
Brockelman WY, Upatham ES, Viyanant V, Ardsungnoen S, Chantanawat R (1986) Field studies on the transmission of the human liver fluke, Opisthorchis viverrini, in northeast Thailand: population changes of the snail intermediate host. Int J Parasitol 16:545–552
Cattadori IM, Haydon DT, Hudson PJ (2005) Parasites and climate synchronize red grouse populations. Nature 433(7027):737–741. doi:10.1038/nature03276
Chanawong A, Waikagul J (1991) Laboratory studies on host-parasite relationship of Bithynia snails and the liver fluke, Opisthorchis viverrini. Southeast Asian J Trop Med Public Health 22(2):235–239
Chitramvong YP (1992) The Bithyniidae (Gastropoda: Prosobranchia) of Thailand: comparative external morphology. Malacol Rev 25:21–38
Chu KY, Dawood IK (1970) Cercarial transmission seasons of Schistosoma mansoni in the Nile Delta area. Bull World Health Organ 42:575–580
Combes C, Fournier A, Mone H, Theron A (1994) Behaviours in trematode cercariae that enhance parasite transmission: patterns and processes. Parasitology 109(Suppl):S3–S13
Combes C, Bartoli P, Théron A (2002) Trematode transmission strategies. In: Lewis EE, Campbell JF, Sukhdeo MVK (eds) The behavioural ecology of parasites. CABI Publishing, London, pp 1–12
Dar Y, Rondelaud D, Dreyfuss G (2003) Cercarial shedding from Galba truncatula infected with Fasciola gigantica of distinct geographic origins. Parasitol Res 89(3):185–187
Fingerut JT, Zimmer CA, Zimmer RK (2003) Patterns and processes of larval emergence in an estuarine parasite system. Biol Bull 205(2):110–120
Fredensborg BL, Mouritsen KN, Poulin R (2005) Impact of trematodes on host survival and population density in the intertidal gastropod Zeacumantus subcarinatus. Mar Ecol Prog Ser 290:109–117
Galaktionov KV, Dobrovolskij AA (2003) The biology and evolution of trematodes. Kluwer, Boston
Graham AL (2003) Effects of snail size and age on the prevalence and intensity of avian schistosome infection: relating laboratory to field studies. J Parasitol 89(3):458–463. doi:10.1645/0022-3395(2003)089[0458:EOSSAA]2.0.CO;2
Haas W (1994) Physiological analyses of host-finding behaviour in trematode cercariae: adaptations for transmission success. Parasitology 109:S15–S29
Harinasuta C, Harinasuta T (1984) Opisthorchis viverrini: life cycle, intermediate hosts, transmission to man and geographical distribution in Thailand. Arzneimittelforschung 34(9B):1164–1167
IARC (2011) IARC monographs on the evaluation of carcinogenic risks to humans. World Health Organization, International Agency for Research on Cancer, Lyon
Kaewkes S (2003) Taxonomy and biology of liver flukes. Acta Trop 88:177–186
Kaewkes S, Kaewkes W, Boonmars T, Sripa B (2012) Effect of light intensity on Opisthorchis viverrini cercarial shedding levels from Bithynia snails—a preliminary study. Parasitol Int 61(1):46–48
Karvonen A, Savolainen M, Seppala O, Valtonen ET (2006) Dynamics of Diplostomum spathaceum infection in snail hosts at a fish farm. Parasitol Res 99(4):341–345. doi:10.1007/s00436-006-0137-8
Kendall SB (1949) Nutritional factors affecting the rate of development of Fasciola hepatica in Limnaea truncatum. J Helminthol 23(3–4):179–190
Kendall SB, Ollerenshaw CB (1963) The effect of nutrition on the growth of Fasciola hepatica in its snail host. Proc Nutr Soc 22:41–46
Kiatsopit N, Sithithaworn P, Saijuntha W, Boonmars T, Tesana S, Sithithaworn J, Petney TN, Andrews RH (2012) Exceptionally high prevalence of infection of Bithynia siamensis goniomphalos with Opisthorchis viverrini cercariae in different wetlands in Thailand and Lao PDR. Am J Trop Med Hyg 86(3):464–469
Kiatsopit N, Sithithaworn P, Saijuntha W, Petney TN, Andrews RH (2013) Opisthorchis viverrini: implications of the systematics of first intermediate hosts, Bithynia snail species in Thailand and Lao PDR. Infect Genet Evol 14:313–319. doi:10.1016/j.meegid.2012.12.026
Kiatsopit N, Sithithaworn P, Kopolrat K, Andrews RH, Petney TN (2014) Seasonal cercarial emergence patterns of Opisthorchis viverrini infecting Bithynia siamensis goniomphalos from Vientiane Province, Lao PDR. Parasit Vectors 7:551. doi:10.1186/s13071-014-0551-1
Kiatsopit N, Sithithaworn P, Kopolrat K, Namsanor J, Andrews RH, Petney TN (2015) Trematode diversity in the freshwater snail Bithynia siamensis goniomphalos sensu lato from Thailand and Lao PDR. J Helminthol 6:1–9
Kim K, Dobson AP, Gulland FMD, Harvell CD (2005) Diseases and the conservation of marine biodiversity. In Marine conservation biology. The science of maintaining the sea’s biodiversity. Edited by EA Norse and LB Crowder Marine Conservation Biology Institute, Island Press, Washington, DC: 149–166
Laoprom N, Saijuntha W, Sithithaworn P, Wongkham S, Laha T, Ando K, Andrews RH, Petney TN (2009) Biological variation within Opisthorchis viverrini sensu lato in Thailand and Lao PDR. J Parasitol 95(6):1307–1313
Loker ES (1983) A comparative study of the life-histories of mammalian schistosomes. Parasitology 87(Pt 2):343–369
Lu DB, Wang TP, Rudge JW, Donnelly CA, Fang GR, Webster JP (2009) Evolution in a multi-host parasite: chronobiological circadian rhythm and population genetics of Schistosoma japonicum cercariae indicates contrasting definitive host reservoirs by habitat. Int J Parasitol 39(14):1581–1588
Mas-Coma S, Valero MA, Bargues MD (2009) Climate change effects on trematodiases, with emphasis on zoonotic fascioliasis and schistosomiasis. Vet Parasitol 163(4):264–280. doi:10.1016/j.vetpar.2009.03.024
Morley NJ, Adam ME, Lewis JW (2010) The effects of host size and temperature on the emergence of Echinoparyphium recurvatum cercariae from Lymnaea peregra under natural light conditions. J Helminthol 84(3):317–326. doi:10.1017/S0022149X09990666
Mouritsen KN, Poulin R (2002) Parasitism, community structure and biodiversity in intertidal ecosystems. Parasitology 124:S101–S117
Namsanor J, Sithithaworn P, Kopolrat K, Kiatsopit N, Pitaksakulrat O, Tesana S, Andrews RH, Petney TN (2015) Seasonal transmission of Opisthorchis viverrini sensu lato and a Lecithodendriid trematode species in Bithynia siamensis goniomphalos snails in Northeast Thailand. Am J Trop Med Hyg 93(1):87–93. doi:10.4269/ajtmh.14-0639
Ngern-klun R, Sukontason KL, Tesana S, Sripakdee D, Irvine KN, Sukontason K (2006) Field investigation of Bithynia funiculata, intermediate host of Opisthorchis viverrini in northern Thailand. Southeast Asian J Trop Med Public Health 37(4):662–672
Niemann GM, Lewis FA (1990) Schistosoma mansoni: influence of Biomphalaria glabrata size on susceptibility to infection and resultant cercarial production. Exp Parasitol 70(3):286–292
Pages JR, Theron A (1990) Schistosoma intercalatum from Cameroon and Zaire: chronobiological differentiation of cercarial emergence. J Parasitol 76(5):743–745
Petney T, Sithithaworn P, Andrews R, Kiatsopit N, Tesana S, Grundy-Warr C, Ziegler A (2012) The ecology of the Bithynia first intermediate hosts of Opisthorchis viverrini. Parasitol Int 61(1):38–45. doi:10.1016/j.parint.2011.07.019
Poulin R (2006) Global warming and temperature-mediated increases in cercarial emergence in trematode parasites. Parasitology 132(Pt 1):143–151
Prasopdee S, Kulsantiwong J, Piratae S, Khampoosa P, Thammasiri C, Suwannatrai A, Laha T, Grams R, Loukas A, Tesana S (2015) Temperature dependence of Opisthorchis viverrini infection in first intermediate host snail, Bithynia siamensis goniomphalos. Acta Trop 141(Pt A):112–117. doi:10.1016/j.actatropica.2013.10.011
Rondelaud D, Abrous M, Dreyfuss G (2002) The influence of different food sources on cercarial production in Lymnaea truncatula experimentally infected with Digenea. Vet Res 33(1):95–99
Rondelaud D et al (2004) Variability in the prevalence of infection and cercarial production in Galba truncatula raised on a high-quality diet. Parasitol Res 92(3):242–245
Saijuntha W, Sithithaworn P, Wongkham S, Laha T, Pipitgool V, Tesana S, Chilton NB, Petney TN, Andrews RH (2007) Evidence of a species complex within the food-borne trematode Opisthorchis viverrini and possible co-evolution with their first intermediate hosts. Int J Parasitol 37(6):695–703
Saijuntha W, Sithithaworn P, Kaitsopit N, Andrews RH, Petney TN (2014) Liver flukes: Clonorchis and Opisthorchis. Adv Exp Med Biol 766:153–199. doi:10.1007/978-1-4939-0915-5_6
Schmidt KA, Fried B (1996) Emergence of cercariae of Echinostoma trivolvis from Helisoma trivolvis under different conditions. J Parasitol 82(4):674–676
Seppala O, Liljeroos K, Karvonen A, Jokela J (2008) Host condition as a constraint for parasite reproduction. Oikos 117:749–753
Sithithaworn P, Andrews RH, Nguyen VD, Wongsaroj T, Sinuon M, Odermatt P, Nawa Y, Liang S, Brindley PJ, Sripa B (2012) The current status of opisthorchiasis and clonorchiasis in the Mekong Basin. Parasitol Int 61(1):10–16. doi:10.1016/j.parint.2011.08.014
Smyth JD, Halton DW (1983) Trematoda; Platyhelminthes; Physiology. Cambridge University Press, New York
Sorensen RE, Minchella DJ (1998) Parasite influences on host life history: Echinostoma revolutum parasitism of Lymnea elodes snails. Oecologia 115:188–195
Taskinen J (1998) Cercarial production of the trematode Rhipidocotyle fennica in clams kept in the field. J Parasitol 84(2):345–349
Upatham ES, Sukhapanth N (1980) Field studies on the bionomics of Bithynia siamensis siamensis and the transmission of Opisthorchis viverrini in Bangna, Bangkok, Thailand. Southeast Asian J Trop Med Public Health 11(3):355–358
Upatham ES, Sornmani S, Kitikoon V, Lohachit C, Bruch JB (1983) Identification key for fresh-brackish water snails of Thailand. Malacol Rev 16:107–132
Wongratanacheewin S, Pumidonming W, Sermswan RW, Maleewong W (2001) Development of a PCR-based method for the detection of Opisthorchis viverrini in experimentally infected hamsters. Parasitology 122(Pt 2):175–180
Acknowledgments
We acknowledge the support of the Faculty of Medicine, Khon Kaen University, Visiting International Professor Program. This work was supported by the Higher Education Research Promotion and office of the Higher Education Commission, through health cluster (SHeP-GMS), the Post-Doctoral Training, Graduate School Khon Kaen University, Thailand, the Thailand Research Fund through the Basic Research Grant and Cholangiocarcinoma Screening and Care Program (CASCAP), Khon Kaen University. We would like to thank the Deutsche Forschungsgemeinschaft (PE1611/1-3), the National Research Council of Thailand, and the International Excellence Fund of Karlsruhe Institute of Technology as well as ASEAN-EU Year of Science, Technology and Innovation, 2012 for providing funding for cooperative workshops.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Laoprom, N., Kiatsopit, N., Sithithaworn, P. et al. Cercarial emergence patterns for Opisthorchis viverrini sensu lato infecting Bithynia siamensis goniomphalos from Sakon Nakhon Province, Thailand. Parasitol Res 115, 3313–3321 (2016). https://doi.org/10.1007/s00436-016-5089-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-016-5089-z