
Abstract
The present study investigated how response mode determines the specificity of control responsible for the congruency sequence effect (CSE), especially when conflict arises from spatial dimensions. Horizontal and vertical Simon tasks were presented in turn, while response mode (Experiment 1) or task-relevant stimulus dimension (Experiment 2) was manipulated. All responses were made by aimed movements to make the relative salience of the horizontal and vertical dimensions equivalent regardless of response mode. The confound-minimized CSEs were significant only when the two tasks shared the same response mode, which did not vary as a function of task-relevant stimulus dimension. This result suggests that response mode determines the scope of control, as it reconfigures the representations of the task-irrelevant spatial dimensions (i.e., the horizontal and vertical dimensions), which is corroborated by distributional analyses. This response mode-specific control was also consistently found for the horizontal and vertical arrow versions of flanker-compatibility tasks in Experiment 3, in which conflict does not directly arise from the response dimension. Furthermore, the current findings revealed that the CSEs were more evident in movement times than in initiation times, which provides new insight on how control inhibits the response activated by a task-irrelevant stimulus dimension, especially at a motor level.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
A cognitive control mechanism enables efficient performance as it facilitates the selective processing of information that is relevant to the current task goal. When task-irrelevant information activates a response that does not correspond to the response activated by task-relevant information, cognitive conflict occurs, leading to slower response times or more frequent errors, which is called the congruency effect. For example, in a Simon task, participants are instructed to make a left or right response to a non-spatial feature of the target stimulus (e.g., color or shape) that is randomly presented at either the left side or right side of a display. Even though the target location is irrelevant to the task, performance is impaired when it does not match the correct response location (Simon & Rudell, 1967). This Simon effect is found not only when the target appears along the horizontal axis (horizontal Simon task) but also when it appears along the vertical axis (vertical Simon task, e.g., Stürmer, Leuthold, Soetens, Schröter, & Sommer, 2002).
Gratton, Coles, and Donchin (1992) found that the magnitude of the congruency effect is larger following congruent trials than following incongruent trials, which is referred to as the congruency sequence effect (CSE). A wealth of research suggests that an executive control mechanism reactively modulates the magnitude of the congruency effect depending on the previous trial’s congruency (e.g., Botvinick, Braver, Barch, Carter, & Cohen, 2001; Egner & Hirsch, 2005; Ridderinkhof, 2002). However, it remains unclear how the control mechanism specifically operates after detecting conflict. To elaborate on the nature of the control mechanism, many studies adopted a cross-task design, in which two different tasks are presented alternately in a trial-by-trial manner (for a review, see Braem, Abrahamse, Duthoo, & Notebaert, 2014). By examining what kinds of task feature the two tasks should have in common to obtain the CSE between them, cross-task experimental designs provide insights into what is controlled during the sequential modulation and what determines its boundary.
Some researchers have proposed that the control mechanism modulates the size of the congruency effect, because it facilitates the processing of task-relevant stimulus dimensions (Egner & Hirsch, 2005; Notebaert & Verguts, 2008; Verguts & Notebaert, 2008, 2009). For example, Verguts and Notebaert (2008) suggested that the currently activated associations between the task-relevant stimulus information and its corresponding response are strengthened after the conflict is detected. Eventually, the detection of conflict leads to the enhanced processing of the task-relevant stimulus dimension. In contrast, others have proposed that the control mechanism suppresses the processing of task-irrelevant stimulus dimensions (e.g., Lee & Cho, 2013; Stürmer et al., 2002). In this case, the association between the task-irrelevant stimulus information and its incorrectly activated response is to be inhibited.
However, previous findings also suggested that the boundary of control does not simply depend on the task-relevant or task-irrelevant stimulus dimensions. For example, when the horizontal and vertical Simon tasks are presented in turn, sequential modulation between the two tasks has not been consistently observed. Even though the tasks shared both the task-relevant and task-irrelevant stimulus dimensions, which are non-spatial and spatial, respectively, the cross-task CSE was found in some studies (e.g., Braem, Verguts, & Notebaert, 2011; Weissman, 2019a, b; Wühr, 2005; Wühr, Duthoo, & Notebaert, 2015, Experiment 1), but not in others (e.g., Hoppe, Küper, & Wascher, 2017; Lee & Cho, 2013; Wühr et al., 2015, Experiment 2).
The major difference in those contradictory results possibly lies in how response alternatives are represented. To perform the horizontal and vertical Simon tasks, several types of different manual responses, such as keypress, aimed-movement, or joystick responses, have been used. Each response type involves different response configurations, which affect how the corresponding spatial dimensions (i.e., horizontal and vertical) are cognitively coded (Adam, Hommel, & Umiltà, 2003; Ansorge & Wühr, 2004; Hommel, 2011; Vu, Proctor, & Pick, 2000). Because these spatial dimensions are the main source of the Simon-type conflict, the scope of control possibly varies depending on the response representation, as it changes the cognitive coding of the spatial dimensions. Especially, the left- and right-hand keypress responses involve a different response configuration from the unimanual aimed-movement or joystick responses, in terms of response mode and relative salience of the spatial dimensions.
The response mode is a representational group of response alternatives sharing similar properties or being distinguished by a salient distinction, such as anatomical feature, relative response location, spatial arrangements of stimuli, spatial cues, or task representation (Adam, 1994; Adam et al., 2003; Ansorge & Wühr, 2004; Freedberg, Wagschal, & Hazeltine, 2014; Hazeltine, 2005; Kim & Cho, 2014; Lippa, 1996; Reeve & Proctor, 1984). For example, when participants make directional responses with unimanual joystick or aimed-movement responses, the representations of the four response alternatives (i.e., left, right, above, and below) are supposedly grouped into a single response mode. In contrast, when the horizontal and vertical response arrays are separately assigned to the left and right hands using keypress responses, the horizontal and vertical response sets are likely to form two respective response modes (Kim & Cho, 2014; Proctor & Vu, 2010). This is because the specification of hand provides a relatively salient distinction in terms of both the spatial separation (Reeve & Proctor, 1984) and the hierarchy of response features (Miller, 1982; Rosenbaum, 1983). Previous studies suggested that response mode modulates the specificity of control, as the CSE was evident between two tasks when they shared a common response mode, but not when they had different response modes (Braem et al. 2011; Hazeltine, Lightman, Schwarb, & Schumacher, 2011; Janczyk & Leuthold, 2018; Kim & Cho, 2014; Lim & Cho, 2018). Nevertheless, the specific role of response mode in determining the boundary of control has not clearly unfolded.
Especially for the Simon task, response mode is expected to play an important role, because the spatial coding of the horizontal and vertical dimensions varies depending on how the spatially corresponding responses are represented (Adam et al., 2003; Vu, Pellicano, & Proctor, 2005; Vu et al., 2000). It has been suggested that spatial codes are determined by response-discriminating features, as significant horizontal or vertical Simon effects were observed only when response alternatives were discriminated in terms of horizontal or vertical dimension, respectively (Ansorge & Wühr, 2004; Hommel, 2011). In this regard, when the response sets of the horizontal and vertical Simon tasks are grouped as a same response mode, the horizontal and vertical stimulus dimensions are also likely to be represented as a single spatial dimension consisting of four directions. This is because the distinction between the horizontal and vertical dimensions is not clear or necessary, as there is no salient or relevant reference to generate the horizontal and vertical spatial codes separately (Ansorge & Wühr, 2004; Vu & Proctor, 2001, 2002). Conversely, when different response modes are employed, the distinction between the horizontal and vertical responses would become clear, and thus, the horizontal and vertical dimensions are presumably represented as two separate spatial dimensions. Indeed, the previous studies obtained the sequential modulation between the horizontal and vertical Simon tasks when they were performed with the same response mode, using unimanual joystick response, aimed-movement response, or the same response set (Lee & Cho, 2013, Experiment 4; Wühr et al., 2015, Experiment 1). However, no CSE was found when they were performed with different response modes (Hoppe et al., 2017; Lee & Cho, 2013, Experiment 1).
The response configuration causes another disparity in the relative salience of the horizontal and vertical dimensions (Proctor, Vu, & Nicoletti, 2003; Vu et al., 2005). For example, when keypress responses are used, the response alternatives of the vertical Simon tasks are usually assigned to the left and right hands, of which placements are distinguished in terms of the horizontal dimension, as well as the vertical dimension. However, because the two hands are horizontally distinguished by the body midline, the horizontal dimension is likely to be more salient than the vertical dimension, even in the vertical Simon task (Proctor et al., 2003). Accordingly, the strength of the automatic link between stimulus and response locations in the vertical Simon task may not be equivalent to that in the horizontal Simon task. This may explain why the magnitude of the horizontal Simon effect was larger than that of the vertical Simon effect in the previous studies where the vertical response alternatives were distinguished by the left–right distinction (e.g., Proctor et al., 2003, Experiment 3; Rubichi, Nicoletti, & Umiltà, 2005).
However, when joystick or aimed-movement responses are employed, the horizontal response alternatives only involve horizontal spatial codes, whereas the vertical response alternatives only involve vertical ones. Therefore, the horizontal and vertical spatial correspondences become more equivalent, resulting in a similar magnitude of the horizontal and vertical Simon effects (Hommel, 1996; Vu et al., 2005, Experiment 3; Vu & Proctor, 2001). This is because the response alternatives in the vertical Simon tasks are not discriminated by horizontally aligned effectors, and thus, the frame of reference for the horizontal dimension is not favored over that for the vertical dimension.
This asymmetry in the spatial correspondence for the horizontal and vertical Simon tasks can make qualitative (e.g., Wiegand & Wascher, 2007) or quantitative (e.g., Vu et al., 2005) differences, as corroborated by response time distributional analyses (for a review, see Proctor, Miles, & Baroni, 2011). When plotting the averaged Simon effect for each response time quintile, the size of the horizontal Simon effect typically decreases as the response time gets slower, resulting in a decreasing delta function. On the other hand, the delta function of the vertical Simon effect remains constant or increases (e.g., Proctor et al., 2003; Wiegand & Wascher, 2005). This difference implies that the strength of the automatic association between the stimulus and response locations is not comparable for the horizontal and vertical Simon tasks, because the decreasing Simon effect presumably reflects the response activation from the task-irrelevant stimulus location, which is automatically formed and diminishes rapidly over time (De Jong, Liang, & Lauber, 1994).
The purpose of the present study was to examine what determines the specificity of control process responsible for the CSE, especially when the task-irrelevant spatial information varies along the horizontal and vertical dimensions. Because the horizontal and vertical tasks consisting of different stimulus and response sets were presented on alternate trials, the cross-task CSE between previous and current trials was not confounded with the repetition priming effect (Hommel, Proctor, & Vu, 2004; Mayr, Awh, & Laurey, 2003) or contingency learning effect (Schmidt & De Houwer, 2011), of which avoidance has been an important concern in research on the CSE (for review, Braem et al., 2019; Schmidt, 2019). Aimed-movement responses were adopted to make the relative salience of the vertical dimension more equivalent to that of the horizontal dimension, regardless of whether the horizontal and vertical tasks were performed with the same or different response modes. In this way, the current study could determine more clearly whether the response activated by the task-irrelevant horizontal and vertical locations can be regulated by the same control mechanism when one dimension is not dominant over the other dimension. Moreover, as aimed-movement responses allow one to examine how the competing response activations are processed even after the movement is initiated (Doucet & Stelmack, 1999; Hietanen & Pia, 1995; Rubichi & Pellicano, 2004), the current study provides further information about how the control mechanism keeps operating during and after an abstract level of response selection.
In Experiment 1, the horizontal and vertical Simon tasks were presented in turn, while response mode was manipulated. If response mode determines the specificity of control triggered by spatial conflict, because it reconfigures the representations of spatial dimensions, the CSE would be found when the same response mode was adopted to perform the two tasks, but not when different response modes were adopted. However, there is another possibility that response mode modulates the cross-task CSE by reconfiguring the two task sets to be represented as the same or different task sets, rather than by reconfiguring the representations of spatial dimensions. To test this possibility, in Experiment 2, we manipulated task-relevant stimulus dimension, which is another critical factor determining task sets. If the distinction between the two task sets plays a crucial role in determining the scope of control, the CSE should disappear when the two tasks have different task-relevant stimulus dimensions despite the use of the same response mode. To generalize the role of response mode to tasks not involving the conflict from the response dimension, in Experiment 3, horizontal and vertical versions of flanker-compatibility tasks were performed with the same or different response modes. If the specificity of the control mechanism depends on response mode, even though conflict is not directly derived from the response dimension, the CSE would be significant between the tasks only for the same response mode.
Experiment 1
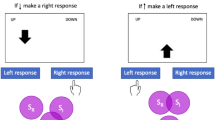
The goal of Experiment 1 was to examine whether the CSE between horizontal and vertical Simon tasks varies depending on response mode as it reconfigures the spatial coding of the horizontal and vertical dimensions. To make the strength of the automatic link between spatial codes for stimulus and response locations in the vertical Simon task more equivalent to that in the horizontal Simon task, aimed-movement responses were adopted. Half of the participants performed the horizontal and vertical Simon tasks with the same, while the other half performed with different response modes. For the same mode, participants were to perform both the horizontal and vertical Simon tasks with their right index finger. For different modes, they responded to the horizontal Simon task with their left index finger and the vertical Simon task with their right index finger (see Fig. 1).

Illustrations of the stimulus and response sets used in Experiment 1. The horizontal and vertical Simon tasks consisting of different color stimuli were presented alternately in a trial-by-trial manner. The horizontal and vertical responses, denoted by black and gray arrows, respectively, were carried out with the same or different response modes
Regardless of response mode, the horizontal and vertical Simon tasks always shared the same task-relevant stimulus dimension, as participants were instructed to make a response to the color of the target in both tasks. Therefore, if the scope of control is modulated by response mode, a CSE should occur between the two tasks when they had the same response mode but not when they had different response modes. However, if the control mechanism is independent of response mode and applicable to both the horizontal and vertical Simon tasks, sequential modulation should be observed irrespective of response mode. Alternatively, if the control mechanism is independent of response mode, but cannot be transferred from the horizontal to the vertical Simon tasks, and vice versa, the CSE would not be evident regardless of response mode.
Another prediction about the role of the response mode was that it would determine whether the horizontal and vertical dimensions are represented as the same or different dimensions. For this reason, distributional analyses were conducted to examine whether a decreasing delta function, which is typically observed in the horizontal Simon task, would be found in the vertical Simon task, as well. If the stimulus alternatives in the horizontal and vertical dimensions are spatially coded as a single dimension by employing the same response mode, the same pattern of the delta function would be obtained in the horizontal and vertical Simon tasks only when they were responded by the same response mode.
Method
Participants. To detect the difference in the CSE depending on response mode with a power 1-β = 0.95 at an α = 0.05, a power analysis using the G*Power 3.1 (Faul, Erdfelder, Buchner, & Lang, 2009) for mixed-measures analyses of variance (ANOVAs), as a function of n-1 congruency (congruency of the previous trial), congruency, and response mode (a correlation among repeated measures = 0.5), resulted in a minimum sample size of 22, assuming the effect size as η2p = 0.0979 based on a similar experiment by Lim and Cho (2018, Experiment 2). Given a possible difference in the effect-size estimation for aimed-movement responses, a larger number of participants were employed. 32 participants (15 males, mean age = 23.13) recruited from Korea University participated in Experiment 1. Sixteen participants performed the horizontal and vertical Simon tasks with the same mode, whereas the other 16 participants performed the tasks with different modes. In this experiment and in Experiments 2 and 3, all participants self-reported to have no deficit in visual acuity and color vision, signed an informed consent before performing the task, and received KRW 6,000 (about 5 US dollars) after the participation. The current and following experiments were approved by the Institutional Review Board at Korea University (KU-IRB-16-142-A-1).
Stimulus and apparatus. Visual stimuli were presented on a 17-inch CRT monitor of a personal computer, and viewing distance was about 60 cm. The experiment was programmed and controlled by Matlab software (version 2015a), using Psychtoolbox3. A white (R = 255, G = 255, B = 255) cross (approximately 0.3° × 0.3° of visual angle) appeared at the center of the display as a fixation point. For the horizontal Simon task, a square (approximately 1.51° × 1.51°), which was colored in red (R = 255, G = 0, B = 0) or yellow (R = 255, G = 255, B = 0), was presented at either left side or right side of the fixation cross. For the vertical Simon task, a green (R = 0, G = 255, B = 0) or blue (R = 0, G = 0, B = 255) square was presented either above or below the fixation cross. The target stimulus appeared at an equal distance from the center of the display (approximately 5.4°). All stimuli were displayed on the gray background (R = 128, G = 128, B = 128).
The aimed-movement responses were recorded by a standard 101-key computer keyboard. For the same mode, the direction keys for the ‘left’, ‘right’, ‘up’, and ‘down’ were the ‘4’, ‘6’, ‘8’, and ‘2’ keys on the numeric keypad of the computer keyboard, respectively, and the home key was the ‘5’ key on it. Participants were asked to keep the home key pressed with their right index finger until making a response to the target and to make a response to the target by pressing one of the direction keys with the same finger. For different modes, they were instructed to make responses by pressing the ‘a’ and ‘d’ keys as the ‘left’ and ‘right’ direction keys with their left index finger and the ‘8’ and ‘2’ keys on the numeric keypad as the ‘up’ and ‘down’ direction keys with their right index finger. The ‘s’ and ‘5’ keys were used as the home keys for the left and right hands, respectively. Both home keys were to be pressed simultaneously with the left and right index fingers until the target appeared. Then, only the corresponding finger was to be lifted to press a direction key, while the other one was to keep pressing the home key. All participants were instructed to press the ‘left’ direction key to the red, the ‘right’ direction key to the yellow, the ‘up’ direction key to the green, and the ‘down’ direction key to the blue target square.
Procedure. After receiving the informed consent, the instruction for the experimental procedures was provided in a dimly lit sound-proof chamber. The body midline of the participant was aligned with the center of the monitor and the midline of the numeric keypad when the same mode was employed or the whole keyboard when the different modes were employed. When each trial began with the fixation cross, participants were instructed to keep pressing the home key(s) until the target appeared and its corresponding response was determined. 500 ms after the home key was pressed, the target stimulus was presented until any home key was released or for a maximum of 250 ms, which was followed by a blank display for 1750 ms. If any home key was released before the target stimulus appeared, a visual feedback message of “Press the home key” was presented. As soon as the home key was released after the onset of the target display, the display screen went blank until participants pressed a direction key or for 2000 ms.
Participants were instructed to press one of the four direction keys depending on the color of the target stimulus, with either the same or different modes (see Fig. 1). Regardless of whether they were to respond with the same or different modes, all participants were not explicitly instructed that the two different tasks (i.e., the horizontal and vertical Simon tasks) would be presented in turn. The instructions basically informed of the four stimulus–response mappings between the stimulus colors and response locations (i.e., red—left, yellow—right, green—up, and blue—down). Also, note that participants were explicitly emphasized not to release the home key before they finished deciding a direction key to prevent them from deliberately delaying the whole decision process after releasing the home key (Rubichi & Pellicano, 2004; Smith & Carew, 1987). Then, a 150-ms tone was sounded when participants pressed a wrong direction key, did not release the home key within 2000 ms after the target onset, or did not press any direction key within 2000 ms after releasing the home key. After that, another blank display was presented for 1000 ms before the beginning of the next trial. Participants were asked to make a response as rapidly and accurately as possible.
The main experiment was composed of 8 blocks of 82 trials, preceded by a practice block of 34 trials. A 1-min break was given between blocks. The congruency of the first and second trials was randomly determined; for the remaining trials within the blocks, the sequence of congruency was pseudo-randomly determined, so that the numbers of the congruent trials after a congruent trial (cC), congruent trials after an incongruent trial (iC), incongruent trials after a congruent trial (cI), and incongruent trials after an incongruent trial (iI) were equated for the horizontal and vertical Simon tasks, respectively.
Results
Initiation time (IT) and movement time (MT) were recorded separately. IT was defined as the time between the target onset and the moment when the home key was released to initiate the response. The temporal interval between when participants released the home key and when they pressed a direction key was measured as MT. Considering a trade-offFootnote 1 between IT and MT, outliers were defined as the trials of which either MT or IT was more than 3 standard deviations away from its conditional mean for each individual participant. The first two trials of each block, outliers, and the trials following outliers or incorrect trials were excluded from the analyses (about 8.63% of the total trials). Mean correct ITs, MTs, and percent errors (PEs) were calculated for each participant as a function of n-1 congruency (congruent vs. incongruent) and current congruency (congruent vs. incongruent; see Table 1). Then, the mean correct ITs, MTs, and PEs were submitted to three-way repeated-measures analyses of variance (ANOVAs), with the above factors as within-subject variables and response mode (same vs. different) as a between-subjects variable.
IT. The mean IT was significantly greater after incongruent trials (M = 449 ms) than after congruent trials (M = 446 ms), F(1, 30) = 5.77, p = 0.0227, MSE = 54, η2p = 0.16. This post-conflict slowing is often found in congruency tasks and indicates that responses become slower after experiencing conflict to reduce the risk of making an incorrect response (Verguts, Notebaert, Kunde, & Wühr, 2011). The post-conflict slowing was modulated by response mode, F(1, 30) = 11.43, p = 0.002, MSE = 54, η2p = 0.28. It was evident with the same mode (7 ms), F(1, 15) = 15.81, p = 0.0012, MSE = 57, η2p = 0.51, but not with different modes (– 1 ms), F(1, 15) = 0.51, p = 0.49, MSE = 51, η2p = 0.03.
A significant Simon effect was observed, as a main effect of current congruency was significant, F(1, 30) = 55.79, p < 0.001, MSE = 227, η2p = 0.65. Participants made slower responses on incongruent trials (M = 458 ms) than congruent trials (M = 438 ms). The overall CSE was not found between the two task congruencies, as indicated by no interaction between n-1 congruency and current congruency, F(1, 30) = 0.34, p = 0.56, MSE = 38, η2p = 0.01. The three-way interaction of n-1 congruency, current congruency, and response mode was also not significant, F(1, 30) = 0.04, p = 0.84, MSE = 38, η2p = 0.0014 (see Fig. 2).

MT. A significant Simon effect was obtained, F(1, 30) = 33.67, p < 0.001, MSE = 102, η2p = 0.53, as the mean MT was longer on incongruent trials (M = 136 ms) than on congruent trials (M = 126 ms). More importantly, current congruency interacted with n-1 congruency, F(1, 30) = 6.65, p = 0.0151, MSE = 34, η2p = 0.18, indicating a significant CSE. The magnitude of the Simon effect was reduced after incongruent trials (8 ms), F(1, 30) = 16.13, p < 0.001, MSE = 59, η2p = 0.35, compared to after congruent trials (13 ms), F(1, 30) = 34.94, p < 0.001, MSE = 78, η2p = 0.54. Furthermore, the CSE was modulated by response mode, F(1, 30) = 5.17, p = 0.0303, MSE = 34, η2p = 0.15. Separate analyses on each response mode demonstrated that the CSE was evident only when the same mode was used, F(1, 15) = 9.68, p = 0.0071, MSE = 42, η2p = 0.4. For the same mode, a smaller Simon effect was obtained after incongruent trials (9 ms), F(1, 15) = 5.7, p = 0.0305, MSE = 101, η2p = 0.28, than congruent trials (19 ms), F(1, 15) = 33.69, p < 0.001, MSE = 82, η2p = 0.7. However, for different modes, no sequential modulation was obtained, F(1, 15) = 0.06, p = 0.81, MSE = 27, η2p = 0.0039 (see Fig. 2).
PE. The overall PE was 2.18%. The only effect that reached significance was the main effect of current congruency, F(1, 30) = 38.63, p < 0.001, MSE = 2.33, η2p = 0.56, indicating a 1.68% Simon effect.
Distributional analyses. To compare the temporal dynamics of the horizontal and vertical Simon effects, distributional analyses were conducted on the IT and MT data. Conditioned on task (horizontal vs. vertical) and current congruency (congruent vs. incongruent), each participant’s responses were divided into quintiles based on total response time, which was the sum of IT and MT. For each quintile, ITs and MTs data were averaged, respectively, and the quintile-averaged ITs and MTs of congruent trials were subtracted from those of incongruent trials to calculate the delta functions (see Fig. 3). To test the statistical difference between the delta functions of the horizontal and vertical Simon effects depending on response mode, quintile-averaged data were entered into four-way repeated-measures ANOVAs with response mode (same vs. different) as a between-subject variable and task (horizontal vs. vertical Simon task), congruency (congruent vs. incongruent), and quintile (1–5) as within-subject variables, after the Greenhouse–Geisser correction if necessary.

The magnitude of Simon effects across quintiles for the horizontal (solid lines) and vertical (dashed lines) Simon tasks in Experiment 1. ITs (black lines) and MTs (gray lines) were plotted respectively, for each response mode (upper vs. below)
The results showed the overall decrease in the Simon effect across quintiles, in the IT data, F(2, 60) = 25.5, p < 0.001, MSE = 393, η2p = 0.46, but not in the MT data, F(2, 51) = 2.02, p = 0.15, MSE = 340, η2p = 0.06. The three-way interaction of task, current congruency, and quintile were not significant either in the IT, F(2, 72) = 1.6, p = 0.21, MSE = 315, η2p = 0.05, or MT, F(2, 47) = 0.59, p = 0.52, MSE = 541, η2p = 0.02. However, most importantly, the four-way interaction among task, current congruency, quintile, and response mode was significant in the IT data, F(2, 72) = 4.87, p = 0.007, MSE = 315, η2p = 0.14, but not in the MT data, F(2, 47) = 1.97, p = 0.16, MSE = 541, η2p = 0.06. These results suggest that the delta functions of the horizontal and vertical Simon effects showed similar or different patterns depending on response mode in the IT data.
For the further examination of the four-way interaction in the IT data, separate analyses were conducted for each response mode. When the horizontal and vertical Simon tasks shared the same mode, the Simon effect decreased across the quintiles in the IT data, F(2, 34) = 22.37, p < 0.001, MSE = 240, η2p = 0.6, and it was not modulated by task, F(2, 34) = 1.76, p = 0.25, MSE = 294, η2p = 0.11. These results indicate that the decreasing delta function was evident in the vertical Simon task, F(2, 28) = 11.71, p < 0.001, MSE = 440, η2p = 0.44, as well as in the horizontal Simon task, F(4, 60) = 9.12, p < 0.001, MSE = 96, η2p = 0.38, when the same mode was used. For different modes, the Simon effect varied across quintiles, F(2, 26) = 7.95, p = 0.003, MSE = 594, η2p = 0.35. Moreover, this interaction between congruency and quintile was modulated by task, F(2, 36) = 4.65, p = 0.012, MSE = 354, η2p = 0.24, which means that the Simon effect varied differently depending on task. The separate analyses for each task showed a significant decrease in the horizontal Simon effect, F(2, 35) = 16.17, p < 0.001, MSE = 283, η2p = 0.52, but not in the vertical Simon effect, F(2, 25) = 1.15, p = 0.32, MSE = 716, η2p = 0.07. These findings suggest that the decreasing delta function was evident in the horizontal Simon task, but not in the vertical Simon task, when responded to by different modes.
Discussion
The sequential modulation between the horizontal and vertical Simon effects was significant in the MT. Furthermore, response mode modulated the CSE found in the MT data, as it was significant when the tasks were performed with the same response mode, but not with different response modes. Even though the magnitude of the Simon effect was smaller with different modes than with the same mode, separate analyses on each response mode showed that the Simon effect was still significant when different modes were employed. Moreover, because the CSE is not proportionate to the size of the congruency effect (Weissman, Jiang, & Egner, 2014), it is unlikely that the lack of the CSE was completely attributable to a relatively small amount of conflict using different modes.
Considering the automatic association between stimulus and response locations, which is the main source of Simon-type conflict, it is possible that the spatial representation of the horizontal and vertical stimulus dimensions varies depending on response mode. When the distinction of response sets between the horizontal and vertical Simon tasks was clear using different modes, their spatial dimensions seemed to be represented separately as horizontal and vertical dimensions, resulting in no cross-task CSE. On the contrary, when the same mode was employed, the distinction between the horizontal and vertical axes was less clear. Thus, the spatial representations of the two dimensions were unlikely to be differentiated, which allowed them to share a common control mechanism.
The distributional analyses corroborate the evidence that the processing of the horizontal and vertical dimensions varies depending on response mode. For different modes, the Simon effect decreased across quintiles only in the horizontal Simon task, but not in the vertical Simon task, which is consistent with the previous findings showing different patterns of delta functions between the horizontal and vertical Simon tasks (e.g., Proctor et al., 2003). Proctor et al. demonstrated that the response activation from the vertical spatial stimulus codes was not comparable to that from the horizontal spatial stimulus codes, even when one dimension was not dominant over the other dimension. When using the same mode, on the other hand, the current study observed a decreasing delta function in the vertical Simon task, as well as in the horizontal Simon task. It is possible that the horizontal and vertical spatial stimulus codes decayed or were suppressed in a similar manner (Hommel, 1997; Ridderinkhof, 2002). This finding is consistent with the idea that the spatial codes for the horizontal and vertical dimensions are not distinguished in terms of axes when responding with the same mode, as the axes do not provide a salient frame of reference any more (Ansorge & Wühr, 2004; Hommel, 2011; Proctor & Reeve, 1985). Therefore, the horizontal and vertical spatial codes are more likely to be represented as four spatial codes that belong to a single spatial dimension.
Experiment 2
Experiment 1 provided evidence for the idea that the scope of control depends on response mode as it reconfigures the spatial representations of the horizontal and vertical dimensions. However, some may argue that the different response mode renders not only the distinction between the horizontal and vertical axes but also the distinction between the horizontal and vertical Simon tasks clearer. Indeed, the previous studies proposed that two tasks need to be represented as a single task to engender the cross-task CSE (Akçay & Hazeltine, 2008; Braem et al., 2014; Hazeltine et al., 2011). Also, Lim and Cho (2018) suggested that response mode reconfigures the task representation in flanker-compatibility tasks. Therefore, we cannot rule out the possibility that the scope of control is modulated by response mode, because it reconfigures whether the horizontal and vertical tasks are represented as a single task set or two different task sets in general, rather than more specifically reconfigures their spatial dimensions.
To examine this possibility, Experiment 2 manipulated the task-relevant stimulus dimension, which is another essential component to construct task representations (Rogers & Monsell, 1995), but employed the same response mode throughout the experiment. When the horizontal and vertical Simon tasks shared the same task-relevant stimulus dimension, participants were to identify a letter stimulus in both tasks. In contrast, when they had different task-relevant stimulus dimensions, participants were to respond to the identity of a letter stimulus in the horizontal Simon task and the color of the target stimulus in the vertical Simon task (see Fig. 4). If the cross-task CSE is dependent on task representations when two task sets are clearly distinguished, then sequential modulation should occur only when the tasks shared the same task-relevant stimulus dimension. Alternatively, if the scope of control depends on response mode as it reconfigures the representation of spatial dimension (i.e., task-irrelevant stimulus dimension), the CSE would be observed between the horizontal and vertical Simon tasks, regardless of whether they shared the same task-relevant stimulus dimension or not.

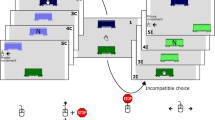
Illustrations of the stimulus sets used in Experiment 2. For the same dimension condition (left), the horizontal and vertical Simon tasks consisting of different letter stimuli were presented in turn. For different dimension condition (right), a colored square appeared in the horizontal Simon task, while a letter stimulus appeared in the vertical Simon tasks
Method
Participants. Thirty-two participants (16 males, mean age = 23.28) were recruited from the same pools as in Experiment 1. Sixteen participants performed the horizontal and vertical Simon tasks having the same task-relevant stimulus dimension, while the other 16 participants performed the two tasks having different task-relevant stimulus dimensions.
Stimulus and apparatus. The apparatus and stimuli were the same as in Experiment 1, with the following differences. For the same stimulus dimension, a set of letter stimuli was used in both the horizontal and vertical Simon tasks. When a letter ‘T’ or ‘L’ was presented in the horizontal Simon task, participants were to press the ‘left’ or ‘right’ direction key, and when a letter ‘H’ or ‘N’ appeared in the vertical Simon task, they were to press the ‘up’ or ‘down’ direction key, respectively. For the horizontal and vertical Simon tasks having different stimulus dimensions, the task-relevant stimulus dimension was the color of the target in the horizontal task and the identity of the letter in the vertical task. This group of participants was asked to make the ‘left’ or ‘right’ response to a red (R = 255, G = 0, B = 0) or yellow (R = 255, G = 255, B = 0) square (approximately 0.72° × 0.72°) in the horizontal Simon task, and the ‘up’ or ‘down’ response to a letter ‘T’ or ‘L’ (approximately 0.72° × 0.72°) in the vertical Simon task, respectively. Since the two tasks were always responded to with the same mode, ‘left’, ‘right’, ‘up’, and ‘down’ direction keys were allocated to the ‘4’, ‘6’, ‘8’, and ‘2’ keys on the numeric keypad of a standard personal computer 101-key keyboard, respectively. The home key was the ‘5’ key on a numeric keypad and all keys were pressed with the right index finger.
Procedure. All procedures were identical to those of Experiment 1 with the following exceptions. For the same stimulus dimension, participants were to perform the letter Simon task both in horizontal and vertical Simon tasks. For different stimulus dimensions, they were to perform the horizontal color Simon task and the vertical letter Simon task (see Fig. 4). Both for the same and different stimulus dimensions, the instructions were always based on the four stimulus–response mappings, without explicitly informing that participants would perform two different tasks (i.e., the horizontal and vertical Simon tasks) in turn. All participants performed the horizontal and vertical Simon tasks with the same mode.
Results
IT and MT were measured in the same way as in Experiment 1. Using the same exclusion criteria used in Experiment 1, approximately 7.5% of the total trials were excluded from the analyses. Mean correct ITs, MTs, and PEs were calculated for each participant in terms of n-1 congruency (congruent vs. incongruent) and current congruency (congruent vs. incongruent; see Table 2). The mean correct ITs, MTs, and PEs were analyzed by three-way repeated-measures ANOVAs, with the above factors as within-subject variables and stimulus dimension (same vs. different) as a between-subjects variable.
IT. The mean IT was significantly greater when the two tasks shared the same stimulus dimension (M = 466 ms) than when they had different stimulus dimensions (M = 409 ms), F(1, 30) = 6.76, p = 0.0143, MSE = 15,750, η2p = 0.18. The main effect of n-1 congruency was also significant, F(1, 30) = 50.78, p < 0.001, MSE = 23, η2p = 0.63, as the mean IT was greater after incongruent trial (M = 440 ms) than after congruent trials (M = 434 ms). Moreover, this post-conflict slowing interacted with stimulus dimension, F(1, 30) = 8.48, p = 0.0067, MSE = 23, η2p = 0.22, as it was larger in the same stimulus dimension (9 ms), F(1, 15) = 44.79, p < 0.001, MSE = 26, η2p = 0.75, than in different stimulus dimensions (4 ms), F(1, 15) = 10.14, p = 0.0062, MSE = 20, η2p = 0.4.
A typical Simon effect was found, F(1, 30) = 80.88, p < 0.001, MSE = 239, η2p = 0.73. The mean IT was longer on incongruent trials (M = 450 ms) than congruent trials (M = 425 ms). It further interacted with n-1 congruency, F(1, 30) = 12.41, p = 0.0014, MSE = 26, η2p = 0.29, as the size of the Simon effect was larger when preceded by congruent trials, (28 ms), F(1, 30) = 73.35, p < 0.001, MSE = 168, η2p = 0.71, than incongruent trials, (21 ms), F(1, 30) = 75.39, p < 0.001, MSE = 97, η2p = 0.72. However, because incongruent trials, as well as congruent trials, showed longer IT after incongruent trials than after congruent trials, it did not correspond to the typical pattern of the CSE that features faster responses on iI trials than cI trials (Erb & Marcovitch, 2018; Gratton et al., 1992). The three-way interaction of current congruency, n-1 congruency, and stimulus dimension was not significant, F(1, 30) = 0.18, p = 0.67, MSE = 26, η2p = 0.006 (see Fig. 5).

Mean ITs (upper) and MTs (below) as a function of n-1 congruency and current congruency (dashed vs. solid lines) for the same task-relevant stimulus dimension (left) and the different task-relevant stimulus dimension (right) in Experiment 2
MT. A main effect of n-1 congruency was significant, F(1, 30) = 5.05, p = 0.0322, MSE = 31, η2p = 0.14, as the mean MT was greater after congruent trials (M = 133 ms) than after incongruent trials (M = 131 ms). The MT data showed a significant Simon effect, F(1, 30) = 58.66, p < 0.001, MSE = 153, η2p = 0.66, as MT was greater on incongruent trials (M = 140 ms) than on congruent trials (M = 123 ms). The interaction between current congruency and n-1 congruency was also significant, F(1, 30) = 12.59, p = 0.0013, MSE = 23, η2p = 0.3, indicating a CSE. The magnitude of the Simon effect was smaller after incongruent trials (14 ms), F(1, 30) = 43.04, p < 0.001, MSE = 70, η2p = 0.59, than after congruent trials (20 ms), F(1, 30) = 58.96, p < 0.001, MSE = 106, η2p = 0.66. Other main effects or interaction effects were not significant, Fs(1, 30) < 0.89, ps > 0.35, including the three-way interaction of current congruency, n-1 congruency, and stimulus dimension, F(1, 30) = 0.61, p = 0.44, MSE = 23, η2p = 0.02 (see Fig. 5).
PE. The overall PE was 1.73%. There was a significant main effect of n-1 congruency, F(1, 30) = 6.02, p = 0.0201, MSE = 1.4, η2p = 0.17, as participants made more errors after congruent trials (1.99%) than after incongruent trials (1.48%). A typical Simon effect was also found (1.54%), F(1, 30) = 16.29, p < 0.001, MSE = 4.63, η2p = 0.35, and it interacted with n-1 congruency, F(1, 30) = 10.65, p = 0.0027, MSE = 1.84, η2p = 0.26. A significant Simon effect was observed following congruent trials (2.32%), F(1, 30) = 21.49, p < 0.001, MSE = 4.01, η2p = 0.42, but not following incongruent trials (0.75%), F(1, 30) = 3.67, p = 0.07, MSE = 2.47, η2p = 0.11. The three-way interaction of current congruency, n-1 congruency, and stimulus dimension was not significant, F(1, 30) = 1.43, p = 0.24, MSE = 1.84, η2p = 0.05.
Distributional analysis. Delta functions were calculated in the same way as in Experiment 1 (see Fig. 6) and four-way repeated-measures ANOVAs were conducted on the quintile-averaged data with stimulus dimension (same vs. different) as a between-subject variable and task (horizontal vs. vertical Simon task), current congruency (congruent vs. incongruent), and quintile (1–5) as within-subject variables, after the Greenhouse–Geisser correction.

The magnitude of Simon effects across quintiles for the horizontal (solid lines) and vertical (dashed lines) Simon tasks in Experiment 2. ITs (black lines) and MTs (gray lines) were plotted, respectively, for each task-relevant stimulus dimension (upper vs. below)
The interaction between current congruency and quintile was significant in the IT, F(1, 43) = 41.05, p < 0.001, MSE = 620, η2p = 0.58, and MT data, F(1, 43) = 12.99, p < 0.001, MSE = 476, η2p = 0.3. However, the overall Simon effect decreased across quintiles in the IT data, but increased in the MT data. These increasing delta function found in the MT data was modulated by task, F(2, 55) = 6.66, p = 0.003, MSE = 276, η2p = 0.18, as the slope of delta function was steeper for the vertical Simon effect than for the horizontal Simon effect. A significant four-way interaction of stimulus dimension, task, current congruency, and quintile was not obtained in the MT data, F(2, 55) = 0.76, p = 0.46, MSE = 276, η2p = 0.02. Most importantly, the magnitude of the Simon effect in the IT data decreased across quintiles both for the horizontal and vertical Simon tasks, as the three-way interaction of task, current congruency, and quintile was not significant, F(3, 91) = 0.67, p = 0.57, MSE = 109, η2p = 0.02, regardless of stimulus dimension, F(3, 91) = 0.04, p = 0.99, MSE = 109, η2p = 0.0014. These results in the IT data indicate that the temporal dynamics of the horizontal and vertical Simon effects were not statistically different, regardless of whether the two tasks shared the same task-relevant stimulus dimension or not.
Discussion
When responding with the same response mode, a significant CSE was obtained between the horizontal and vertical Simon congruencies in the MT data as well as in the PE data. Moreover, the CSE found in the MT data did not vary depending on whether the two Simon tasks had the same or different task-relevant stimulus dimensions. The magnitude of the Simon effect was smaller after incongruent trials (12 ms) than after congruent trials (19 ms) when the two tasks shared the task-relevant stimulus dimension, and the same pattern of sequential modulation was obtained when they had different task-relevant stimulus dimensions (13 ms after incongruent trials, and 19 ms after congruent trials). Also, the IT data showed that the magnitude of the Simon effect decreased across quintiles both in the horizontal and vertical Simon tasks, regardless of task-relevant stimulus dimension. Those results from the distributional analyses provide further evidence that using the same response mode, the horizontal and vertical spatial stimulus codes activate their corresponding responses in a comparable manner, which allowed them to be administered by the same control mechanism. These results suggest that the same control mechanism was recruited even when the two task sets were distinguished clearly, as long as the spatial representation of the horizontal and vertical dimensions remained comparable by employing the same response mode. Furthermore, unlike spatial dimension and response mode in Experiment 1, task-relevant dimensions did not determine the boundary of control. These results together suggest that control underlying the CSE resolves Simon-type conflict by suppressing the response activated by the task-irrelevant stimulus feature (i.e., stimulus location), rather than enhancing the response activated by the task-relevant stimulus feature (Kim, Lee, & Cho, 2015; Lee & Cho, 2013; Stürmer et al., 2002).
Additionally, the overall IT was greater when the two tasks shared the same stimulus dimension (M = 466 ms) than when they had different stimulus dimensions (M = 409 ms). It is possible that maintaining four stimulus–response mappings was more difficult than maintaining two sets of two stimulus–response mappings. This result is consistent with the previous findings that categorizing individual stimulus–response mappings into different task sets depending on additional mapping rules helps to maintain only relevant information selectively, resulting in improved performance efficiency, even though it compromises the flexibility in the face of changing environment or induces a task-switching cost (Dreisbach, 2012; Lim & Cho, 2018).
Experiment 3
The findings of Experiments 1 and 2 suggested that the horizontal and vertical Simon tasks are controlled by the same mechanism when they are performed with the same response mode. By using the same mode, the horizontal and vertical locations seemed to be spatially coded in a single spatial dimension, which was corroborated by the results from the distributional analyses. However, it is possible that response mode plays a dominant role particularly in the Simon task, because it features response-based conflict between two competing responses activated by the task-relevant (e.g., color) and the task-irrelevant (i.e., location) stimulus features (Kornblum, 1994). In contrast, other congruency tasks, such as the Stroop or flanker-compatibility task, involve stimulus-based conflict which occurs between the task-relevant and task-irrelevant stimulus features. Since their conflicts do not directly arise from the response dimension, the role of response mode in those tasks may not be as critical as in the Simon task. Accordingly, Experiment 3 was conducted to investigate whether response mode reconfigures spatial representations to modulate the boundary of control, even when conflict arises not directly from response dimensions.
For this purpose, the horizontal and vertical versions of the flanker-compatibility tasks were presented in turn, in which participants were to respond to the arrow direction of the target. Response mode was manipulated as in Experiment 1 (see Fig. 7). However, unlike Experiments 1 and 2, the stimulus–response (S–R) mappings of the horizontal and vertical arrow flanker-compatibility tasks were related to each other in a non-arbitrary manner, because the movement direction of responses always corresponded to the spatial features of the target. Interestingly, when two tasks having the same stimulus dimension also share this kind of non-arbitrary S–R mapping rule, such as alphabetical order, numerical order, or directional correspondence in case of the present situation, significant cross-task CSEs have been observed regardless of response mode (Lim & Cho, 2018; Weissman, Colter, Drake, & Morgan, 2015). Lim and Cho suggested that a common logical rule encompassing two tasks’ S–R mappings makes the distinction between different response modes less salient, allowing the two tasks to be controlled by the same mechanism. Thus, if response mode cannot reconfigure the spatial representations of the horizontal and vertical dimensions in case the conflict does not originate from response dimensions, the CSE should be evident regardless of response mode due to the non-arbitrary S–R mappings. Otherwise, if response mode reconfigures the spatial dimensions due to intrinsic associations between the spatial codes of stimulus and response (Hommel, 2011; Zhang, Zhang, & Kornblum, 1999), the CSE should be obtained only when the horizontal and vertical dimensions are represented as a single spatial dimension by employing the same mode. However, no CSE would occur when the horizontal and vertical dimensions are separately represented by employing different modes, because in this case, the two tasks would not share any task features including the task-relevant or task-irrelevant stimulus dimensions (Egner & Hirsch, 2005; Verguts & Notebaert, 2008, 2009).

Illustrations of the stimulus and response sets used in Experiment 3. In the horizontal and vertical flanker-compatibility tasks, horizontal or vertical arrow target and flanker stimuli were presented, respectively. The horizontal and vertical responses, denoted by black and gray arrows respectively, were carried out with the same or different response modes
Method
Participants. Thirty-two new participants (9 males, mean age = 23) from the same pool as the previous experiments took part. Sixteen participants performed the horizontal and vertical arrow flanker-compatibility tasks with the same mode, while the other 16 participants performed those tasks with different modes.
Stimulus and apparatus. The apparatus and stimuli were the same as in Experiment 1 except for the following differences. Left and/or right arrows (approximately 0.36° × 0.72°) appeared as a target and flankers in the horizontal arrow flanker-compatibility task and up and/or down arrows (approximately 0.72° × 0.36°) were presented in the vertical arrow flanker-compatibility task. Participants were to press the ‘left’, ‘right’, ‘up’, and ‘down’ direction keys to left-, right-, up-, and down-pointing target arrows, respectively. The target was presented at the center of the display with two flankers on both left and right sides for the horizontal task and on both above and below for the vertical task. The target and flanker stimuli were equally spaced about 0.3° apart. The same direction keys and home keys were used as in Experiment 1.
Procedure. All procedures were identical to those in Experiment 1 with the following exceptions. The horizontal and vertical arrow flanker-compatibility tasks alternated in a trial-by-trial manner. Participants were to respond to the direction of the target arrow while ignoring the flanker arrows (see Fig. 7).
Results
Using the same exclusion criteria as in the previous experiments, approximately 6.04% of the total trials were removed from the analyses. Individual participant’s mean correct ITs, MTs, and PEs were calculated as a function of n-1 congruency (congruent vs. incongruent) and current congruency (congruent vs. incongruent; see Table 3). Three-way repeated-measures ANOVAs were conducted on the mean correct ITs, MTs, and PEs with the above factors as within-subject variables and response mode (same vs. different) as a between-subject variable.
IT. Participants responded more slowly after incongruent trials (M = 402 ms) than after congruent trials (M = 398 ms), F(1, 30) = 28.04, p < 0.001, MSE = 23, η2p = 0.48, indicating post-conflict slowing. A significant main effect of response mode was also observed, F(1, 30) = 12.84, p = 0.0012, MSE = 10,883, η2p = 0.2, as the mean IT was greater for different modes (M = 433 ms) than for the same mode (M = 367 ms). A flanker-compatibility effect was significant, F(1, 30) = 128.42, p < 0.001, MSE = 488, η2p = 0.81. It took longer to respond to incongruent trials (M = 422 ms) than congruent trials (M = 378 ms). The interaction between congruency and response mode was significant, F(1, 30) = 11.94, p = 0.0017, MSE = 488, η2p = 0.28. The flanker-compatibility effect was larger with different modes (58 ms), F(1, 15) = 80.9, p < 0.001, MSE = 660, η2p = 0.84, than with the same mode (31 ms), F(1, 15) = 47.9, p < 0.001, MSE = 316, η2p = 0.76 (see Fig. 8).

Mean ITs (upper) and MTs (below) as a function of n-1 congruency and current congruency (dashed vs. solid lines) for the same mode (left) and different modes (right) in Experiment 3
MT. The mean MT was significantly greater after congruent trials (M = 124 ms) than after incongruent trials (M = 121 ms), F(1, 30) = 25.42, p < 0.001, MSE = 17, η2p = 0.46, indicating post-conflict slowing. A significant flanker-compatibility effect was found, F(1, 30) = 60.2, p < 0.001, MSE = 331, η2p = 0.67. The mean MT was greater on incongruent trials (M = 135 ms) than congruent trials (M = 110 ms). The interaction between congruency and response mode was also obtained, F(1, 30) = 9.79, p = 0.0039, MSE = 331, η2p = 0.25, as the flanker-compatibility effect was larger with the same mode (35 ms), F(1, 15) = 47.96, p < 0.001, MSE = 409, η2p = 0.76, than with different modes (15 ms), F(1, 15) = 14.03, p = 0.002, MSE = 253, η2p = 0.48.
Furthermore, the flanker-compatibility effect was modulated by n-1 congruency, F(1, 30) = 10.42, p = 0.003, MSE = 21, η2p = 0.26, indicating a CSE. The magnitude of the flanker-compatibility effect was smaller after incongruent trials (22 ms), F(1, 30) = 56.96, p < 0.001, MSE = 140, η2p = 0.66, than after congruent trials (28 ms), F(1, 30) = 57.43, p < 0.001, MSE = 212, η2p = 0.66. Importantly, the three-way interaction of current congruency, n-1 congruency, and response mode was evident, F(1, 30) = 4.2, p = 0.0493, MSE = 21, η2p = 0.12. Separate analyses on each response mode revealed that the CSE was significant with the same mode, F(1, 15) = 18.35, p < 0.001, MSE = 16, η2p = 0.55, as the size of the flanker-compatibility effect became smaller after incongruent trials (31 ms), F(1, 15) = 39.45, p < 0.001, MSE = 192, η2p = 0.72, than after congruent trials (39 ms), F(1, 15) = 52.95, p < 0.001, MSE = 233, η2p = 0.78. However, no CSE was found with different modes, F(1, 15) = 0.56, p = 0.47, MSE = 26, η2p = 0.04 (see Fig. 8).
PE. The overall PE was 0.83%. A significant 1.23% flanker-compatibility effect was observed, F(1, 30) = 23.13, p < 0.001, MSE = 2.09, η2p = 0.44, indicating a flanker-compatibility effect of 1.23%. The interaction of current congruency with response mode was significant, F(1, 30) = 6.03, p = 0.0201, MSE = 2.09, η2p = 0.17, as the size of the flanker-compatibility effect was smaller with different modes (0.6%), F(1, 15) = 6.97, p = 0.0186, MSE = 0.83, η2p = 0.32, than with the same mode (1.86%), F(1, 15) = 16.47, p = 0.001, MSE = 3.35, η2p = 0.52. No other effects reached significance, Fs(1, 30) < 1.96, ps > 0.17, including the interaction of current congruency with n-1 congruency, F(1, 30) = 0.13, p = 0.72, MSE = 0.59, η2p = 0.0043, and the three-way interaction of current congruency, n-1 congruency, and response mode, F(1, 30) = 1.15, p = 0.29, MSE = 0.59, η2p = 0.04.
Distributional analysis. Although the flanker-compatibility effect has not shown the typical decreasing pattern of the delta function, unlike the Simon effect, distributional analyses were conducted for the sake of completeness and to be briefly discussed. Delta functions were calculated in the same way as in Experiments 1 and 2 and four-way repeated-measures ANOVAs were conducted on the quintile-averaged data with response mode (same vs. different) as a between-subject variable and task (horizontal vs. vertical Simon task), current congruency (congruent vs. incongruent) and quintile (1–5) as within-subject variables, after the Greenhouse–Geisser correction.
The interaction between current congruency and quintile was significant in the MT data, F(2, 50) = 14.99, p < 0.001, MSE = 433, η2p = 0.33, as the overall flanker-compatibility effects increased across quintiles (see also, Wylie et al., 2009). Also, the MT data showed a significant interaction of task, current congruency, and quintile, F(2, 72) = 6.24, p = 0.002, MSE = 121, η2p = 0.17, as the slope of the delta function was steeper for the horizontal flanker-compatibility effect than for the vertical flanker-compatibility task. No other interaction effects related to quintile and current congruency reached significance, Fs < 1.64, ps > 0.2.
Discussion
Response mode modulated the boundary of control by reconfiguring the spatial dimensions even when the conflict did not arise from the response dimension. The cross-task CSEs were significant in the MT data, and furthermore, both varied depending on response mode. As in Experiment 1, the CSEs were significant when the same mode was employed to perform the tasks. However, unlike previous studies (Lim & Cho, 2018; Weissman et al., 2015), no CSE was obtained when different modes were employed, in spite of the non-arbitrary S-R mappings. These results indicate that spatial dimensions are different from other non-arbitrary but non-spatial dimensions in that they are malleable to form separate representations based on the axis distinction (Adam et al., 2003; Ansorge & Wühr, 2004; Vu et al., 2005, 2000). Since different modes made the distinction between the horizontal and vertical dimensions salient to reconfigure them as different dimensions, the horizontal and vertical arrow flanker-compatibility tasks did not share either the task-relevant or task-irrelevant stimulus dimension, resulting in no cross-task CSE (Egner & Hirsch, 2005; Verguts & Notebaert, 2008, 2009). These results are consistent with the idea that response mode determines the scope of control by affecting how spatial dimensions are cognitively represented.
General discussion
The present study investigated how response mode determines the specificity of control especially when conflict stems from the horizontal and vertical spatial dimensions. In Experiment 1, the CSE between the horizontal and vertical Simon tasks was significant in the MT data, but only when their response sets were represented as a single response mode. In Experiment 2, by sharing the same mode, the CSE was evident in the MT data, regardless of whether the two tasks had the same or different task-relevant stimulus dimensions. These results suggest that the scope of control depends on response mode, as it reconfigures the spatial coding of the horizontal and vertical dimensions into either a single or two separate dimensions. The distributional analyses provided further corroboration as the magnitude of the vertical Simon effect decreased across quintiles, similar to that of the horizontal Simon effect, only when the two tasks were responded to with the same mode. This conclusion was generalizable to the tasks of which conflict is not derived directly from the response dimensions. When the horizontal and vertical arrow flanker-compatibility tasks were employed in Experiment 3, the CSE was observed between the tasks when they were carried out with the same mode but not with different modes, consistent with the results from Experiment 1.
Moreover, the scope of control was determined independently of the task-relevant stimulus dimension (Experiment 2). This indicates that response mode determined the scope of control by reconfiguring the spatial representation of the task-irrelevant stimulus dimension, even when the distinction between the two task sets was clear. Furthermore, taken together with the results of Experiment 1, it seems that the control mechanism inhibits the response activated by the task-irrelevant stimulus dimension rather than facilitates the processing of the task-relevant stimulus dimension to resolve, at least, Simon-type conflict. Consistent with this idea, the previous findings also suggested that sharing a common spatial dimension and response mode, not the task-relevant stimulus dimension, is necessary for recruiting the same control mechanism to resolve, at least, the Simon-type conflict (Kim et al., 2015; Lee & Cho, 2013; Stürmer et al., 2002). For example, when the same response mode was employed, the CSE was significant between two horizontal Simon tasks having different task-relevant stimulus dimensions (Kim et al., 2015), whereas no cross-task CSE was obtained between horizontal and orthogonal Simon tasks, which had the same task-relevant stimulus dimension but different task-irrelevant stimulus dimensions (Lee & Cho, 2013).
Contrary to the current findings, Notebaert and Verguts (2008) found no CSE between Simon and SNARC tasks when they had different task-relevant stimulus dimensions. However, in their experiment, because the two tasks employed exactly the same response alternatives, the two task sets could not be actively maintained at the same time. It has been suggested that in order for the control triggered by one task to be transferred to another task, the two task sets (including both task-relevant and task-irrelevant features) should be highly similar or dissimilar enough to be simultaneously maintained in working memory without interference (Braem et al., 2014), because only active associations can be reinforced by a control mechanism (Verguts & Notebaert, 2008, 2009). Thus, if two tasks employ the same response alternatives, but have different task-relevant dimensions (i.e., partially similar), as in Notebaert and Verguts’ experiment, the cross-task CSE cannot occur due to the mutual interference between the two tasks. However, in the present study, all stimulus features (both task-relevant and task-irrelevant) were assigned to different response alternatives, preventing any interference between the task sets.
In addition, as the present study is the first to observe the confound-minimized CSE in MT, a mini meta-analysis was conducted to examine whether response mode reliably modulates the cross-task CSE in MT across Experiments 1 and 3 (Goh, Hall, & Rosenthal, 2016). Experiments 1 and 3 were meta-analyzed using a fixed-effects approach. The interaction of current congruency, n-1 congruency, and response mode was reliably found to be significant in MT, d = 9.19, 95% CI = [7.51, 10.86], Z = 10.76, p < 0.001.
How response mode determines the specificity of control
Regarding how response mode modulates the specificity of control, the current findings suggest that response mode reconfigures the spatial representations of the horizontal and vertical dimensions. The stimulus and response alternatives can involve several spatial codes based on multiple frames of references, which are not equally coded but differently weighted according to the task environments, such as stimulus arrangements, response configurations, and response-discriminating feature (e.g., Adam et al., 2003; Ansorge & Wühr, 2004; Proctor et al., 2003; Vu et al., 2000). For example, Ansorge and Wühr (2004) found that the Simon effect was evident only for the response-discriminating dimension, when response alternatives were discriminated in terms of either horizontal or vertical dimensions. Their results indicate that even though the stimulus location itself varied along both the horizontal and vertical dimensions, spatial codes were activated strongly enough to engender the Simon effect only for the dimension that discriminated response alternatives. Hommel (2011) even suggested that the representations of stimuli and responses share the same feature codes, so the features that define or discriminate the correct response influence how a stimulus is cognitively coded. Therefore, when the representations of the horizontal and vertical Simon tasks’ response sets are grouped into a same response mode, it is possible that the spatial distinction between the horizontal and vertical dimensions became less salient or redundant, and thus, they are likely to be spatially coded as a single dimension.
This idea is in accordance with the distributional analyses in the present study, as the decreasing delta functions were consistently observed in the IT data for the vertical Simon tasks, as well as for the horizontal Simon tasks, only when they were performed with the same response mode. An ample amount of studies suggested that the decreasing delta function is particularly found in horizontal Simon tasks, and it reflects the temporal dynamics of the response activation from the stimulus location (for a review, see Proctor et al., 2011). However, unlike the current findings, the vertical Simon tasks predominantly show constant or increasing delta functions (Proctor et al., 2003; Wiegand & Wascher, 2005). Moreover, in Buetti and Kerzel’s (2008) experiment, only the horizontal but not the vertical Simon effect decreased across quintiles even with aimed-movement responses, when the two tasks were performed on two separate days. Exactly what mechanism underlies the delta functions and what causes the discrepancies between the horizontal and vertical Simon effects are still controversial (e.g., Miller & Roüast, 2016; Proctor et al., 2011; Zhang & Kornblum, 1997). However, at least, the decreasing delta function of the vertical Simon effect observed in the current findings implies that the responses from the vertical codes were activated and controlled in a comparable manner as those from the horizontal codes (Töbel, Hübner, & Stürmer, 2014; Wiegand & Wascher, 2007).
Note that response mode is not merely determined by an anatomical distinction. Some studies obtained significant sequential modulation between the horizontal and vertical Simon tasks, even when their response sets were discriminated in terms of hands (Weissman et al., 2014), which seems to be contradictory to the current findings. However, although the anatomical distinction is certainly one of the salient features that distinguishes response modes, response mode is more flexibly configured depending on other relevant features, as well (Adam, 1994; Adam et al., 2003; Ansorge & Wühr, 2004; Freedberg et al., 2014; Hazeltine, 2005; Lippa, 1996; Proctor & Reeve, 1985, 1986; Reeve & Proctor, 1984). Therefore, a closer examination is required to evaluate whether the response sets distinguished by different effectors are indeed represented as different modes. For instance, in Weissman et al.’s (2014) study, the sequential modulation occurred between the horizontal and vertical Simon tasks performed with the index and middle fingers of the left and right hands, respectively. However, to press the neighboring response keys, the left and right hands must be spatially overlapped to a great extent. In this case, various frames of reference are involved in spatial coding of each response alternative, and among them, the distinction between the left- and right-hand responses is unlikely to be salient enough to form two separate response modes.
How control mechanisms modulate the activation of competing responses
By measuring IT and MT separately, the aimed-movement responses provide further insight into how the control mechanism modulates competing response activations, from an abstract level of response selection to motor programming processes (Doucet & Stelmack, 1999; Hietanen & Pia, 1995; Rubichi & Pellicano, 2004) with relatively simple and clear experimental designs, compared to those devised for standard keypress responses. The CSEs consistently found in the MT data of the present study provide evidence that the goal-directed movement is constantly controlled even after the movement was initiated. Recent studies suggested that the response selection and motor execution are not discrete or serial processes (Buetti & Kerzel, 2008, 2009; Calderon, Gevers, & Verguts, 2018; Erlhagen & Schöner, 2002; Hommel, 2009; Resulaj, Kiani, Wolpert, & Shadlen, 2009). When a particular response activation exceeds other response activations and reaches a certain threshold, the movement is likely to be initiated, although the selection is not fully completed. Such partial completion means that response selection is not a process that exclusively leaves one response activation and discards all the others entirely. The influences of those unselected response activations can linger on the post-response selection stages.
Therefore, motor control should operate to keep inhibiting the task-irrelevant response activation from biasing the response towards an incorrect direction even after the movement is initiated. In other words, motor execution is not a process that merely transforms the selected response activation into a physical action. It also involves motor control that programs movement parameters, such as spatial coordination of movement, velocity, movement trajectory, or grasping force (e.g., Buetti & Kerzel, 2009; Freud, Aisenberg, Salzer, Henik, & Ganel, 2015). During this motor programming stage, motor control assigns values to movement parameters, constantly tunes, and updates those values depending on the task environments even after the movement initiation (Calderon et al., 2018; Erlhagen & Schöner, 2002). From these perspectives, it is possible that IT mainly reflects how the strengths of the abstract response activations (e.g., left, right, above, or below) are gradually modulated over time, of which temporal dynamics are illustrated in the distributional analyses. In contrast, MT may reflect how specific movement parameters are programmed after the difference between multiple response activations reaches the thresholds to initiate the action (Buetti & Kerzel, 2009).
Indeed, the present study found significant Simon and flanker-compatibility effects in the MT data as well as in the IT data, which is in accordance with other studies that found significant Simon effects in movement parameters such as the curvature of movement trajectories (Buetti & Kerzel, 2008, 2009; Scherbaum, Dshemuchadse, Fischer, & Goschke, 2010), the maximum height of the hand (Kerzel & Buetti, 2012), grasping trajectories (Freud et al., 2015), or keypress force (Miller & Roüast, 2016). These results indicate that the response activated by the task-irrelevant spatial information keeps influencing the motor programming of movement parameters even after the movement initiation (Buetti & Kerzel, 2009). However, the slope of the delta function, which reflects the temporal dynamic of the two competing response activations, decreased in the IT data but not in the MT data. Rather, the MT data showed constant or slightly increasing delta functions. Considering that the movement is supposed to be initiated after the activation level of a certain response exceeds the other competing ones, it is possible that once the movement is initiated, the movement parameters become the main target of control, instead of the response activation proper (Scherbaum et al., 2010). Thus, the delta function, which reflects the temporal dynamics of response activations, does not necessarily decrease in MT anymore.
Most interestingly, the present study demonstrates unique findings as it observed the CSE clearly in the MT data, rather than in the IT data. This suggests an intriguing possibility that the control mechanism underlying the CSE regulates motor programming processes after the movement is initiated. One can raise the possibility that the motivational effects caused by the release of a key (i.e., IT) and the press of a key (i.e., MT) might have different influences on the congruency effect, resulting in the absence of the CSE in the IT data. Since the avoidance motivation tendencies caused by release force (Hochman, Milman, & Tal, 2017) can lead to faster responses on incongruent trials (e.g., Dignath & Eder, 2015), the congruency effect, and thus, the CSE might be reduced in IT. However, significant magnitudes of congruency effects to engender the CSE were consistently observed across three experiments (20 ms, 25 ms, 44 ms, respectively). In addition, it has been suggested that the magnitude of the CSE is not correlated with that of the congruency effect (Weissman et al., 2014). Thus, it is unlikely that the absence of the CSE in the IT data was entirely due to the avoidance motivation inherent in key-release responses.
Consistent with the current findings, some recent studies also found the sequential modulation in various kinds of movement parameters (e.g., Dignath et al., 2019; Erb & Marcovitch, 2018; Erb, Moher, Sobel, & Song, 2016; Miller & Roüast, 2016; Scherbaum et al, 2010; Weissman, 2019a, b). It is worth noting that these movement parameters provide enriched information about how competing response activations are gradually modulated over time, which enables researchers to have a closer look at the dynamics of response selection and motor programming processes. For example, Scherbaum et al. (2010) found the sequential modulation of the Simon effect in the curvature of mouse trajectories. They suggested that control underlying the CSE enhances the reactivation of the conflict resolution process and promotes the final execution of responses, which is consistent with the present findings that the CSE was evident after an abstract response was selected.
Similarly, some studies measured the trajectories of hand movement reaching toward the target and observed significant CSEs in motoric measures, like MT and reach curvature, but not in IT (e.g., Erb & Marcovitch, 2018; Erb et al., 2016). Even though Dignath and his colleges (2020) found significant sequential modulations triggered by the motivational mechanism both in IT and reach curvature, their patterns were different. They suggested that the sequential modulation in IT came from the facilitation of target processing after experiencing conflict, while that in reach curvature was caused by the suppression of distractor processing. In line with the current findings that the confound-minimized CSE was evident in MT data but not IT data, these results imply that the control operates in different manners before and after movement is initiated, even though the distinction between response selection and motor execution might be not necessarily clear.
Miller and Roüast (2016) measured another movement parameter, the response force (RF), while participants performed a horizontal Simon task. RF denotes the forcefulness of keypress, which reflects the response activation at a motor level, like other measures such as LRP or EMG (e.g., Burle, Possamaï, Vidal, Bonnet, & Hasbroucq, 2002; Stürmer et al., 2002). Especially, in the Simon task, RF generated by the non-responding effector reflects the sub-threshold response activation from the stimulus location. They found not only a significant Simon effect, but also the CSE in RF of the non-responding finger. In line with the current findings in Experiment 2, this result suggests that the top-down control mechanism underlying the CSE modulates the programming of movement parameters, especially those for the responses corresponding to the task-irrelevant stimulus location.
Conclusion
The present findings suggest that the scope of control responsible for the CSE depends on response mode. Although some found the cross-task CSE independently of response mode when the prime-probe task was used (Weissman et al., 2014, 2015), the present study consistently found that the scope of control depended on response mode, at least when spatial conflict arose in Simon and flanker-compatibility tasks. It suggests that response mode determines the scope of control triggered by spatial conflict as it reconfigures the spatial representation of the horizontal and vertical dimensions. This account explains why existing studies, which involve different response configurations, have shown inconsistent results. Unlike previous studies, the current study successfully examined the effect of response mode on the CSE while keeping the relative salience of the horizontal and vertical dimensions equivalent using aimed-movement responses. Furthermore, it provides new insight on the nature of control that gradually resolves conflict, from the response selection to motor programming processes, and its temporal dynamics with a relatively simple and affordable method, compared to previous studies relying on standard keypress responses. To our knowledge, the present study is the first to consistently observe the confound-minimized CSE in MT data, which implies that the response bias towards the task-irrelevant spatial dimension is controlled during the motor programming process, even after an abstract response is selected.
Notes
To prevent participants from explicitly selecting the correct response after releasing home key (Rubichi & Pellicano, 2004; Smith & Carew, 1987), the current study minimized the possibility of delayed response selection in the following ways. First, participants were explicitly instructed to release the home key after they decided which direction key to press. Also, the target disappeared from the screen as soon as the home key was released, which minimized the possibility that participants decided the response key after initiating movement (Smith & Carew, 1987). Finally, only those trials, of which both IT and MT were not too fast or slow from its conditional mean, were analyzed. In this way, the trials where the faster IT was traded for slower MT could be excluded from the analyses.
References
Adam, J. J. (1994). Manipulating the spatial arrangement of stimuli in a precuing task. Acta Psychologica, 85(3), 183–202.
Adam, J. J., Hommel, B., & Umiltà, C. (2003). Preparing for perception and action (I): The role of grouping in the response-cuing paradigm. Cognitive Psychology, 46(3), 302–358.
Akçay, Ç., & Hazeltine, E. (2008). Conflict adaptation depends on task structure. Journal of Experimental Psychology: Human Perception and Performance, 34(4), 958–973.
Ansorge, U., & Wühr, P. (2004). A response-discrimination account of the simon effect. Journal of Experimental Psychology: Human Perception and Performance, 30(2), 365–377.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624–652.
Braem, S., Abrahamse, E. L., Duthoo, W., & Notebaert, W. (2014). What determines the specificity of conflict adaptation? A review, critical analysis, and proposed synthesis. Frontiers in Psychology, 5, 1134.
Braem, S., Bugg, J. M., Schmidt, J. R., Crump, M. J., Weissman, D. H., Notebaert, W., et al. (2019). Measuring adaptive control in conflict tasks. Trends in Cognitive Sciences, 23(9), 769–783.
Braem, S., Verguts, T., & Notebaert, W. (2011). Conflict adaptation by means of associative learning. Journal of Experimental Psychology: Human Perception and Performance, 37(5), 1662–1666.
Buetti, S., & Kerzel, D. (2008). Time course of the Simon effect in pointing movements for horizontal, vertical, and acoustic stimuli: Evidence for a common mechanism. Acta Psychologica, 129(3), 420–428.
Buetti, S., & Kerzel, D. (2009). Conflicts during response selection affect response programming: Reactions toward the source of stimulation. Journal of Experimental Psychology: Human Perception and Performance, 35(3), 816–834.
Burle, B., Possamaï, C. A., Vidal, F., Bonnet, M., & Hasbroucq, T. (2002). Executive control in the Simon effect: An electromyographic and distributional analysis. Psychological Research Psychologische Forschung, 66(4), 324–336.
Calderon, C. B., Gevers, W., & Verguts, T. (2018). The unfolding action model of initiation times, movement times, and movement paths. Psychological Review, 125(5), 785–805.
De Jong, R., Liang, C.-C., & Lauber, E. (1994). Conditional and unconditional automaticity: A dual-process model of effects of spatial stimulus-response correspondence. Journal of Experimental Psychology: Human Perception and Performance, 20(4), 731–750.
Dignath, D., & Eder, A. B. (2015). Stimulus conflict triggers behavioral avoidance. Cognitive, Affective, & Behavioral Neuroscience, 15(4), 822–836.
Dignath, D., Wirth, R., Kühnhausen, J., Gawrilow, C., Kunde, W., & Kiesel, A. (2019). Motivation drives conflict adaptation. Motivation Science, 6(1), 84–89.
Doucet, C., & Stelmack, R. M. (1999). The effect of response execution on P3 latency, reaction time, and movement time. Psychophysiology, 36(3), 351–363.
Dreisbach, G. (2012). Mechanisms of cognitive control: The functional role of task rules. Current Directions in Psychological Science, 21(4), 227–231.
Egner, T., & Hirsch, J. (2005). Cognitive control mechanisms resolve conflict through cortical amplification of task-relevant information. Nature Neuroscience, 8(12), 1784–1790.
Erb, C. D., & Marcovitch, S. (2018). Deconstructing the Gratton effect: Targeting dissociable trial sequence effects in children, pre-adolescents, and adults. Cognition, 179, 150–162.
Erb, C. D., Moher, J., Sobel, D. M., & Song, J. H. (2016). Reach tracking reveals dissociable processes underlying cognitive control. Cognition, 152, 114–126.
Erlhagen, W., & Schöner, G. (2002). Dynamic field theory of movement preparation. Psychological Review, 109(3), 545–572.
Faul, F., Erdfelder, E., Buchner, A., & Lang, A. G. (2009). Statistical power analyses using G* Power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41(4), 1149–1160.
Freedberg, M., Wagschal, T. T., & Hazeltine, E. (2014). Incidental learning and task boundaries. Journal of Experimental Psychology: Learning, Memory, and Cognition, 40(6), 1680–1700.
Freud, E., Aisenberg, D., Salzer, Y., Henik, A., & Ganel, T. (2015). Simon in action: The effect of spatial congruency on grasping trajectories. Psychological Research Psychologische Forschung, 79(1), 134–142.
Goh, J. X., Hall, J. A., & Rosenthal, R. (2016). Mini meta-analysis of your own studies: Some arguments on why and a primer on how. Social and Personality Psychology Compass, 10(10), 535–549.
Gratton, G., Coles, M. G., & Donchin, E. (1992). Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology: General, 121(4), 480–506.
Hazeltine, E. (2005). Response-response compatibility during bimanual movements: Evidence for the conceptual coding of action. Psychonomic Bulletin & Review, 12(4), 682–688.
Hazeltine, E., Lightman, E., Schwarb, H., & Schumacher, E. H. (2011). The boundaries of sequential modulations: Evidence for set-level control. Journal of Experimental Psychology: Human Perception and Performance, 37(6), 1898–1914.
Hietanen, J. K., & Pia, R. (1995). Facilitation and interference occur at different stages of processing in the Simon paradigm. European Journal of Cognitive Psychology, 7(2), 183–199.
Hochman, E. Y., Milman, V., & Tal, L. (2017). Evidence for aversive withdrawal response to own errors. Acta Psychologica, 180, 147–154.
Hommel, B. (1996). No prevalence of right-left over top-bottom spatial codes. Perception & Psychophysics, 58(1), 102–110.
Hommel, B. (1997). Interactions between stimulus-stimulus congruence and stimulus-response compatibility. Psychological Research Psychologische Forschung, 59(4), 248–260.
Hommel, B. (2009). Action control according to TEC (theory of event coding). Psychological Research Psychologische Forschung, 73(4), 512–526.
Hommel, B. (2011). The Simon effect as tool and heuristic. Acta Psychologica, 136(2), 189–202.
Hommel, B., Proctor, R. W., & Vu, K. P. L. (2004). A feature-integration account of sequential effects in the Simon task. Psychological Research Psychologische Forschung, 68(1), 1–17.
Hoppe, K., Küper, K., & Wascher, E. (2017). Sequential modulations in a combined horizontal and vertical Simon task: Is there ERP evidence for feature integration effects? Frontiers in Psychology, 8, 1094.
Janczyk, M., & Leuthold, H. (2018). Effector system-specific sequential modulations of congruency effects. Psychonomic Bulletin & Review, 25(3), 1066–1072.
Kerzel, D., & Buetti, S. (2012). Approach and avoidance movements are unaffected by cognitive conflict: A comparison of the Simon effect and stimulus–response compatibility. Psychonomic Bulletin & Review, 19(3), 456–461.
Kim, S., & Cho, Y. S. (2014). CSE without feature integration and contingency learning. Acta Psychologica, 149(1), 60–68.
Kim, S., Lee, S. H., & Cho, Y. S. (2015). Control processes through the suppression of the automatic response activation triggered by task-irrelevant information in the Simon-type tasks. Acta Psychologica, 162(1), 51–61.
Kornblum, S. (1994). The way irrelevant dimensions are processed depends on what they overlap with: The case of Stroop-and Simon-like stimuli. Psychological Research Psychologische Forschung, 56(3), 130–135.
Lee, J., & Cho, Y. S. (2013). CSE in cross-task context: Evidence for dimension-specific modulation. Acta Psychologica, 144(3), 617–627.
Lim, C. E., & Cho, Y. S. (2018). Determining the scope of control underlying the congruency sequence effect: Roles of stimulus-response mapping and response mode. Acta Psychologica, 190, 267–276.
Lippa, Y. (1996). A referential coding explanation for compatibility effects of physically orthogonal stimulus and response dimensions. The Quarterly Journal of Experimental Psychology Section A, 49(4), 950–971.
Loftus, G. R., & Masson, M. E. (1994). Using confidence intervals in within-subject designs. Psychonomic Bulletin & Review, 1(4), 476–490.
Mayr, U., Awh, E., & Laurey, P. (2003). Conflict adaptation effects in the absence of executive control. Nature Neuroscience, 6(5), 450–452.
Miller, J. (1982). Discrete versus continuous stage models of human information processing: In search of partial output. Journal of Experimental Psychology: Human Perception and Performance, 8(2), 273.
Miller, J., & Roüast, N. M. (2016). Dissociations of spatial congruence effects across response measures: An examination of delta plots. Psychological Research Psychologische Forschung, 80(5), 805–820.
Notebaert, W., & Verguts, T. (2008). Cognitive control acts locally. Cognition, 106(2), 1071–1080.
Proctor, R. W., & Reeve, T. G. (1985). Compatibility effects in the assignment of symbolic stimuli to discrete finger responses. Journal of Experimental Psychology: Human Perception and Performance, 11(5), 623–639.
Proctor, R. W., & Reeve, T. G. (1986). Salient-feature coding operations in spatial precuing tasks. Journal of Experimental Psychology: Human Perception and Performance, 12(3), 277.
Proctor, R. W., & Vu, K.-P. L. (2010). Stimulus–response compatibility for mixed mappings and tasks with unique responses. The Quarterly Journal of Experimental Psychology, 63(2), 320–340.
Proctor, R. W., Miles, J. D., & Baroni, G. (2011). Reaction time distribution analysis of spatial correspondence effects. Psychonomic Bulletin & Review, 18(2), 242–266.
Proctor, R. W., Vu, K.-P. L., & Nicoletti, R. (2003). Does right-left prevalence occur for the Simon effect? Attention, Perception, & Psychophysics, 65(8), 1318–1329.
Reeve, T. G., & Proctor, R. W. (1984). On the advance preparation of discrete finger responses. Journal of Experimental Psychology: Human Perception and Performance, 10(4), 541–553.
Resulaj, A., Kiani, R., Wolpert, D. M., & Shadlen, M. N. (2009). Changes of mind in decision-making. Nature, 461(7261), 263–266.
Ridderinkhof, R. K. (2002). Micro-and macro-adjustments of task set: activation and suppression in conflict tasks. Psychological Research Psychologische Forschung, 66(4), 312–323.
Rogers, R. D., & Monsell, S. (1995). Costs of a predictable switch between simple cognitive tasks. Journal of Experimental Psychology: General, 124(2), 207–231.
Rosenbaum, D. A. (1983). The movement precuing technique: Assumptions, applications, and extensions. In R. A. Magill (Ed.), Memory and control of action (pp. 231–274). Amsterdam: North-Holland.
Rubichi, S., & Pellicano, A. (2004). Does the Simon effect affect movement execution? European Journal of Cognitive Psychology, 16(6), 825–840.
Rubichi, S., Nicoletti, R., & Umiltà, C. (2005). Right-left prevalence with task-irrelevant spatial codes. Psychological Research Psychologische Forschung, 69(3), 167–178.
Scherbaum, S., Dshemuchadse, M., Fischer, R., & Goschke, T. (2010). How decisions evolve: The temporal dynamics of action selection. Cognition, 115(3), 407–416.
Schmidt, J. R. (2019). Evidence against conflict monitoring and adaptation: An updated review. Psychonomic Bulletin & Review, 26(3), 753–771.
Schmidt, J. R., & De Houwer, J. (2011). Now you see it, now you don’t: Controlling for contingencies and stimulus repetitions eliminates the Gratton effect. Acta Psychologica, 138(1), 176–186.
Simon, J. R., & Rudell, A. P. (1967). Auditory SR compatibility: The effect of an irrelevant cue on information processing. Journal of Applied Psychology, 51(3), 300–304.
Smith, G. A., & Carew, M. (1987). Decision time unmasked: Individuals adopt different strategies. Australian Journal of Psychology, 39(3), 339–351.
Stürmer, B., Leuthold, H., Soetens, E., Schröter, H., & Sommer, W. (2002). Control over location-based response activation in the Simon task: Behavioral and electrophysiological evidence. Journal of Experimental Psychology: Human Perception and Performance, 28(6), 1345–1363.
Töbel, L., Hübner, R., & Stürmer, B. (2014). Suppression of irrelevant activation in the horizontal and vertical Simon task differs quantitatively not qualitatively. Acta Psychologica, 152, 47–55.
Verguts, T., & Notebaert, W. (2008). Hebbian learning of cognitive control: Dealing with specific and nonspecific adaptation. Psychological Review, 115(2), 518–525.
Verguts, T., & Notebaert, W. (2009). Adaptation by binding: A learning account of cognitive control. Trends in Cognitive Sciences, 13(6), 252–257.
Verguts, T., Notebaert, W., Kunde, W., & Wühr, P. (2011). Post-conflict slowing: cognitive adaptation after conflict processing. Psychonomic Bulletin & Review, 18(1), 76–82.
Vu, K.-P. L., & Proctor, R. W. (2001). Determinants of right-left and top-bottom prevalence for two-dimensional spatial compatibility. Journal of Experimental Psychology: Human Perception and Performance, 27(4), 813–828.
Vu, K.-P. L., & Proctor, R. W. (2002). The prevalence effect in two-dimensional stimulus-response compatibility is a function of the relative salience of the dimensions. Perception & Psychophysics, 64(5), 815–828.
Vu, K.-P. L., Pellicano, A., & Proctor, R. W. (2005). No overall right-left prevalence for horizontal. Perception & Psychophysics, 67(5), 929–938.
Vu, K. P. L., Proctor, R. W., & Pick, D. F. (2000). Vertical versus horizontal spatial compatibility: Right-left prevalence with bimanual responses. Psychological Research Psychologische Forschung, 64(1), 25–40.
Weissman, D. H. (2019a). Interacting congruency effects in the hybrid Stroop-Simon task prevent conclusions regarding the domain specificity or generality of the congruency sequence effect. Journal of Experimental Psychology. Learning, memory, and cognition.
Weissman, D. H. (2019b). Let your fingers do the walking: Finger force distinguishes competing accounts of the congruency sequence effect. Psychonomic Bulletin & Review, 26, 1619–1626.
Weissman, D. H., Colter, K., Drake, B., & Morgan, C. (2015). The congruency sequence effect transfers across different response modes. Acta Psychologica, 161, 86–94.
Weissman, D. H., Jiang, J., & Egner, T. (2014). Determinants of CSEs without learning and memory confounds. Journal of Experimental Psychology: Human Perception and Performance, 40(5), 2022–2037.
Wiegand, K., & Wascher, E. (2005). Dynamic aspects of stimulus-response correspondence: Evidence for two mechanisms involved in the Simon effect. Journal of Experimental Psychology: Human Perception and Performance, 31(3), 453–464.
Wiegand, K., & Wascher, E. (2007). The Simon effect for vertical S-R relations: changing the mechanism by randomly varying the S–R mapping rule? Psychological Research Psychologische Forschung, 71(2), 219–233.
Wühr, P. (2005). Evidence for gating of direct response activation in the Simon task. Psychonomic Bulletin & Review, 12(2), 282–288.
Wühr, P., Duthoo, W., & Notebaert, W. (2015). Generalizing attentional control across dimensions and tasks: Evidence from transfer of proportion-congruent effects. The Quarterly Journal of Experimental Psychology, 68(4), 779–801.
Wylie, S. A., van den Wildenberg, W. P. M., Ridderinkhof, K. R., Bashore, T. R., Powell, V. D., Manning, C. A., et al. (2009). The effect of Parkinson's disease on interference control during action selection. Neuropsychologia, 47(1), 145–157.
Zhang, J., & Kornblum, S. (1997). Distributional analysis and De Jong, Liang, and Lauber’s (1994) dual-process model of the Simon effect. Journal of Experimental Psychology: Human Perception and Performance, 23(5), 1543.
Zhang, H. H., Zhang, J., & Kornblum, S. (1999). A parallel distributed processing model of stimulus–stimulus and stimulus–response compatibility. Cognitive Psychology, 38(3), 386–432.
Acknowledgements
We would like to thank Robert Proctor for his detailed comments on a preliminary version of the paper. This research was supported by the Korean Research Foundation Grant funded by the Korean Government (NRF-2016R1D1A1A09918865). The raw datasets and the analysis scripts are available at the Open Science Framework: https://osf.io/6jmpa/?view_only=ab275590e9a0478ea2679485f2c4f1a0.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Lim, C.E., Cho, Y.S. Response mode modulates the congruency sequence effect in spatial conflict tasks: evidence from aimed-movement responses. Psychological Research 85, 2047–2068 (2021). https://doi.org/10.1007/s00426-020-01376-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-020-01376-3