
Abstract
Dual-task performance typically leads to performance impairments in comparison to single tasks (i.e., dual-task costs). The literature discusses the contribution to these dual-task costs due to (1) bottleneck limitations in the dual-component tasks and (2) executive control processes regulating access to this bottleneck. Previous studies investigated the characteristics of executive control processes primarily triggered by external stimulus information. In the present study, however, we investigated the existence as well as the characteristics of internally triggered and driven endogenous control processes to regulate bottleneck access. In detail, we presented dual-task blocks with varying task orders and informed participants in advance about repetitions of the same task order as well as switches between different task orders (i.e., task-order repetitions and switches were predictable). Experiment 1 demonstrated that task-order information and an increased preparation time generally increase the efficiency for endogenous task-order control and improves preparation for task-order switches. This finding is basically consistent with the assumption of the existence of endogenous control processes. Experiment 2, however, did not provide evidence that this endogenous control is related with working-memory maintenance mechanisms. Experiment 3 showed that endogenous control does not only fully complete task-order preparation but also requires exogenous, stimulus-driven components.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A common real-world experience is that attempting to perform two tasks simultaneously compromises performance in one or both of these tasks, that is: there are dual-tasks costs that manifest in an increase in errors and/or the time needed to perform the two tasks, as compared to their isolated performance in single tasks (e.g., Kahneman, 1973; Welford, 1952). Studies in the context of the ‘Psychological refractory period’ (PRP) paradigm investigated these costs through the presentation of two stimuli with a variable, short delay between their onsets (stimulus onset asynchrony; SOA), and participants have to make choice reaction time (RT) responses to these stimuli. The typical finding is that RTs to the second (task) stimulus (RT2) increases with decreasing SOA, (e.g., Pashler, 1994; Schubert, 2008); this RT2 pattern is referred to as the ‘PRP effect’.
The PRP effect has been attributed to capacity limitations in resource sharing models assuming that two tasks can simultaneously share processing resources, so that neither task in dual tasks is performed as quickly as it would be performed in single tasks (e.g., Hazeltine, Ruthruff, & Remington, 2006; Meyer & Kieras, 1997; Miller & Durst, 2015; Miller, Ulrich, & Rolke, 2009; Mittelstädt & Miller, 2017); importantly, the amount of capacity shared between tasks is subject to a strategic choice in the context of resource sharing models. Supportive evidence for these models is an increase with decreasing SOA not only of RT2, but also of RTs to the first (task) stimulus (RT1; Strobach, Schütz, & Schubert, 2015). The specific case of a complete distribution of processing resources to one component task and no such resources shared with the other task is consistent with assumptions of the central bottleneck model; however, the central bottleneck model assumes a structural and unavoidable bottleneck limitation. According to this model, a processing bottleneck limitation structurally prevents the two tasks to be performed in parallel. Specifically, this bottleneck limitation is located at a central processing stage between stimulus perception and response execution, namely the response selection stage (e.g., Pashler, 1994; Welford, 1952, 1980). Irrespective of its nature (i.e., strategic capacity limitation or structural bottleneck limitation), the notion of a processing-limited central response selection accounts for the PRP effect by assuming that, if the two tasks are presented with short SOAs between task stimulus 1 and task stimulus 2, response selection for the two tasks operates serially, while the initial perception and final response execution stages of the two tasks can operate in parallel. In other words, response selection of the second task is postponed until the end of this stage in the first task and until the first task has left the bottleneck stage. While this limitation assumption provides a commonly accepted explanation for a variety of findings in dual-task research, the following questions remain unresolved: how is the capacity limitation (e.g., the bottleneck mechanism) allocated to the two task streams? And how is the task processing order controlled and determined? In the present study, we investigated the assumption that the order in which two component tasks are performed at the limitations might be planned and prepared in advance due to internally driven endogenous control processes.
Determination of task-order processing: endogenous and exogenous control
The regulation of task order in dual tasks has been investigated in a specific variety of the PRP paradigm (Kübler, Reimer, Strobach, & Schubert, 2018; Luria & Meiran, 2003; Sigman & Dehaene, 2006; Szameitat, Lepsien, von Cramon, Sterr, & Schubert, 2006). In this variety, participants are asked to execute two-choice RT tasks on two target stimuli that are presented in quick succession. Importantly, the presentation order of both stimuli varies randomly from trial to trial and participants are instructed to execute the two tasks according to the order of stimulus presentation, resulting in repetitions of the same task order and switches between different task orders in successive trials. By applying this paradigm, two potential mechanisms of task-order regulation have been identified: One possibility to characterize order control is that bottleneck processing is simply recruited by the stimuli on an exogenous first-come-first-served basis, so that the order in which the tasks are handled is determined by which of the two task stimuli arrives at the bottleneck first. Recent studies provided empirical evidence that is consistent with the predictions of this first-come-first-served mode (Hendrich, Strobach, Buss, Müller, & Schubert, 2012; Leonhard, Ruiz Fernández, Ulrich, & Miller, 2011; Ruiz Fernández, Leonhard, Rolke, & Ulrich, 2011; Sigman & Dehaene, 2006; Strobach, Hendrich, Kübler, Müller, & Schubert, 2018). The second possible mechanism assumes that the order in which the tasks are performed might be planned and prepared in advance due to stimulus presentation and internally driven endogenous control processes (de Jong, 2000; Schubert, 2008; Szameitat et al., 2006). The assumption of such an endogenous model is consistent with the general idea of intentional, self-instigated, goal-driven processes of executive control (Band & van Nes, 2006; Logan, 2003; Logan & Bundesen, 2003; Monsell & Mizon, 2006) without foreknowledge due to task stimulus information (Baddeley, 2012; Norman & Shallice, 1986; Rogers & Monsell, 1995; W. Schneider & Shiffrin, 1977). The central issue addressed in the present research concerns (1) whether these endogenous control components are involved in task-order processing of dual tasks and (2) the nature of this type of dual-task control.
One explicit test of the endogenous model could require an experimental situation in which subjects have the chance to prepare for a specific task order in a dual-task situation with variable task orders, i.e., with task-order repetitions and switches relative to the preceding trial. In this situation, improvements of dual-task performance compared to a similar dual-task situation that, in contrast, does not allow for the preparation of the appropriate task order would indicate the existence of endogenous control processes that prepare and schedule the processing order of the component tasks in advance, i.e., before the presentation of the stimuli.
Direct evidence regarding the existence of endogenous preparation of task order in dual tasks still appears to be scarce. This is so, because most of the studies investigating dual-task situations with varying task orders and, thus, task-order switches as well as task-order repetitions realized no manipulation of the available information about task order before stimulus onset (Hendrich et al., 2012; Hirsch, Nolden, & Koch, 2017; Ruiz Fernández et al., 2011; Sigman & Dehaene, 2006; Strobach et al., 2018; Szameitat et al., 2006; Töllner, Strobach, Schubert, & Müller, 2012). Instead, task order was exclusively determined by the presentation order of the two stimuli. So, it might be that the bottleneck processing stage is not explicitly allocated to the information streams of the component tasks by endogenous and preparatory processes but, instead, could be subjected to the first-come-first-served principle. That is, bottleneck processing would be allocated passively to the two tasks based on their central arrival times as proposed by the exogenous model.
One way to investigate the existence of endogenous task-order preparation and to provide information about task order before stimulus onset might be to provide cues indicating the stimulus order of the component tasks. de Jong (1995; see also Luria & Meiran, 2003, 2006, for a similar approach with top–down and bottom–up processes) investigated endogenous preparation with the use of such cues under valid and invalid conditions. Under valid conditions, the cues correctly prompted the task order, while incorrect task order was prompted under invalid conditions. Consistent with the assumption of the endogenous model, de Jong found performance improvements under valid conditions; that is, under these conditions, RTs, error rates, and response reversals (i.e., the response order is different to the order of stimulus presentation) were decreased. These improvements can be explained with an endogenous preparation of task order as following: Based on cue information, participants prepare for a specific task order. This prepared order, however, contradicts the stimulus-presentation order under invalid conditions and participants have to reconfigure their processing order after stimulus presentation which leads to impaired task performance.
However, the situation of de Jong (1995) is not conclusive to investigate task-order processing in the context of dual tasks with variable order, because his paradigm included confounds of variable stimulus-presentation order and variable cue identity. Studies of task switching and, thus, cognitive flexibility have investigated the crucial role of the latter on task preparation. In these studies (Kiesel et al., 2010; Koch, Poljac, Müller, & Kiesel, 2018; Monsell, 2003; Rubinstein, Meyer, & Evans, 2001; Sudevan & Taylor, 1987), two different tasks are presented in random order on a trial-by-trial basis resulting in an alternation between different tasks or repetitions of the same task; thus, in contrast to the dual-task paradigm, participants performed only one task per trial in task-switching situations. Usually, both tasks are executed on the same set of stimuli, and as a result, these tasks are frequently afforded by distinctly different perceptual dimensions thereof, such as when participants switch between form classification and color classification when presented with colored geometrical forms. In the task-cuing procedure, participants are informed about the identity of the upcoming task by an informative cue that precedes or accompanies the presentation of the target stimulus (e.g., Meiran, 1996). Switching between tasks (i.e., executing a different task on the current trial than on the directly preceding trial) incurs a cost in RTs and sometimes error rates in comparison to task repetitions (i.e., executing the same task on successive trials). However, these task switch costs (overview in Kiesel et al., 2010; Monsell, 2003; Vandierendonck, Liefooghe, & Verbruggen, 2010), can also be explained by switches between different cues associated with switches between different tasks (Logan & Bundesen, 2003; Mayr & Kliegl, 2003; D. W. Schneider & Logan, 2007); thus, there is a confound between cue switch and task switch to explain task-switch costs. Equivalent to this confound in the task-switching context, dual tasks with varying task orders introduced by de Jong (1995) realized a confounded procedure for investigating task-order switches. Whenever task-order switches in de Jong’s dual-task situation, cue identity also switches as well, while on order repetitions, there was no cue identity switch. As a result, there was no valid investigation of varying task orders in this study, since this study might have investigated rather cue switching than task-order switching. In sum, previous studies investigating potential endogenous preparation of task order in dual tasks either informed about task order by stimulus-presentation order (e.g., Sigman & Dehaene, 2006; Szameitat et al., 2006) or by providing cues informing about task order (de Jong, 1995). In the former procedure, however, it is unclear whether task-order preparation was endogenous since this preparation could be based on stimulus order and reflects, thus, exogenous preparation exclusively. The latter procedure is not completely valid since it might have investigated rather cue switching than task-order switching and preparation. In the present study, we, therefore, introduced a rather novel procedure to investigate endogenous task-order preparation.
The present study
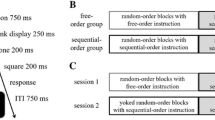
Similar to previous studies in this field, we presented dual tasks with different stimulus-presentation orders and asked participants to perform both tasks in these orders. However, these orders were not randomly mixed within blocks, but their sequence was varied systematically and participants were explicitly informed about these systematic sequences before the start of these systematic-order blocks (for a similar approach see de Jong, 1995, Experiment 3); in this way, participants were informed about task-order switches and repetitions in advance (Fig. 1). This type of manipulation is equivalent to the manipulation of task sequences in the alternating runs paradigm in the context of task switching in which the required task was indicated by pre-specified task sequences (e.g., participants are instructed to switch tasks on every second trial; Karbach & Kray, 2009; Rogers & Monsell, 1995). In the following experiments, we aimed to provide evidence for endogenous task-order preparation processes and how they are specified (Experiment 1), how these processes relate to other cognitive components, namely working memory (Experiment 2), and whether these processes are sufficient for preparing and planning the processing order of the component tasks in a dual-task situation (Experiment 3).

Illustration of systematic trial order as demonstrated to participants in Experiment 1 and 2 (a) as well as in Experiment 3 (b)
General methods
Participants
Participants were students of the Medical School Hamburg and other universities in the Hamburg area, recruited via online databases and personal contacts. All participants were right-handed and German native-speakers. They reported normal or corrected-to-normal hearing and vision, and received course credit for their participation.
Apparatus
Visual stimuli in the following experiments were presented on a 22-inch color monitor (Refresh rate: 60 Hertz; viewed from a distance of approximately 60 cm) and auditory stimuli were presented via headphones which were connected to IBM-compatible personal computers. Experiments were controlled by the experimental software package Presentation (Version 18).
Stimuli
Participants performed a visual and an auditory choice RT task in the present dual-task test. The auditory task included the presentation of sine-wave tones with pitches of 350, 900, or 1650 Hz. Participants responded with the index, middle, and ring finger of the right hand, respectively. The visual task included the presentation of small, middle, and large visually presented triangles and responses with the ring, the middle, and the index finger of the left hand, respectively. Under conditions of low task difficulty, only 2 stimuli were presented in each task (i.e., lowest and highest tone in the auditory task, small and large triangle in the visual task), while there were all 3 stimuli in each task under conditions of high difficulty.
Procedure and design
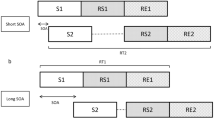
Participants performed single-task blocks in which only 1 of the 2 tasks were presented. They also performed dual-task blocks that included the presentation of both tasks. Trials of single-task blocks started with the presentation of 3 dashes next to each other, of which the middle dash was located at the center of the screen. The dashes remained on the screen until the end of each trial, while they disappeared between trials. An auditory stimulus (i.e., a sine-wave tone) appeared for 100 ms in auditory single-task block trials, or a visual stimulus (i.e., a triangle) appeared centrally in the visual single-task block trials 500 ms after onset of the presentation of the dashes and, thus, trial start; visual stimuli were presented until response or a maximum of 2500 ms. Similar to single-task trials, dual-task block trials started with the presentation of 3 white dashes that remained on screen until the end of each trials, but disappeared between trials. After 500 ms, a first stimulus (i.e., auditory or visual) was presented, followed by the presentation of the second stimulus (i.e., visual or auditory). The interval between the onsets of both stimuli (i.e., SOA) was 200 ms. Incorrect trials were completed with an error feedback (German word: “Fehler”) for 1500 ms; incorrect trials included wrong or omitted responses as well as response reversals.
Single-task blocks consisted of 36 single-task trials and stimuli were presented with equal frequency in a random order. In all 36 trials of the dual-task blocks, auditory and visual stimuli were presented with equal frequency and stimuli were selected randomly. The stimulus order was counterbalanced within blocks. Participants were instructed to respond as quickly and as accurately as possible in single-task blocks as well as in the dual-task blocks. In dual-task block trials, priority was instructed for the firstly presented stimulus.
Across the experiments of this study, we conducted different types of dual-task blocks with different task orders: dual-task blocks with random task order (random dual tasks) and dual-task blocks with systematic task order (systematic dual tasks). Dual-task trials with a first auditory stimulus (auditory–visual order trials) and with a first visual stimulus (visual–auditory order trials) were randomly mixed in random-order dual-task blocks. Systematic-order dual-task blocks included trials in a fixed, predictable, and pre-specified order, resulting in 50% order-repetition and 50% order-switch trials. This order was verbally instructed and visually illustrated to participants before the block start (Fig. 1). Half of these blocks started with a trial with a first visual stimulus and the remaining blocks started with a trial with a first auditory stimulus. In random and systematic dual-task blocks, the frequencies of auditory–visual order and visual–auditory order trials within each block were equal.
At the beginning of each experiment, one visual and 1 auditory single-task block was presented. Whereas half of the participants started with a visual block followed by an auditory block, the remaining participants performed the blocks in the opposite order. Following, two dual-task blocks with a fixed task order were conducted. Whereas half of the participants started with a dual-task block and trials with a first auditory stimulus followed by a block with trials with a first visual stimulus, the remaining participants performed the blocks in the opposite order. After this initial practice phase, the dual-task test phase started. The remaining dual-task blocks during dual-task test will be specified in the Methods sections of the individual experiments.
Experiment 1
Experiment 1 investigated the potential impact of endogenous preparation of dual-task processing order in a context in which such a preparation is available in contrast to a context without the potential of such preparation. To do so, we presented dual-task blocks with systematic task order as well as dual-task blocks with random task order; both block types include varying task-order trials, resulting in order switches and order repetitions. Further, to investigate the impact of preparation intervals on preparing task order, we introduced different preparation intervals between trials of random-order and systematic-order blocks (i.e., inter-trial intervals [ITIs] of 700 ms, 1200 ms, and 2500 ms). Our hypotheses in Experiment 1 are as following: If endogenous preparation exists and has a general benefit for the maintenance, selection, and implementation of task orders then performance should be particularly improved under systematic-order conditions (i.e., reduced RTs, error rates, and/or response reversals) in comparison to random-order conditions. And if the realization of the endogenous preparation requires time, this general advantage would be increased with long ITIs in comparison to short ITIs. An addition, it could be that if the existence of endogenous preparation of task orders in systematic blocks specifically improves the reconfiguration for a new task-order order, then this type of preparation improves task-order switches. Also, if the prolongation of ITIs optimizes this reconfiguration, the benefit for order switches in systematic blocks increases with longer ITIs.
Methods
Stimuli, procedure, and design were identical to the general methods described above, with the following specifications. In Experiment 1, tasks were performed under high difficulty conditions (i.e., three stimuli per task). This experiment included the following block order during the dual-task test: four dual-task blocks with random task order, eight dual-task blocks with systematic task order, followed again by 4 dual-task blocks with random task order. In systematic task-order blocks, task orders were switched every second trial (Fig. 1a). Dual-task blocks were performed with 3 ITI levels across all blocks, namely 700 ms, 1200 ms, and 2500 ms, varied between three groups of participants (so, one of the 3 ITI levels was chosen for each subject, staying consistent across all blocks). Thirty-two, 30, and 30 participants conducted the experiment versions with ITIs of 700 ms (22 female, mean age: 23.5 years, age range: 18–26 years), 1200 ms (20 female, mean age: 22.8 years, age range: 18–25 years), and 2500 ms (18 female, mean age: 23.8 years, age range: 19–26 years), respectively. Each experimental session lasted approximately 60 min.
Results
Before analyzing RTs, error rates, and response reversal rates, we excluded those participants with total error rates of more than 25% across the dual-task blocks. As a consequence, we included 19, 22, and 26 participants in the experiment versions with ITIs of 700 ms, 1200 ms, and 2500 ms, respectively. RTs (after the exclusion of trials with errors on the choice decisions within the component tasks and response reversals), error rates, and response reversal rates (after the exclusion of trials with errors on the choice decisions within the component tasks) were aggregated across visual–auditory and auditory–visual order trials in the dual-task blocks. The first trial of each block was not analyzed.
We included the error rate and RT dual-task data of the trials with same task order and different task order across blocks with systematic task orders as well as blocks with random task orders into the analyses of Task 1 (firstly presented task) and Task 2 (secondly presented task) performance. The performance of both tasks were separately analyzed in mixed measures ANOVAs including the within-subjects factors TRIALTYPE (same order vs. different order) and BLOCKTYPE (systematic order vs. random order) as well as the between-subjects factor ITI (700 ms, 1200 ms, and 2500 ms).
RTs (Fig. 2) Analyzing RT1, the main effect of TRIALTYPE was significant, F(1, 64) = 170.050, p < .001, ŋp2 = .73, generally replicating findings of order-switch costs in previous studies (e.g., Kübler et al., 2018; Szameitat et al., 2006): same-order trials showed lower RTs (M = 1374 ms) than different-order trials (M = 1477 ms). Further, BLOCKTYPE was also significant, F(1, 64) = 148.650, p < .001, ŋp2 = .70, demonstrating generally faster RTs in systematic-order blocks (M = 1323 ms) in comparison to random-order blocks (M = 1528 ms). This finding demonstrated a general endogenous control benefit. The RT1 analysis showed no interaction including the factors TRIALTYPE and BLOCKTYPE, Fs < 1.411, ps > .24. Thus, this analysis provided no evidence that information about task order modulates the order-switch costs across all or within ITI conditions. Also, the main effect of ITI, F(2, 64) = 1.700, p > .19, as well as the interactions BLOCKTYPE x ITI and TRIALTYPE x ITI were non-significant, Fs(2, 64) < 1.550, ps > .22.

Reaction times (RTs) in ms in Experiment 1 across the groups of participants with inter-trial intervals (ITI) of 700 ms, 1200 ms, and 2500 ms, in blocks with random dual-task orders and systematic dual-task orders as well as in trials with repetitions of task order (same) and switches between different task orders (diff.). a RT1. b RT2. Error bars represent standard errors of the mean
Analyzing RT2 similar to RT1, the main effect of TRIALTYPE was significant, F(1, 64) = 161.928, p < .001, ŋp2 = .72, generally replicating findings of order-switch costs in previous studies (e.g., Kübler et al., 2018; Szameitat et al., 2006): same-order trials showed lower RTs (M = 1478 ms) than different-order trials (M = 1584 ms). Furthermore, BLOCKTYPE was also significant, F(1, 64) = 117.757, p < .001, ŋp2 = .65, demonstrating generally faster RTs in systematic-order blocks (M = 1433 ms) in comparison to random-order blocks (M = 1628 ms). This finding demonstrated the endogenous control benefit on the maintenance of different task orders in the RT2 data. The present analysis showed no significant interaction including the factors TRIALTYPE and BLOCKTYPE, Fs < 3.214, ps > .08. Thus, there is no convincing evidence that information about task order modulates RT2’s order-switch costs across all or within ITI conditions. Also, the main effect of ITI, F(2, 64) = 2.412, p > .10, as well as the interaction BLOCKTYPE x ITI and TRIALTYPE x ITI was non-significant, Fs(2, 64) < 1.
Errors (Table 1) The error analysis of the first task was equivalent to the RT analysis and revealed main effects for TRIALTYPE, F(1, 64) = 113.013, p < .001, ŋp2 = .64, BLOCKTYPE, F(1, 64) = 140.615, p < .001, ŋp2 = .69, and ITI, F(2, 64) = 13.127, p < .001, ŋp2 = .29. Error rates were lower in same-order trials (M = 12.4%) than in different-order trials (M = 17.2%); they were lower under the systematic-order condition (M = 9.6%) compared to the random-order condition (M = 20.0%); and participants with ITIs of 2500 ms showed the lowest error rates (M = 8.9%), followed by participants under ITI condition of 1200 ms (M = 14.0%), and of 700 ms (M = 21.5%), all ps < .05. The interaction of BLOCKTYPE and TRIALTYPE was significant, F(1, 64) = 20.559, p < .001, ŋp2 = .24, demonstrating lower order-switch costs in systematic-order blocks (M = 2.8%) than in random-order blocks (M = 7.0%), both ps < .05. BLOCKTYPE interacted with ITI, F(2, 64) = 19.459, p < .001, ŋp2 = .38, revealing that the error rates in systematic-order blocks in comparison to random-order blocks were significantly lower under the ITI condition of 1200 ms (M = 17.0%) and 2500 ms (M = 10.7%), both ps < .05, while there was no significant difference between systematic- and random-order blocks under the ITI condition of 700 ms (M = 3.2%), p = .051; this is general evidence for endogenous task preparation. TRIALTYPE also interacted with ITI, F(2, 64) = 18.418, p < .001, ŋp2 = .37, demonstrating significant order-switch costs under the ITI condition of 1200 ms (M = 8.3%) and 2500 ms (M = 5.1%), both ps < .05, while there was no significant evidence for such costs under the 700-ms ITI condition (M = 1.2%), p = .16. All factors, BLOCKTYPE x TRIALTYPE x ITI, interacted, F(2, 64) = 4.545, p < .05, ŋp2 = .12. This three-way interaction revealed that the reduced order-switch costs under systematic in comparison to random-order conditions (i.e., the interaction of BLOCKTYPE x TRIALTYPE) was exclusively present when the ITI was 1200 ms, F(1, 21) = 12.524, p < .01, ŋp2 = .37, and 2500 ms, F(1, 25) = 16.468, p < .001, ŋp2 = .40, but not with an IRI of 700 ms, F(1, 18) < 1.
The error analysis of the second task revealed main effects for TRIALTYPE, F(1, 64) = 93.116, p < .001, ŋp2 = .59, BLOCKTYPE, F(1, 64) = 133.513, p < .001, ŋp2 = .68, and ITI, F(2, 64) = 12.838, p < .001, ŋp2 = .29. Error rates were lower in same-order trials (M = 14.5%) than in different-order trials (M = 19.0%); they were lower under the systematic-order condition (M = 11.9%) compared to the random-order condition (M = 21.6%); and participants with ITIs of 2500 ms (M = 10.9%) and of 1200 ms (M = 15.7%) showed no conclusive evidence for differences in error rates, p = .055, while the error rates under the ITI condition of 700 ms (M = 23.6%) was increased, both ps < .01. The interaction of BLOCKTYPE and TRIALTYPE was significant, F(1, 64) = 14.746, p < .001, ŋp2 = .19, demonstrating lower order-switch costs in systematic-order blocks (M = 2.6%) than in random-order blocks (M = 6.3%), both ps < .001. BLOCKTYPE interacted with ITI, F(2, 64) = 23.567, p < .001, ŋp2 = .42, revealing that the error rates in systematic-order blocks in comparison to random-order blocks were significantly lower under the ITI condition of 1200 ms (M = 16.5%) and 2500 ms (M = 10.8%), both ps < .001, while there was no evidence for a significant difference under the ITI condition of 700 ms (M = 1.9%), p = .24. Similarly, TRIALTYPE also interacted with ITI, F(2, 64) = 18.885, p < .001, ŋp2 = .37, demonstrating significant order-switch costs under the ITI condition of 1200 ms (M = 8.1%) and 2500 ms (M = 4.5%), both ps < .001, while there was no significant evidence for such costs under the 700-ms ITI condition (M = .9%), p = .33. The combination of all factors, BLOCKTYPE x TRIALTYPE x ITI, interacted, F(2, 64) = 3.965, p < .05. This three-way interaction revealed that the reduced order-switch costs under systematic in comparison to random-order conditions (i.e., the interaction of BLOCKTYPE x TRIALTYPE) was exclusively present when the ITI was 1200 ms, F(1, 21) = 8.845, p < .01, ŋp2 = .28, and 2500 ms, F(1, 25) = 9.810, p < .001, ŋp2 = .36, but not with an IRI of 700 ms, F(1, 18) < 1.
Response reversal rates (Fig. 3) The analyses of the response reversal rates revealed main effects for TRIALTYPE, F(1, 64) = 138.706, p < .001, ŋp2 = .68, and BLOCKTYPE, F(1, 64) = 127.171, p < .001, ŋp2 = .67. The response reversal rates were significantly reduced in same-order trials (M = 5.9%) than in different-order trials (M = 11.1%) and these rates were also reduced under systematic conditions (M = 4.4%) in comparison to random-order conditions (M = 12.7%). The combination of both factors interacted significantly, F(1, 64) = 62.070, p < .001, ŋp2 = .49, revealing that the difference between response reversal rates in different-order trials and same-order trials was increased under random-order conditions (M = 8.2%) in comparison to this difference under systematic-order conditions (M = 2.3%). This finding demonstrates that participants’ endogenous preparation benefited from information about the upcoming order and this benefit was particularly relevant for task-order switches. The interaction of TRIALTYPE and ITI, F(1, 64) = 62.070, p < .001, ŋp2 = .49, demonstrated increased response reversal rates in switch-order trials after an ITI of 700 ms (M = 6.9%) in contrast to ITIs of 1200 ms (M = 4.7%) and 2500 ms (M = 4.2%). However, this data pattern was similar across random and systematic-order conditions, F(2, 64) = 1.051, p > .36. The combination of BLOCKTYPE and ITI was also non-significant, F(2, 64) = 1.923, p > .16.

Response reversal rates in percent in Experiment 1 across the groups of participants with inter-trial intervals (ITI) of 700 ms (a), 1200 ms (b), and 2500 ms (c), in blocks with random dual-task orders and systematic dual-task orders as well as in trials with repetitions of task order (same) and switches between different task orders (different). Error bars represent standard errors of the mean
Note that due to the exclusion criterion and upper cap of 25% of errors, we excluded different numbers of participants in each ITI group (i.e., 700 ms: N = 13; 1200 ms: N = 8; 2500 ms: N = 4). We note that this exclusion criterion might have introduced an artificial bias into these groups with more participants with high error rates excluded and generally higher skilled participants in the group of the ITI 700 ms condition in contrast to the alternative groups. Thus, the group with an ITI of 700 ms might have been less heterogeneous than these alternative groups. To validate our results of Experiment 1 in more heterogeneous groups, we repeated the RT and error analyses including all participants regardless of the exclusion criterion. In sum, these analyses showed a pattern similar to the analyses including this criterion reported in detail above. Thus, the present findings also hold in more heterogeneous samples.
Discussion
The present experiment aimed to provide empirical evidence and investigated the characteristics of endogenous preparation in dual tasks with different task orders; in detail, we investigated the endogenous preparation of dual tasks with same and different orders between successive trials. RTs, error data, and response reversal rates consistently revealed two conclusions. First, dual-task performance with different task orders after an order switch results in order-switch costs, i.e., performance is impaired under different-order conditions compared with performance under same-order conditions. This result is consistent with previous studies (e.g., Szameitat et al., 2006). Further, information about the upcoming task order leads to general performance improvements in contrast to conditions under which no such information is provided. We, thus, conclude that the general maintenance, selection, and implementation of different task orders are improved by endogenous control and that our method to provide participants with information about task order to allow predictions about this order was generally effective. This general view on endogenous dual-task control is consistent with conclusions from previous studies (e.g., Band & van Nes, 2006; de Jong, 2000).
Importantly, the error data and the response reversal rates revealed that, in addition to the general improvement due to task-order preparation processes, performance in trials in which task order changes relative to the preceding trial specifically benefits from endogenous preparation. More specifically, the error data demonstrated that with increased ITIs before the start of a trial with a predictable order switch, performance is improved in contrast to decreased ITIs. This improvement is demonstrated by reduced order-switch costs with ITIs of 1200 ms and 2500 ms in comparison to ITIs of 700 ms. That is, with a prolonged interval before an order switch, there is more time to correctly prepare for this switch. Since this correct preparation is anticipatory before the start of dual-task trials including this switch, we interpret this finding as evidence for the existence of endogenous preparation of task order in dual tasks. The response reversal rates demonstrated an increased rate of two tasks performed in the wrong order in order-switch trials in comparison to order-repetition trials. However, this impairment in order-switch trials was reduced when participants have the opportunity to correctly prepare for an order switch in systematic dual-task blocks. Importantly, the correctness of this preparation increases with prolonged preparation time (i.e., with prolonged ITIs). This result pattern suggests that endogenous task-order preparation specifically improves the reconfiguration of a new task order. However, the RT data provided no convincing evidence for endogenous task-order preparation since there is no interaction including the ITI factor in the RT analyses. There are two speculative explanations for this lacking evidence which we cannot differentiate at this point. First, it might be that endogenous task-order preparation leads to an increase in the correctness of task-order preparation. However, this preparation does not increase the processing speed and thus the processing efficiency of this type of preparation. Second, it might be that participants strategically put more effort into correct task-order preparation instead of fast preparation of task order and the effect of this effort increased with prolonged ITI. Such a strategic effect might occur although participants were instructed to focus on both speed and correctness (i.e., participants were instructed to respond as quickly and as accurately as possible). The following experiments aimed to specify the processing characteristics of this endogenous dual-task control (Experiment 2) and its relation to exogenous, externally triggered components of this control (Experiment 3).
Experiment 2
While Experiment 1 provided first evidence for endogenous preparation of task order in general and task-order switches in particular, Experiment 2 further explores the characteristics of this order preparation and the specific factors which determine the efficiency of this preparation and the related cognitive components. In the present experiment, we addressed the question whether a varying working-memory (WM) capacity may affect the efficiency of endogenous preparation of task-order switches, and thus whether the WM component is involved in this switch preparation. This component is of special interest here because converging evidence from different research areas suggests that executive control and WM processes rely on related mechanisms. That is, many authors assumed that task representations are maintained in WM during ongoing task processing with one (Braver et al., 1997; Dreisbach & Haider, 2009; Kane & Engle, 2003; MacDonald, Cohen, Stenger, & Carter, 2000) or multiple tasks (Hartley & Little, 1999; Huestegge & Koch, 2009; Maquestiaux, Hartley, & Bertsch, 2004; Schubert & Strobach, 2018). For instance, we showed that task information of both component tasks will be activated in WM at the beginning of a dual-task trial (Schubert & Strobach, 2018). While this activation may be incomplete or ineffective at the beginning of practice and after separate practice of component tasks under single-task practice condition, dual-task practice of both component tasks together enables the activation of task order information in WM. It is open, however, whether the maintenance of task-order information and the preparation of task-order switches are related with WM capacity.
We varied WM capacity by varying (1) load of a WM task and (2) complexity of the component tasks that constitute the dual task. To vary the WM load, we presented dual-task blocks with systematic task order under low and high WM load conditions, i.e., participants had to maintain one or four digits across blocks. If WM capacity is involved in the endogenous preparation (i.e., maintenance, selection, and implementation) of task orders, we should find an impaired performance (i.e., increased RTs, error rates, and/or response reversal rates) in the present systematic-order blocks under high in contrast to low-WM-load conditions. In addition, if WM capacity is specifically involved when endogenous preparation reconfigure a new task order, we should specifically find an impaired performance in the order-switch trials under high-WM-load conditions. To investigate the impact of task complexity, we realized visual and auditory tasks with sets of two stimulus–response mappings (low difficulty) and sets of three stimulus–response mappings (high difficulty); we assume that an increased number of stimulus–response mappings require increased WM capacity. If endogenous preparation and the reconfiguration of a new task order (i.e., order-switch trials) is affected by this complexity manipulation in WM, order-switch costs should be increased under high- in comparison to the low-complexity conditions. General dual-task performance differences between high- and low-complexity conditions cannot be interpreted in the context of endogenous task-order preparation, since these differences simply reflect complexity differences within the dual tasks.
Methods
Stimuli, procedure, and design were identical to the general methods described above, with the following specifications. The dual-task test of Experiment 2 included 32 dual-task blocks with systematic order exclusively; each block had 18 trials. These blocks were realized with an ITI of 700 ms, and 2 levels of WM load. High WM load included the presentation of 4 digits before the start of the dual-task blocks and we instructed the maintenance of these digits across the dual-task blocks, while low load included the presentation of 1 digit before dual-task blocks and its maintenance across blocks. The order of high-WM-load blocks (16 blocks) and low-WM-load blocks (16 blocks) was alternating and half of the participants started with high-load blocks, while the other half started with low-load blocks. Under both WM load conditions, participants reported the maintained information (i.e., 4 digits and 1 digit under high- and low-load conditions, respectively) to the experimenter after the individual dual-task blocks. This manipulation of WM load was realized under 2 conditions of stimulus–response difficulty: two stimulus–response mappings under the low difficulty condition and three stimulus–response mappings under the high difficulty condition (see General methods). Thirty-two and 32 participants conducted the experiment versions with low task complexity (23 females, mean age: 22.5 years, age range: 19–27 years) and high task complexity (22 females, mean age: 24.0 years, age range: 20–28 years), respectively.
Results
Before analyzing, we excluded those participants with total error rates of more than 25% across dual-task blocks. As a consequence, we included 20 and 22 participants in the experiment versions with two and three stimulus–response mappings, respectively. Further, to check our WM manipulation, we analyzed the number of correct responses in the WM task (maximum 16 correct responses) in a mixed measures ANOVA with the within-subjects factor WM LOAD (low load vs. high load) and the between-subjects factor TASK COMPLEXITY (low complexity vs. high complexity). TASK COMPLEXITY showed a significant effect, F(1, 40) = 8.256, p < .01, ŋp2 = .17, resulting from more correct responses under the low-complexity condition (M = 15.9) in comparison to the number of correct responses under the high-complexity condition (M = 15.6). Importantly, there were significantly more correct responses with low WM load (M = 15.8) in contrast to the number of these responses with high WM load (M = 15.6), F(1, 40) = 5.253, p < .05, ŋp2 = .12. The factors did not interact, F(1, 40) = 1.870, p = .18. Thus, this analysis provided evidence that the different task complexity levels as well as the different load levels of our WM task resulted in different performance levels in this task which is a necessary precondition to investigate its effect on dual-task performance.
RTs (after the exclusion of trials with errors on the choice decisions within the component tasks and response reversals), error rates, and response reversal rates (after the exclusion of trials with errors on the choice decisions within the component tasks) were aggregated across visual–auditory and auditory–visual order trials in dual-task blocks with a correct response in the WM task. We included the aggregated RT, error, and response reversal data of the trials with same task order and different task orders across blocks with low and high WM load. Task 1 (firstly presented task) and Task 2 (secondly presented task) performance was separately analyzed in mixed measures ANOVAs including the within-subjects factors TRIALTYPE (same order vs. different order) and WM LOAD (low load vs. high load) as well as the between-subjects factor TASK COMPLEXITY (low complexity vs. high complexity).
RTs (Fig. 4) Analyzing RT1, the main effect of TRIALTYPE was significant, F(1, 40) = 47.977, p < .001, ŋp2 = .54, generally replicating findings of order-switch costs in previous experiments (e.g., Experiment 1) and studies (e.g., Kübler et al., 2018; Szameitat et al., 2006); same-order trials showed lower RTs (M = 1177 ms) than different-order trials (M = 1276 ms). Further, while WM LOAD showed no significant effect, F(1, 40) < 1, TASK COMPLEXITY was also significant, F(1, 40) = 11.816, p < .001, ŋp2 = .23, demonstrating faster RTs under conditions of low complexity (M = 1091 ms) in comparison to high complexity (M = 1363 ms). The RT1 analysis showed no interaction, Fs < 3.493, ps > .07. Thus, this analysis provided no convincing evidence that task complexity and WM load hampered general endogenous preparation of task order in DT as well as specific endogenous preparation of order switches.

Reaction times (RTs) in ms in Experiment 2 across the groups of participants with low and high task complexity, in blocks with low and high working-memory (WM) load as well as in trials with repetitions of task order (same) and switches between different task orders (diff.). a RT1. b RT2. Error bars represent standard errors of the mean
Analyzing RT2 similar to RT1, the main effect of TRIALTYPE was significant, F(1, 40) = 55.651, p < .001, ŋp2 = .58, consistent with previous experiments (Experiment 1) and studies (e.g., Kübler et al., 2018; Szameitat et al., 2006): same-order trials showed lower RTs (M = 1269 ms) than different-order trials (M = 1381 ms). Furthermore, TASK COMPLEXITY was also significant, F(1, 40) = 11.309, p < .001, ŋp2 = .22, demonstrating faster RTs under conditions of low complexity (M = 1187 ms) in comparison to high complexity (M = 1463 ms). Similar to RT1, the present analysis showed no significant interactions, Fs < 3.181, ps > .08. Thus, there is no substantial evidence that manipulations of WM capacity (i.e., task complexity and WM load) modulated endogenous order preparation. The main effect of WM LOAD was also not significant, F(1, 40) < 1.
Errors (Table 2) The error analysis of the first task was equivalent to the RT analysis and revealed a main effect for TASK COMPLEXITY, F(1, 40) = 36.032, p < .001, ŋp2 = .47. Error rates were lower under the low-complexity condition (M = 1.6%) than under the high-complexity condition (M = 6.5%). All other main effects, Fs < 3.777, ps > .06, as well as interactions, Fs < 1.109, ps > .30, were non-significant.
The error analysis of the second task also revealed a main effect or TASK COMPLEXITY, F(1, 40) = 45.763, p < .001, ŋp2 = .54; error rates were lower under the low-complexity condition (M = 3.7%) compared to high-complexity condition (M = 11.1%). The remaining factors showed no main effects, Fs < 1, and did not interact, Fs < 3.467, ps > .07.
Response reversal rates (Fig. 5) The analyses of the response reversal rates revealed main effects for TRIALTYPE, F(1, 40) = 10.343, p < .01, ŋp2 = .21, and WM LOAD, F(1, 40) = 7.708, p < .01, ŋp2 = .16. The response reversal rates were significantly reduced in same-order trials (M = 4.5%) than in different-order trials (M = 6.1%); replicating the findings of Experiment 1. These rates were also reduced under low-load conditions (M = 4.7%) in comparison to high-load conditions (M = 5.9%). However, this impact of the manipulation of WM capacity on task performance was limited to only a single condition, F(1, 40) = 4.417, p < .05, ŋp2 = .10. While WM load had an impact under the condition of high task complexity and same-order trials (p < .01), this load had no impact under the remaining conditions (high task complexity–different order, low task complexity–same order, low task complexity–different order) (ps > .19). Since the impact of WM load is rather limited in the response reversal data and this impact is not corroborated by the RT and error data, we are careful with interpretations about the impact of WM load and exogenous task-order preparation. The remaining main effect and factor combinations were not significant, Fs(1, 40) < 2.708, ps > .11.

Response reversal rates (in percent) in Experiment 2 across the groups of participants in blocks with low and high working-memory (WM) load as well as in trials with repetitions of task order (same) and switches between different task orders (diff.). a Low task complexity. b High task complexity. Error bars represent standard errors of the mean
Discussion
The present experiment aimed to further specify the processing characteristics of endogenous task-order preparation and the involvement of WM capacity in this preparation. Although it cannot be excluded that there was a lack of statistical power resulting in a beta-error despite the decent sample size, the present data did not demonstrate a significant impact of the present manipulations of WM load on the general dual-task performance with varying task order as well as the specific order-switch costs. Further, the task complexity manipulation revealed no effect on these costs as well. Thus, these data provided no reliable evidence that the WM component is involved in the endogenous preparation of task order. (Note that the only impact of WM load on dual-task performance emerged under the conditions of high task complexity and same-order trials and this impact was limited to the response reversal data, but was not evident in the RT and error data.)
There are several options to explain these results. First, although we found an impact of WM load on the performance in the WM task and an impact of task complexity on this task as well as on the dual task, it might still be possible that these manipulations were not effective to tap the limits of the subjects’ WM capacity in a way that affects preparation of task order. In essence, there might be still capacity left for an efficient preparation because of an incomplete exhaustion of available WM capacity across individual subjects. Assessing individual differences in WM capacity would, thus, be an idea of investigating the relation between WM and task-order preparation in future studies. Second, the present manipulations rather required the maintenance of information in WM, i.e., the maintenance of one or four digits as well as the maintenance of two-choice or three-choice component tasks. However, it might be that at least the information of the WM task are represented in a component that differs from the component related to the processing of dual tasks (Baddeley, 2012). While the digits of the WM task are verbal information, the dual task includes task information that is convertible into spatial and procedural information. The lacking impact of our manipulation of the WM task on the dual task in general is consistent with this assumption. This lacking impact is also consistent with the assumption that response selection processes within the component tasks are not only related with domain-general processes, but also they engage domain-specific WM processes (Hazeltine & Wifall, 2011). That is potential interference between the WM task and the dual task might not occur because of the different WM load conditions, but because of the manual responses in the dual task. However, this latter task characteristic (i.e., manual responses) is held constant across the different WM load conditions and we would not assume varying dual-task performances across these conditions. Future studies could investigate this assumption using different response modalities between different dual tasks and consider individual differences as a further factor moderating the relation between WM and dual-task processing.
Third, it might be that endogenous preparation of order switches and thus, executive control is not related to the maintenance of WM content, but rather to building and updating processes on this content (Oberauer, 2009; Stelzel, Kraft, Brandt, & Schubert, 2008). This assumption is still consistent with the assumption that endogenous order preparation is associated with WM. However, this preparation is rather associated with WM updating processes, than with maintenance of information in WM. These options require further investigations in future studies.
Experiment 3
The previous experiments demonstrated, as did Szameitat et al. (2006), that there is a substantial component of the order-switch cost that cannot be eliminated by allowing participants appropriate time to endogenously prepare for the task-order switch (e.g., up to 2500 ms in Experiment 1). There are three hypotheses concerning this residual component of order-switch costs and their predictions for what should be observed when we extend the sequence of trials between task-order switches beyond two, as we did in this Experiment 3.
It could be that the costs of task-order switching arise because of interference exerted proactively by the residue of the previous task order. The time costs of switching order cannot be understood as the reflection of a discrete stage that must be completed before the start of the next dual-task trial. Rather, these costs may represent the additional time needed for the cognitive control to settle to a unique task order (or first component task) after the trial started. Such a settlement would be equivalent to phenomena in the context of task switching that show persistent adaptation of cognitive control over several trials (Alport, Styles, & Hsieh, 1994). A crucial feature of this residue order activation hypothesis is that the residue activation responsible for this additional time might have persistence on the order of several following trials. Hence, if we increase the length of the runs in systematic dual-task blocks, not only the first trial following a task-order switch, but also a number of trials thereafter, should still show the effects of this residue activation. As a consequence, performance should not only be improved in trials with the first repetition of a task order in contrast to trials with task-order switches, but there should be further improvements after the first repetition in following trials, i.e., order-switch costs decrease with an increase in the number of order repetitions.
A similar prediction can be derived from dynamic processes of optimization that happen on a time scale of individual trials. For example, over a series of trials, RTs reduce steadily until the participant makes an error, lengthens abruptly on the posterror trial, and then improves steadily again until another error is made (a phenomenon called post-error slowing; Dutilh et al., 2012; Rabbitt & Vyas, 1970). This performance pattern implies a dynamic tracking process for staying close to the optimum speed–accuracy trade-off. It also seems likely that order-specific processing is adjusted dynamically from trial to trial. This option explains order-switch costs by assuming that the participant embarks on each new sequence of trials and it takes a few trials to re-optimize performance on the basis of trial-by-trial feedback; such a re-optimization might be one form of endogenous task-order preparation. Like the hypothesis of residue order activation, this optimization hypothesis appears to predict that the order-switch costs should take more than a single trial with the new order to dissipate.
The completion hypothesis proposes that an endogenous act of executive control deployed before the onset of the dual-task stimuli can achieve only part of the process of task-order reconfiguration. Completion of this reconfiguration is triggered by the presentation of these stimuli and their order. In detail, moment-by-moment reorganizations of processing resources to accomplish different tasks and different task orders are achieved by the combined and often sequential operation of anticipatory endogenous and stimulus-triggered exogenous executive control mechanisms. Luria and Meiran (2003, 2006) proposed a similar interplay of top–down mechanisms (such as the endogenous preparation of task order) and bottom–up mechanisms (such as the presentation order of the task stimuli). This hypothesis makes a strong prediction different from that made by either the residue order activation hypothesis or the optimization hypothesis. After the stimulus-cued completion of task-order reconfiguration has been triggered once by dual-task stimuli, this reconfiguration should not be required again while the task order remains unchanged. Hence the cost of switching between orders should not be further reducible after the first trial with an order repetition. Experiment 3 was a test of this prediction on order-switch costs. Instead of switching between different task orders after sequences of only two trials as in the previous experiments, participants switched order, still completely predictably, after sequences of four trials in each order.
Methods
Stimuli, procedure, and design were identical to the general methods described above, with the following specifications. Experiment 3’s dual-task test was performed with 10 dual-task blocks with systematic orders exclusively. Each block included 36 trials; we selected this increased number of trials per block to have a sufficient number of trials under each trial condition (i.e., order switch, first order repetition, second order repetition, and third order repetition). This is because, in contrast to the previous experiments, task order switched every 4 trials, resulting in sequences of 4 trials with the same task order. ITI was 700 ms across all blocks. Twenty participants conducted this experiment (12 females, mean age: 22.6 years, age range: 19–25 years).
Results
Identical to the previous experiments, we excluded those participants with total error rates of more than 25% across the dual-task blocks before analyzing RTs and error rates. As a consequence, we included the data of 16 participants in this experiment. Similar to the previous experiments, RTs (after the exclusion of trials with errors on the choice decisions within the component tasks and response reversals), error rates, and response reversal rates (after the exclusion of trials with errors on the choice decisions within the component tasks) were aggregated across visual–auditory and auditory–visual order trials in the dual-task blocks.
We included the RT, error, and reversal data of the trials with different task order as well as the first, second, and third repetition of the same task order (same order 1, same order 2, and same order 3, respectively). Task 1 (firstly presented task) and Task 2 (secondly presented task) performance was separately analyzed in repeated measures ANOVAs including the within-subjects factor TRIALTYPE (different order, same order 1, same order 2, and same order 3).
RTs (Fig. 6) Analyzing RT1, the main effect of TRIALTYPE was significant, F(3, 42) = 21.113, p < .001, ŋp2 = .60, generally replicating findings of order-switch costs in previous experiments (Experiment 1 and 2) and studies (e.g., Kübler et al., 2018; Szameitat et al., 2006). That is, different-order trials showed higher RTs (M = 1299 ms) than same-order trials with the first repetition (M = 1120 ms, p < .001), with the second repetition (M = 1145 ms, p < .001), and with the third repetition (M = 1179 ms, p < .001). Repetitions of the same task order did not improve task-order control, since RTs of the second and third order repetitions were not below RTs of the first repetition. Instead, RTs of the second repetition and third repetition did not differ (p = .19) as well as significantly above (uncorrected ps < .05) the first repetition’s RTs, respectively. (We will come back to this latter finding in the discussion.)

Reaction times (RTs) in ms in Experiment 3 in trials with switches between different task orders (diff.), the first repetitions of the same task order (same 1), the second repetitions of the same task order (same 2) as well as the third repetitions of the same task order (same 3). a RT1. b RT2. Error bars represent standard errors of the mean
Analyzing RT2 similar to RT1, the main effect of TRIALTYPE was significant, F(3, 42) = 16.414, p < .001, ŋp2 = .54, consistent with previous experiments (Experiment 1 and 2) and studies (e.g., Kübler et al., 2018; Szameitat et al., 2006). In detail, different-order trials showed higher RTs (M = 1414 ms) than same-order trials with the first repetition (M = 1245 ms, p < .001), with the second repetition (M = 1273 ms, p < .001), and with the third repetition (M = 1301 ms, p < .001). Similar to the conclusions on RT1, repetitions of the same task order did not improve task-order control, since RTs of the second and third order repetitions were not below RTs of the first repetition. Instead, RTs of the second repetition and third repetition did not differ (p = .17) as well as significantly above (uncorrected ps < .05) the first repetition’s RTs, respectively.
Errors (Table 3) The error analysis of the first task was equivalent to the RT analyses and revealed a main effect for TRIALTYPE, F(3, 42) = 12.780, p < .001, ŋp2 = .47. In detail, different-order trials showed higher error rates (M = 14.2%) than same-order trials with the first repetition (M = 7.2%, p < .01), with the second repetition (M = 4.7%, p < .01), and with the third repetition (M = 10.9%, p < .05). Consistent with the RTs, repetitions of the same task order did not improve task-order control consistently, since error rates of at least the third order repetitions were not below these rates of the first and the second repetitions. Instead, error rates of the third repetitions were significantly above the error rates of the first and the second repetitions (uncorrected ps < .05).
The error analysis of the second task revealed a main effect for TRIALTYPE, F(3, 42) = 9.244, p < .001, ŋp2 = .40; different-order trials showed higher error rates (M = 15.6%) than same-order trials with the first repetition (M = 8.9%, p < .01), with the second repetition (M = 8.7%, p < .01), and with the third repetition (M = 12.2%, p < .05). Similar to the previous analyses, repetitions of the same task order did not improve task-order control, since error rates of the third order repetition were not below these rates of the first and the second repetitions. Instead, error rates of the first and second repetitions did not differ (p = .85) and significantly below (uncorrected ps < .05) the third repetition’s error rates.
Response reversal rates (Fig. 7) The analyses of the response reversal rates revealed a main effect for TRIALTYPE, F(3, 42) = 11.858, p < .001, ŋp2 = .46. The response reversal rates were significantly increased in different-order trials compared to all conditions of order-swetitions (ps < .05). Similar to the previous analyses, repetitions of the same task order did not improve task-order control, since response reversal rates of the third order repetition were not below these rates of the first and the second repetitions. Instead, response reversal rates of the first and second repetitions did not differ (p = .50) and significantly below (ps < .05) the third repetition’s rates.

Response reversal rates (in percent) in Experiment 3 in trials with switches between different task orders (diff.), the first repetitions of the same task order (same 1), the second repetitions of the same task order (same 2) as well as the third repetitions of the same task order (same 3). Error bars represent standard errors of the mean
Discussion
The present experiment tested the predictions of the residue order activation hypothesis, the optimization hypothesis, and the completion hypothesis. In essence, our results are largely consistent with the assumptions of the latter hypothesis: the cost of switching between orders was not further reduced after the first trial with an order repetition. Thus, after the stimulus-cued completion of task-order reconfiguration has been triggered once by the dual-task stimuli, this reconfiguration appears not to be required again while the task order remains unchanged. However, it has to be noted at this point that the present data do not fully rule out the residue order activation and the optimization hypothesis, since a further reduction of RTs over task-order repetitions might be masked by anticipatory task-order preparation on the third order repetition and this interpretation is partly based on a non-significant difference between trials with a first and a second order repetition.
However, is there evidence for endogenous preparation of task order at all in this set of data? We assume that the increase of RTs, error rates, and response reversal rates from the first to the last repetition of the same task-order and thus within-sequence slowing reflects components of order preparation that are not exogenously stimulus triggered. If there is exogenous preparation exclusively, we would not expect such a RT slowing and error rate increase. Instead, there are two not mutual exclusive explanations consistent with endogenous task-order preparation. First, it might be that participants expect the end of the current sequence of the same task order and prepare the switch to another order. This preparation initiates the upcoming alternative, but currently irrelevant task order, leading to impairment during progressive sequence positions. Alternatively, it might be that RT slowing and error rate increase reflect a further demand of endogenous task-order preparation under the systematic-order condition. Namely, participants need to monitor task progression and keep the task sequence in mind (i.e., basically count trials) to know when an order switch will occur; this counting is a consequence of not having an explicit cue in the present experiment. However, at the end of the sequences, participants might have lost track when counting and endogenous preparation is less reliable. In the context of both explanations, however, endogenous preparation actually impairs dual-task performance. However, since this impairment is not foreshadowed by the dual-task stimuli, we interpret this data pattern as evidence for the endogenous preparation component and this component is responsible for within-sequence slowing.
These interpretations are also rather inconsistent with an alternative interpretation of the findings in Experiment 1. In this experiment, we interpreted the reduction of order-switch costs with prolonged ITIs and predictable switches in the error rates and response reversals as evidence for the existence of endogenous task-order preparation. However, it could also be that the activation of task-order information merely decays with prolonged ITIs, leading to the reduction of task-order switch costs. In such a case, we would expect a performance improvement with an increase in repetitions of the same task order in the present experiment. Weak evidence against the decay explanation as an alternative explanation to endogenous task-order preparation comes, however, from the constant performance between the first and the second task-order repetition and generally the lacking improvements of performance across repetitions of the same task order in Experiment 3. In fact, there is rather a performance impairment since the last appearance of an alternative order; this impairment could be based on anticipatory preparation effects especially since it was predictable for the participants when the next task-order switch was coming. Thus, since we did not observe a performance improvement, the present data are inconsistent with this decay alternative in Experiment 1.
General discussion
The aim of the present study was to provide evidence for internally driven endogenous control and preparation of dual-task orders that regulate the access to a bottleneck stage. To operationalize endogenous preparation, we presented dual-task blocks with varying task orders and informed participants in advance about repetitions of the same task order (i.e., task-order repetitions) as well as switches between different task orders (i.e., task-order switches); thus task-order repetitions and switches were predictable. Experiment 1 demonstrated that task-order information and an increased preparation time generally increases the efficiency for endogenous task-order control and improves preparation for task-order switches. This finding is basically consistent with the assumption of the existence of endogenous control processes. Experiment 2, however, did not provide conclusive evidence that this endogenous control is related with WM maintenance mechanisms. Experiment 3 showed that endogenous control does not only fully complete task-order preparation but also requires exogenous, stimulus-driven components.
Relations between the present and previous findings in the dual-task literature
Consistent with numerous studies in the dual-task literature applying variations of the PRP dual-task type is that there is control of dual tasks with different task orders and thus control of the access to a potential bottleneck stage (e.g., Hendrich et al., 2012; Hirsch et al., 2017; Ruiz Fernández et al., 2011; Sigman & Dehaene, 2006; Strobach et al., 2018; Szameitat et al., 2006; Töllner et al., 2012). However, some of these studies assumed that bottleneck processing is simply recruited by the stimuli on an exogenous first-come-first-served basis, so that the order in which the tasks are handled is determined by which of the two task stimuli arrives at the bottleneck first (e.g., Hendrich et al., 2012; Kübler et al., 2018; Leonhard et al., 2011; Ruiz Fernández et al., 2011; Sigman & Dehaene, 2006; Strobach et al., 2018). Alternative to this exogenous model, the endogenous model assumes that executive control processes actively coordinate and prepare access to the bottleneck and thus task order. However, previous studies testing this model applied cues indicating the first task and, thus, task order (de Jong, 1995; Luria & Meiran, 2003); in the introduction, we discussed potential issues with this method investigating the endogenous model. Therefore, we provided information about the sequence of task order over successive trials and made task order predictable. Consistent with the previous findings, the present findings supported the assumption of endogenous task-order control and preparation when applying this new experimental method (i.e., provide information about task order before the start of the experimental blocks).
As a consequence, we assume that bottleneck processing models require not only assumptions about the characteristics of processing stages within the component tasks (e.g., bottlenecks at response selection stages), but also assumptions about executive control processing during dual-task processing (e.g., Szameitat et al., 2006). For instance, Szameitat, Schubert, and colleagues (Szameitat et al., 2006; Szameitat, Schubert, Müller, & von Cramon, 2002) suggested that the flow of information from perceptual channels into a central response selection processor is controlled by an endogenous gating mechanism, regulating the order in which the perceptual information of the two tasks is further handled in the putative central processor. According to the endogenous model, the order of processing is explicitly controlled by this gating mechanism by initially allocating the central channel to the task to be performed first (and re-allocating it to the other task after the first task is done with it). Furthermore, endogenous task-order preparation might require monitoring the task progression to keep the task sequence in mind to know when an order switch will occur (see discussion of Experiment 3) and this preparation might be realized by information activating higher-level task representations that includes information about task order (Hirsch et al., 2017; Hirsch, Nolden, Philipp, & Koch, 2018; Lien & Ruthruff, 2004). However, the present experimental design does not allow specifying whether this higher-level task representation includes the relevant task-order information exclusively (Hübner, Futterer, & Steinhauser, 2001) or include the relevant task-order information plus specific information about the component tasks (Philipp & Koch, 2010). Future studies are required to differentiate between these two possibilities. Such study should also characterize the nature of the capacity limitation in situation with the potential for endogenous task-order preparation. So far, we mainly discussed this issue from the perspective of central bottleneck theories and structural bottlenecks (Pashler, 1994; Sigman & Dehaene, 2006). However, when focussing on resource sharing models, endogenous task-order preparation might not define a particular task order, but this preparation regulates the amount of limited processing capacity distributed between simultaneous component tasks based on demands of the present dual task with different orders (Miller et al., 2009).
The findings of the present Experiment 2 provided no convincing evidence that the task-order preparation involves the same WM component that is involved in the maintenance of digits and is not modulated by the number of relevant stimulus–response mappings. In the discussion of Experiment 2, we discussed numerous options that might explain the lack of evidence for the relation between WM capacity and task-order preparation. This lacking evidence is inconsistent with assumptions of other dual-task practice studies. These studies assumed that the practice-related acquisition of task coordination skills leads to an improved task control and this control is at least partially associated with the working-memory component (Stelzel et al., 2008). For instance, dual-task control is improved with practice because of an efficient and conjoint instantiation of relevant information of two component tasks in working memory at the onset of a dual-task trial (Maquestiaux et al., 2004; Schubert & Strobach, 2018; Strobach, Salminen, Karbach, & Schubert, 2014). The present study, however, provided no evidence for an involvement of the working-memory component in predictable task-order control.
Relations between the present dual-task findings and previous findings in the task-switching literature
As foreshadowed in the introduction, endogenous task control was investigated not only in the context of dual tasks, but also in the task-switching literature. For instance, in the task-cuing procedure, the two tasks are presented in random order and participants are informed about the identity of the upcoming task by a cue that precedes the presentation of the target stimulus (Meiran, 1996) and switching between tasks incurs task-switch costs. Because participants are informed about the identity of the upcoming task by the task cue, a manipulation of the length of the cue-target interval (CTI) produces different amounts of processing times for task-specific preparation. Performance usually benefits from an increase of the CTI more so on task-switch trials in comparison to repetition trials, resulting in a reduction of the switch costs (Meiran, 1996). This reduction at long CTIs in contrast to short CTIs has been referred to as the Reduction In Switch Cost (RISC) effect (Liefooghe, Demanet, & Vandierendonck, 2009). The RISC effect in the task-cuing procedure has been accounted for more effective endogenous task preparation in task-switch trials, suggesting some form of advanced task-set reconfiguration during CTIs. Such an advanced reconfiguration is not necessary in task-repetition trials, reducing the impact of CTIs on task repetition performance (Kiesel et al., 2010; Rogers & Monsell, 1995). One option to explain this effect is to assume improved preparatory attentional weighting of perceptual dimensions (i.e., biasing processing towards the target stimulus dimension of the upcoming task; Lien, Ruthruff, & Johnston, 2010; Meiran, 2000; Müller, Reimann, & Krummenacher, 2003) with longer CTIs and task switches. With respect to the present study, both the dual-task and the task-switching literature discuss endogenous task-control processes (Monsell & Mizon, 2006) (however, see Logan, Schneider, & Bundesen, 2007).
The findings of the present Experiment 3 showed that endogenous task-order control does not only fully complete task-order preparation but also requires exogenous, stimulus-driven components. On some level, this conclusion is consistent with explanations of the phenomenon of residual task-switching costs in the task-switching literature. Task-switching costs remain even after the preparation time has been extended to multiples of the switch cost itself (e.g., 5 s), suggesting that there are sources of persisting interference that cannot be overcome by advance preparation for a task switch (e.g., Monsell, 2003). The residual task-switching costs seem to imply that there are structural limits of task-switching preparation (de Jong, 2000). Two-stage models of reconfiguration proposed that a first stage including “advance reconfiguration” (Rogers & Monsell, 1995) or “goal shifting” (Rubinstein et al., 2001) can be completed prior to target onset and thus represents cognitive flexibility. At a second stage “stimulus-cued completion” or “rule activation” can occur only when being triggered by the actual task stimulus and thus represents a structural limitation for final task readiness. This assumption of a structural limitation is consistent with assumptions of a structural limitation to prepare predictable task orders in the present Experiment 3.
In sum, the present findings are basically consistent with the assumption of the existence of endogenous control processes. These processes are, however, not related with working-memory maintenance mechanisms. Further, endogenous control does not only fully complete task-order preparation but also requires exogenous, stimulus-driven components.
References
Alport, A., Styles, E. A., & Hsieh, S. (1994). Shifting Intentional Set: Exploring the Dynamic Control of Tasks. In C. Umilta & M. Moscovitch (Eds.), Attention and performance XV (pp. 421–452). Cambridge, MA: MIT Press.
Baddeley, A. (2012). Working memory: Theories, models, and controversies. Annual Review of Psychology, 63, 1–29.
Band, G. P. H., & van Nes, F. T. (2006). Reconfiguration and the bottleneck: Does task switching affect the refractory period effect? European Journal of Cognitive Psychology, 18(4), 593–623.
Braver, T. S., Cohen, J. D., Nystrom, L. E., Jonides, J., Smith, E. E., & Noll, D. C. (1997). A parametric study of prefrontal cortex involvement in human working memory. Neuroimage, 5(1), 49–62.
de Jong, R. (1995). The role of preparation in overlapping-task performance. The Quarterly Journal of Experimental Psychology A: Human Experimental Psychology, 48A(1), 2–25.
de Jong, R. (2000). An intention-activation account of residual switch costs. In S. Monsell & J. Driver (Eds.), Control of cognitive processes: Attention and performance XVIII (pp. 357–376). Cambridge, MA: MIT Press.
Dreisbach, G., & Haider, H. (2009). How task representations guide attention: Further evidence for the shielding function of task sets. Journal of Experimental Psychology: Learning, Memory, and Cognition, 35(2), 477–486.
Dutilh, G., Vandekerckhove, J., Forstmann, B. U., Keuleers, E., Brysbaert, M., & Wagenmakers, E.-J. (2012). Testing theories of post-error slowing. Attention, Perception, & Psychophysics, 74(2), 454–465.
Hartley, A. A., & Little, D. M. (1999). Age-related differences and similarities in dual-task interference. Journal of Experimental Psychology: General, 128(4), 416–449.
Hazeltine, E., Ruthruff, E., & Remington, R. W. (2006). The role of input and output modality pairings in dual-task performance: Evidence for content-dependent central interference. Cognitive Psychology, 52(4), 291–345.
Hazeltine, E., & Wifall, T. (2011). Searching working memory for the source of dual-task costs. Psychological Research, 75(6), 466–475.
Hendrich, E., Strobach, T., Buss, M., Müller, H. J., & Schubert, T. (2012). Temporal-order judgment of visual and auditory stimuli: Modulations in situations with and without stimulus discrimination. Frontiers in Integrative Neuroscience, 6, 63.
Hirsch, P., Nolden, S., & Koch, I. (2017). Higher-order cognitive control in dual tasks: Evidence from task-pair switching. Journal of Experimental Psychology: Human Perception and Performance, 43(3), 569–580.
Hirsch, P., Nolden, S., Philipp, A. M., & Koch, I. (2018). Hierarchical task organization in dual tasks: Evidence for higher level task representations. Psychological Research, 82(4), 759–770.
Hübner, R., Futterer, T., & Steinhauser, M. (2001). On attentional control as a source of residual shift costs: Evidence from two-component task shifts. Journal of Experimental Psychology. Learning, Memory, and Cognition, 27(3), 640–653.
Huestegge, L., & Koch, I. (2009). Crossmodal action selection: Evidence from dual-task compatibility. Memory & Cognition, 38(4), 493–501.
Kahneman, D. (1973). Attention and effort. Englewood Cliffs, NJ: Prentice-Hall.
Kane, M. J., & Engle, R. W. (2003). Working-memory capacity and the control of attention: The contributions of goal neglect, response competition, and task set to Stroop interference. Journal of Experimental Psychology: General, 132(1), 47–70.
Karbach, J., & Kray, J. (2009). How useful is executive control training? Age differences in near and far transfer of task-switching training. Developmental science, 12(6), 978–990.
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K., Philipp, A. M., & Koch, I. (2010). Control and interference in task switching—A review. Psychological Bulletin, 136(5), 849–874.
Koch, I., Poljac, E., Müller, H., & Kiesel, A. (2018). Cognitive structure, flexibility, and plasticity in human multitasking—An integrative review of dual-task and task-switching research. Psychological Bulletin, 144(6), 557–583.
Kübler, S., Reimer, C. B., Strobach, T., & Schubert, T. (2018). The impact of free-order and sequential-order instructions on task-order regulation in dual tasks. Psychological Research, 82(1), 40–53.
Leonhard, T., Ruiz Fernández, S., Ulrich, R., & Miller, J. (2011). Dual-task processing when task 1 is hard and task 2 is easy: Reversed central processing order? Journal of Experimental Psychology: Human Perception and Performance, 37(1), 115.
Liefooghe, B., Demanet, J., & Vandierendonck, A. (2009). Is advance reconfiguration in voluntary task switching affected by the design employed? The Quarterly Journal of Experimental Psychology, 62(5), 850–857.
Lien, M.-C., & Ruthruff, E. (2004). Task switching in a hierarchical task structure: Evidence for the fragility of the task repetition benefit. Journal of Experimental Psychology: Learning, Memory, and Cognition, 30(3), 697–713.
Lien, M.-C., Ruthruff, E., & Johnston, J. C. (2010). Attentional capture with rapidly changing attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 36(1), 1–16.
Logan, G. D. (2003). Executive control of thought and action: In search of the wild homunculus. Current Directions in Psychological Science, 12(2), 45–48.
Logan, G. D., & Bundesen, C. (2003). Clever homunculus: Is there an endogenous act of control in the explicit task-cuing procedure? Journal of Experimental Psychology: Human Perception and Performance, 29(3), 575–599.
Logan, G. D., Schneider, D. W., & Bundesen, C. (2007). Still clever after all these years: Searching for the homunculus in explicitly cued task switching. Journal of Experimental Psychology: Human Perception and Performance, 33(4), 978–994.
Luria, R., & Meiran, N. (2003). Online order control in the psychological refractory period paradigm. Journal of Experimental Psychology: Human Perception and Performance, 29(3), 556–574.
Luria, R., & Meiran, N. (2006). Dual route for subtask order control: Evidence from the psychological refractory paradigm. The Quarterly Journal of Experimental Psychology, 59(4), 720–744.
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S. (2000). Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control. Science, 288(5472), 1835–1838.
Maquestiaux, F., Hartley, A. A., & Bertsch, J. (2004). Can practice overcome age-related differences in the psychological refractory period effect? Psychology and Aging, 19(4), 649–667.
Mayr, U., & Kliegl, R. (2003). Differential effects of cue changes and task changes on task-set selection costs. Journal of Experimental Psychology. Learning, Memory, and Cognition, 29(3), 362–372.
Meiran, N. (1996). Reconfiguration of processing mode prior to task performance. Journal of Experimental Psychology. Learning, Memory, and Cognition, 22(6), 1423–1442.
Meiran, N. (2000). Modeling cognitive control in task-switching. Psychological Research, 63(3–4), 234–249.
Meyer, D. E., & Kieras, D. E. (1997). A computational theory of executive cognitive processes and multiple-task performance: Part 2. Accounts of psychological refractory-period phenomena. Psychological Review, 104(4), 749–791.
Miller, J., & Durst, M. A. (2015). Comparison of the psychological refractory period and prioritized processing paradigms: Can the response-selection bottleneck model explain them both? Journal of Experimental Psychology: Human Perception and Performance, 41(5), 1420–1441.
Miller, J., Ulrich, R., & Rolke, B. (2009). On the optimality of serial and parallel processing in the psychological refractory period paradigm: Effects of the distribution of stimulus onset asynchronies. Cognitive Psychology, 58(3), 273–310.
Mittelstädt, V., & Miller, J. (2017). Separating limits on preparation versus online processing paradigms: Evidence for resource models. Journal of Experimental Psychology: Human Perception and Performance, 43(1), 89–102.
Monsell, S. (2003). Task switching. Trends in Cognitive Sciences, 7(3), 134–140.
Monsell, S., & Mizon, G. A. (2006). Can the task-cuing paradigm measure an endogenous task-set reconfiguration process? Journal of Experimental Psychology: Human Perception and Performance, 32(3), 493–516.
Müller, H. J., Reimann, B., & Krummenacher, J. (2003). Visual search for singleton feature targets across dimensions: Stimulus-and expectancy-driven effects in dimensional weighting. Journal of Experimental Psychology: Human Perception and Performance, 29(5), 1021–1035.
Norman, D. A., & Shallice, T. (1986). Attention to action: Willed and automatic control of behavior. In R. Davidson, G. Schwartz, & D. Shapiro (Eds.), Consciousness and self regulation: Advances in research and theory (Vol. 4, pp. 1–18). New York, NY: Plenum.
Oberauer, K. (2009). Interference between storage and processing in working memory: Feature overwriting, not similarity-based competition. Memory & Cognition, 37(3), 346–357.
Pashler, H. (1994). Dual-task interference in simple tasks: Data and theory. Psychological Bulletin, 116(2), 220–244.
Philipp, A., & Koch, I. (2010). The integration of task-set components into cognitive task representations. Psychologica Belgica, 50, 383–411.
Rabbitt, P. M. A., & Vyas, S. M. (1970). An elementary preliminary taxonomy for some errors in laboratory choice RT tasks. Acta Psychologica, 33, 56–76.
Rogers, R. D., & Monsell, S. (1995). Costs of a predictible switch between simple cognitive tasks. Journal of Experimental Psychology: General, 124(2), 207–231.
Rubinstein, J. S., Meyer, D. E., & Evans, J. E. (2001). Executive control of cognitive processes in task switching. Journal of Experimental Psychology: Human Perception and Performance, 27(4), 763–797.
Ruiz Fernández, S., Leonhard, T., Rolke, B., & Ulrich, R. (2011). Processing two tasks with varying task order: Central stage duration influences central processing order. Acta Psychologica, 137(1), 10–17.
Schneider, D. W., & Logan, G. D. (2007). Task switching versus cue switching: Using transition cuing to disentangle sequential effects in task-switching performance. Journal of Experimental Psychology: Learning, Memory, and Cognition, 33(2), 370–378.
Schneider, W., & Shiffrin, R. M. (1977). Controlled and automatic human information processing: I. Detection, search, and attention. Psychological Review, 84(1), 1–57.
Schubert, T. (2008). The central attentional limitation and executive control. Frontiers in Bioscience, 13(13), 3569–3580.
Schubert, T., & Strobach, T. (2018). Practice-related optimization of dual-task performance: Efficient task instantiation during overlapping task processing. Journal of Experimental Psychology: Human Perception and Performance, 44(12), 1884–1904.
Sigman, M., & Dehaene, S. (2006). Dynamics of the central bottleneck: Dual-task and task uncertainty. PLoS Biology, 4, e220.
Stelzel, C., Kraft, A., Brandt, S. A., & Schubert, T. (2008). Dissociable neural effects of task order control and task set maintenance during dual-task processing. Journal of Cognitive Neuroscience, 20(4), 613–628.
Strobach, T., Hendrich, E., Kübler, S., Müller, H., & Schubert, T. (2018). Processing order in dual-task situations: The “first-come, first-served” principle and the impact of task order instructions. Attention, Perception, & Psychophysics, 80(7), 1785–1803.
Strobach, T., Salminen, T., Karbach, J., & Schubert, T. (2014). Practice-related optimization and transfer of executive functions: A general review and a specific realization of their mechanisms in dual tasks. Psychological Research, 78(6), 836–851.
Strobach, T., Schütz, A., & Schubert, T. (2015). On the importance of Task 1 and error performance measures in PRP dual-task studies. Frontiers in psychology, 6, 403.
Sudevan, P., & Taylor, D. A. (1987). The cuing and priming of cognitive operations. Journal of Experimental Psychology: Human Perception and Performance, 13(1), 89–103.
Szameitat, A. J., Lepsien, J., von Cramon, D. Y., Sterr, A., & Schubert, T. (2006). Task-order coordination in dual-task performance and the lateral prefrontal cortex: An event-related fMRI study. Psychological Research, 70(6), 541–552.
Szameitat, A. J., Schubert, T., Müller, K., & von Cramon, D. Y. (2002). Localization of executive functions in dual-task performance with fMRI. Journal of Cognitive Neuroscience, 14(8), 1184–1199.
Töllner, T., Strobach, T., Schubert, T., & Müller, H. J. (2012). The effect of task order predictability in audio-visual dual task performance: Just a central capacity limitation? Frontiers in Integrative Neuroscience, 6, 75.
Vandierendonck, A., Liefooghe, B., & Verbruggen, F. (2010). Task switching: interplay of reconfiguration and interference control. Psychological Bulletin, 136(4), 601–626.
Welford, A. T. (1952). The ‘psychological refractory period’and the timing of high-speed performance—A review and a theory. British Journal of Psychology. General Section, 43(1), 2–19.
Welford, A. T. (1980). The single-channel hypothesis. In A. T. Welford (Ed.), Reaction times (pp. 215–252). London: Academic Press.
Acknowledgements
We would like to thank Lea Skaliks, Jule Ballhausen, Svenja Dymke, Marie Bahnsen, Nell Scheffler, Stina Klein, Pauline Grathwohl, Jorma Slyter, and Özlem Kurtoglu for their assistance in data collection. Data files are available at https://osf.io/wdmz7/?view_only=2a5173b0a7674db892de19451a1d616d.
Funding
This study was funded by a grant of the German Research Foundation (Schu 1397/7-1, 7-2) to T.S. (first author) and to T.S. (last author), and it is part of the Priority Program, SPP 1772.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
T.S. (first author) declares that he has no conflict of interest. S.K. declares that he has no conflict of interest. T.S. (last author) declares that he has no conflict of interest.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Strobach, T., Kübler, S. & Schubert, T. Endogenous control of task-order preparation in variable dual tasks. Psychological Research 85, 345–363 (2021). https://doi.org/10.1007/s00426-019-01259-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-019-01259-2