
Abstract
Different contexts with high versus low conflict frequencies require a specific attentional control involvement, i.e., strong attentional control for high conflict contexts and less attentional control for low conflict contexts. While it is assumed that the corresponding control set can be activated upon stimulus presentation at the respective context (e.g., upper versus lower location), the actual features that trigger control set activation are to date not described. Here, we ask whether the perceptual priming of the location context by an abrupt onset of irrelevant stimuli is sufficient in activating the context-specific attentional control set. For example, the mere onset of a stimulus might disambiguate the relevant location context and thus, serve as a low-level perceptual trigger mechanism that activates the context-specific attentional control set. In Experiment 1 and 2, the onsets of task-relevant and task-irrelevant (distracter) stimuli were manipulated at each context location to compete for triggering the activation of the appropriate control set. In Experiment 3, a prior training session enabled distracter stimuli to establish contextual control associations of their own before entering the test session. Results consistently showed that the mere onset of a task-irrelevant stimulus (with or without a context-control association) is not sufficient to activate the context-associated attentional control set by disambiguating the relevant context location. Instead, we argue that the identification of the relevant stimulus at the respective context is a precondition to trigger the activation of the context-associated attentional control set.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
The activation of context-specific attentional control sets
Adaptive human behavior requires the flexible recruitment and implementation of attentional control mechanisms to shield ongoing task performance from distracting and competing stimuli and action tendencies (Miller & Cohen, 2001). Recent developments in cognitive psychology and neuroscience have shown that attentional control is not only adjusted by means of top–down regulation, but also can be triggered by environmental cues, i.e., stimulus-driven or bottom–up priming of control (Bugg, 2014a; Bugg & Crump, 2012; Egner, 2014; Fischer & Dreisbach, 2015; King, Korb, & Egner, 2012; Mayr & Bryck, 2007).
For example, Crump, Gong and Milliken (2006) presented Stroop trials randomly at one of the two locations. In the Stroop task, participants respond to the ink colors of color words. Importantly, responses are slower and more error prone on conflicting trials (e.g., word blue in red ink) compared to non-conflicting trials (e.g., word red in red ink). This Stroop effect depends also on the frequency of presented trials. For example, Crump et al. (2006) presented a high proportion of conflicting Stroop trials (e.g., 75 % incongruent trials) at one location (e.g., at the upper part of the screen) and a low proportion of conflicting Stroop trials (e.g., 25 % incongruent trials) at the other location (e.g., lower part of the screen). The logic is straightforward: At the context with high conflict frequency, stimulus processing mostly requires an attentional control set that reduces the impact of task-irrelevant stimulus features (e.g., by focusing processing on task-relevant features). At the context with low conflict frequency (mostly congruent trials), stimulus processing is more affected by irrelevant stimulus features because of a more relaxed attentional control set. As a consequence, Stroop effects were reduced at the context of high compared to low conflict frequencies. This effect is known as the context-specific proportion congruence (CSPC) effect (Crump et al., 2006; Crump & Milliken, 2009; Crump, Vaquero, & Milliken, 2008). The CSPC effect has been replicated in various conflict paradigms, such as in Flanker tasks (Corballis & Gratton, 2003; King et al., 2012; Vietze & Wendt, 2009; Wendt, Kluwe, & Vietze, 2008), priming tasks (Heinemann, Kunde, & Kiesel, 2009; Reuss, Desender, Kiesel, & Kunde, 2014), and dual tasks (Fischer, Gottschalk, & Dreisbach, 2014) and has been extended to different types of contextual cues that are effective to produce CSPC effects, such as font (Bugg, Jacoby, & Toth, 2008), color (Lehle & Hübner, 2008; Vietze & Wendt, 2009), shape (Crump et al., 2006) and social categories (Cañadas, Rodríguez-Bailón, Milliken, & Lupiáñez, 2013).
As an explanation for CSPC effects, it has been proposed that the repeated application of an attentional control set for processing stimuli at a certain context establishes an associative link between the context (e.g., location) and the corresponding attentional control set in episodic memory (Bugg & Crump, 2012; Crump & Milliken, 2009). Processing the context cue is assumed to activate the associated attentional control set which then biases processing selectivity. This bottom–up priming of attentional control sets is astonishing because the context feature per se, i.e., location of stimulus presentation, is completely irrelevant for handling the instructed task.
Although the assumption of the experience-based formation of a context-control link appears plausible, to date, however, it remains largely unclear how this associative control link is flexibly activated by the context cue. Therefore, in the present study, we applied a location-specific proportion congruence paradigm (Crump et al., 2006) and examined the underlying mechanisms by which an attentional control set is activated by its associated location context. Describing the cognitive mechanisms underlying the context-specific bottom–up recruitment of attentional control sets will further the understanding of how the mind realizes rapid online adjustments of cognitive control to enable behavioral adaptation.
For this we assume that the repeated experience of stimulus processing under the application of specific attentional control sets at a given location establishes an associative compound that includes various features, potentially serving as trigger condition for attentional control set activation (Bugg & Crump, 2012). More specifically, not only specific items that appear at the location, but also responses that are executed, the experience of the conflict status, or contextual features that are employed during performance, are linked together in episodic memory (Bugg & Crump, 2012; Crump & Milliken, 2009) in the form of an event file (Hommel, Müsseler, Aschersleben, & Prinz, 2001). Theoretically, the associative context-control link could be activated by (a) the mere detection of a certain stimulus exemplar, (b) the detection of conflict or (c) the context itself, when for example the appearance of the stimulus at a certain location disambiguates the context.
Although the activation of attentional control sets by specific item exemplars is widely acknowledged and has become known as item-specific proportion congruence (ISPC) effects (Blais, Robidoux, Risko, & Besner, 2007; Bugg & Hutchison, 2012; Bugg, Jacoby, & Chanani, 2011; Jacoby, Lindsay, & Hessels, 2003), for the present location-specific activation of attentional control sets a sole explanation on the basis of individual item exemplars appears unlikely. In particular, for CSPC effects all items can appear with the same frequency at each context (e.g., Fischer et al., 2014). Furthermore, CSPC effects have also been demonstrated for individual unbiased stimuli, i.e., stimuli that appear with equal conflict probability (50/50) at each location (Crump & Milliken, 2009).
It seems also unlikely that conflict processing per se is the core trigger mechanism to recruit the associated attentional control set. Neither conflict awareness nor full information about the conflict frequency manipulation, influence or enhance the CSPC effect (Blais, Harris, Guerrero, & Bunge, 2012; Crump et al., 2008; Ghinescu, Schachtman, Stadler, Fabiani, & Gratton, 2010). For example, in a recent study, we showed that pre-cuing the conflict likelihood of the upcoming trial with 100 % validity did not affect the contextual adjustment of processing selectivity (Fischer et al., 2014). Although participants were explicitly informed about the cuing procedure and were instructed to use the pre-cues, they were not able to utilize the pre-knowledge about the upcoming conflict status to activate the respective attentional control set (for similar difficulties in administering top–down control over interference by pre-cues predicting conflict, see for example Wühr & Kunde, 2008).
In contrast, when pre-cues validly predicted the location of the next trial, the occurrence of CSPC effects was facilitated. Thus, knowledge about the relevant location facilitates the activation of the associated attentional control set (Fischer et al., 2014). This result points towards a specific role of location as context cue to trigger context-sensitive adjustments of processing selectivity. The unique role of location may be founded on the fact that spatial information is a very salient feature which is processed automatically (Logan, 1998) and receives priority during encoding (Mayr, 1996; for a thoughtful comment see Bugg, 2014b). In fact, CSPC effects are also easier to be found when location compared to other features (e.g., shape) serves as context cue (Crump et al., 2008). In addition, Wendt, Kluwe and Vietze (2008) demonstrated CSPC effects for up to four different locations, which underpins that location might be a predestinated feature to serve as contextual cue.
The saliency of the location feature may be at least partially built on the abrupt onset of stimuli that appear at the specific location. For example, in Fischer et al. (2014) the onset of stimuli at one location was additionally accompanied by the offset of the fixation field at the opposite location, thus creating a strong perceptual event. The abrupt onset of a stimulus triggers an involuntary allocation of visual attention (Yantis & Jonides, 1984, 1990). It is assumed that the abrupt appearance of a new object requires the formation of an object representation which automatically shifts attention to the new object (Yantis & Hillstrom, 1994). This attentional capture binds processing resources which obstructs the processing of other stimuli. As a consequence, participants cannot prevent the allocation of visual attention towards the abrupt onset of task-irrelevant stimuli, which delays identification and responding to task-relevant stimuli (Kim & Cave, 1999; Schreij, Owens, & Theeuwes, 2008; Theeuwes, 1995, 2010; Theeuwes, Atchley, & Kramer, 2000; Yantis & Jonides, 1990). Importantly, involuntary attentional capture by task-irrelevant stimuli occurs when these stimuli appear at the same time with relevant stimuli or a delay of up to 100 ms (Kim & Cave, 1999; Theeuwes et al., 2000). For longer durations of abrupt task-irrelevant stimulus onset (i.e., >150 ms), top–down control can be initiated to override the capturing effect by the abrupt onset (for a review see Theeuwes, 2010; Theeuwes et al., 2000).
In typical location-based CSPC studies stimuli appear with an abrupt onset at either location. Therefore, the attentional capture initiated by the abrupt stimulus onset and its automatic capture of processing resources may reflect a low-level attentional mechanism that facilitates the activation of the attentional control set that is associated with the location.
In the present study, we tested whether an abrupt stimulus onset at a given location is sufficient to activate the location-associated attentional control set or whether the identification of abrupt appearing stimuli (e.g., determination of task relevance) at a location is a necessary precondition for the activation of the associated attentional control set. In three experiments, we presented task-relevant stimuli (numbers) at one location and additional task-irrelevant distracter stimuli (letters) at the opposite location. In Experiment 1, task-relevant stimuli were presented either in the absence of task-irrelevant stimuli at one of two locations forming a strong perceptual event for this context alone, or were presented with simultaneously occurring task-irrelevant stimuli that were presented at the opposite location, to eliminate the abrupt attentional capture by a single location context. In Experiment 2, either task-irrelevant or task-relevant stimuli received a temporal head start to implement a strong attentional bias towards the location of first stimulus presentation. In Experiment 3, task-irrelevant letter stimuli were included in a prior practice block to build location-specific context-control association for these letter stimuli. In the test block, both task-relevant number stimuli and task-irrelevant letter stimuli were presented at either location. To foreshadow the results, in none of the experiments, the abrupt onset alone was sufficient to trigger the activation of the associated attentional control set, but required the identification of the stimulus’ task relevance.
Experiment 1
Experiment 1 was closely modeled after the study of Fischer et al. (2014) in which we applied a dual-task paradigm with large between-task conflicts (i.e., response-category compatibility effects) that are sensitive to CSPC manipulations. Participants performed a prioritized number categorization (i.e., odd–even judgment) on a digit (S1) in Task 1 and another number categorization (i.e., odd–even judgment) on a digit (S2) in Task 2. Both digit stimuli (S1 and S2) were presented together at one of two locations. The same task set (i.e., odd–even judgment) was implemented for categorizing S1 and S2 to obtain strong response-category (RC) compatibility effects (see Fischer, Miller, & Schubert, 2007; Logan & Schulkind, 2000). That is, responses in Task 1 are fast for RC compatible trials, when the same categorization is required for both digits (e.g., S1 and S2 are both odd). Responses in Task 1 are slowed for RC incompatible trials, when S2 requires the opposite categorization (e.g., odd) than S1 (e.g., even). Subtracting performance measures in RC compatible from RC incompatible trials denotes the RC compatibility effect, the extent of which reflects the impact of concurrent T2 processing on T1 performance (see Fischer et al., 2007; Lien & Proctor, 2002; Logan & Schulkind, 2000).Footnote 1
Now, the proportion of RC compatible and RC incompatible trials was manipulated for two different locations (i.e., above and below fixation). At one location, RC incompatible trials were presented with a high proportion (e.g., 80 % at location above). At the opposite location RC incompatible trials were presented with a low proportion (i.e., 20 % at location below). This proportion manipulation creates a context with high conflict frequency (HCF, 80 % RC incompatible) and a context with low conflict frequency (LCF, 20 % RC incompatible), respectively. Fischer et al. (2014) showed that participants acquire these contingencies and demonstrate reduced RC compatibility effects for HCF compared to LCF contexts.
Critically, in Experiment 1 in addition to task-relevant stimuli appearing at one location (distracter absent condition), in 50 % of the trials we presented distracter stimuli at the opposite location (distracter present condition). In the distracter absent condition, the abrupt onset of task-relevant stimuli disambiguates the location by involuntarily capturing visual attention to the location of stimulus presentation. In the distracter present condition we reasoned that both stimuli at either location require the formation of a new object representation. Both stimuli capture visual attention and compete for selection. This is typically shown in slowed responses to target stimuli in the distracter present condition and proves an attentional capture by task-irrelevant stimuli.
Importantly, if task-relevant and irrelevant stimuli capture visual attention, the onset of task-relevant stimuli cannot easily disambiguate the location by means of mere stimulus onset. If the perceptual event of an abrupt stimulus onset triggers the activation of the associated control set, the CSPC should be diminished in the distracter present condition.
On the other hand, demonstrating a CSPC effect in both distracter conditions renders the attention capture as the main mechanism activating attentional control sets unlikely. That is, assuming that the onset of stimuli at both locations captures attention to equal shares, the attentional event of attention capture cannot explain the occurrence of location-based CSPC effects.
Method
Participants
Thirty-four students (31 female, 18–35 years, mean age 20.85 years ± 3.67) of the Technische Universität Dresden participated in the experiment, which took about 60 min. Participants received course credits or 5 €.
Stimuli and apparatus
The digits 1–9 (except 5) served as task-relevant stimuli and the letters A, B, E, K, M, O, P and U as task-irrelevant distracter stimuli. All stimuli were presented in white font Arial on black background on a 19″ TFT-monitor (1280 × 1024 pixel). Font size was 20 for digits and 18 for letters.Footnote 2 Four horizontal dashes (each 2 × 0.8 mm) in a squared arrangement formed a fixation field (10 × 11 mm) and served as placeholders for the presentation of two task-relevant (S1 and S2) or two task-irrelevant stimuli (D1 and D2). The fixation field was presented in the upper half (+8.5 mm above screen center) as well as in the lower half (−8.5 mm below screen center) of the screen, thus defining two separate locations (location above versus location below). With a viewing distance of 60 cm both fixation fields extended a visual angle of 4.15°. Irrespective of location on the screen, S1 included the digits 2, 3, 7 and 8, and were presented between the upper two dashes. S2 consisted of the digits 1, 4, 6 and 9, and were presented between the lower two dashes. At the location opposite to S1 and S2, in half of the trials, letter stimuli A, K, M and U, served as D1 and were presented between the upper two dashes whereas the letter stimuli B, E, O and P, served as D2 and were presented between the lower two dashes. In each trial S1 and S2 were presented together at either location (above versus below), whereas in half of the trials D1 and D2 occupy the opposite location. Responses to S1 were made with the index finger (“,” key) and middle finger (“.” key) of the right hand. Responses to S2 were made with the index finger (“X” key) and middle finger (“Y” key) of the left hand on a QWERTZ keyboard. Letter stimuli were presented as task-irrelevant distracter stimuli and afforded no response. A Pentium I computer with a Windows XP platform (Service Pack 2) equipped with Presentation software (Version 16.3 Neurobehavioral Systems) was used for stimulus presentation and data recording.
Procedure
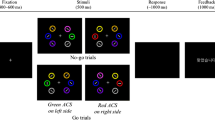
Each trial began with the simultaneous presentation (1100 ms) of a fixation cross at the screen center and the two fixation fields, above and below the screen center, respectively. For further 400 ms the fixation cross vanished and only the two fixation fields remained. In the distracter present condition, S1 and D1 appeared simultaneously between the upper two dashes in either location. After 40 ms S2 and D2 were presented jointly between the lower two dashes in the respective location. In the distracter absent condition, S1 appeared between the upper two dashes at one location, whereby the fixation field at the opposite location disappeared (see Fig. 1). After 40 ms, S2 was presented between the lower two dashes at the same location. After another 1000 ms all stimuli and fixation fields were substituted by a black screen for 2500 ms. Feedback was provided for about 500 ms (“correct”, “false” or “too slow”). The next trial started after a random ten-step interval between 100 and 1000 ms.

Experimental procedure in Experiment 1. S1 and S2 represent digit stimuli for T1 and T2 of the dual task. D1 and D2 represent distracter stimuli to match the timing of the abrupt stimulus onset at the opposite location in half of the trials, whereas in half of the other trials there were no distracter stimuli present
Participants were instructed to categorize S1 as either odd (right index finger) or even (right middle finger) and S2 also as either odd (left index finger) or even (left middle finger). Instructions further emphasized Task 1 priority. That is, they were informed to first categorize S1 as fast and as accurately as possible and only then categorize S2 as fast and accurately as possible. Participants were further informed about the presence of task-irrelevant letter stimuli and were asked to refrain from grouping responses in T1 and T2.
To familiarize participants with the experimental paradigm, the experiment started with a practice block (48 RC compatible and 48 RC incompatible trials) in which the fixation display was presented at the screen center and only task-relevant digit stimuli (S1 and S2) were used. Subsequently, participants worked through two blocks (160 trials each) in which both task-relevant and task-irrelevant stimuli were placed at either location. Importantly, experimental blocks contained the location-specific conflict frequency manipulation. That is, for half of the participants, one location (e.g., above) contained 80 % RC compatible and 20 % RC incompatible trials, thus representing a context of low conflict frequency (LCF). This proportion was reversed at the opposite location (e.g., below) including 20 % RC compatible and 80 % RC incompatible trials, thus representing a context of high conflict frequency (HCF). For the other half of the participants the pairing of conflict frequency with location was reversed. In consequence, within this experiment four stimuli in either task (T1 and T2, respectively), were used, which amounted to sixteen different S1–S2 compounds (i.e., eight RC compatible and eight RC incompatible trials). These sixteen S1–S2 compounds were repeatedly presented with different frequencies across two locations. This amounts to thirty-two single trials in order to present each S1–S2-location compound once. Altogether, the high conflict frequency location contains sixteen repetitions of RC incompatible and four repetitions of RC compatible trials, whereas the low conflict frequency context contains sixteen repetitions of RC compatible trials and four repetitions of RC incompatible trials. Overall, conflict frequency was based on 50 % RC compatible and 50 % RC incompatible trials. Location, stimuli, conflict status, and responses were not predictable in a given trial. A short break was provided after every 40 trials.
Design
A 2 (context: HCF; LCF) × 2 (RC compatibility: RC compatible; RC incompatible) × 2 (distracter condition: present; absent) × 2 (block: 1st block; 2nd block) ANOVA with repeated measures on RT1 and error percent was applied.
Results
Error rates consisted of erroneous responses in either task, because this is considered as unsuccessful dual-task performance (Logan & Schulkind, 2000). Errors (4.9 %) and trials in which RT1 or RT2 exceeded >3 standard deviations from the individual condition mean (1.3 %) were excluded prior to RT analyses. RT1, RT2 and error rates of all experiments are presented in Table 1. In the present study, however, T1 performance (RT1) was of primary interest (see also Fischer et al., 2014).
RT1
Participants reacted faster in RC compatible (875 ms) compared to RC incompatible trials (1089 ms), F(1, 33) = 131.36, p < .001, \(\eta_{p}^{2}\) = 0.80. The factor block influenced RT1, expressed in higher RT1s for Block 1 (1026 ms) compared to Block 2 (938 ms), F(1, 33) = 114.29, p < .001, \(\eta_{p}^{2}\) = 0.78. Notably, the factor distracter condition affected RT1, F(1, 33) = 191.47, p < .001, \(\eta_{p}^{2}\) = 0.85. RT1 were larger for the distracter present (1028 ms) than for distracter absent condition (936 ms). Most importantly, there was a context-specific modulation of the RC compatibility effect, with significantly smaller RC compatibility effects at the HCF context (184 ms) compared to the LCF context (243 ms), F(1, 33) = 13.71, p = .001, \(\eta_{p}^{2}\) = 0.29 (see Table 1). This CSPC effect was not further modulated by the presence or absence of distracter stimuli, F < 1. No further effects were significant, all ps > .11. Separate Anovas for each distracter condition with the factors Context × RC compatibility × Block reveal a reliable context-specific modulation of the RC compatibility effect for both the distracter present condition (66 ms), F(1, 33) = 8.07, p = .008, \(\eta_{p}^{2}\) = 0.20, and the distracter absent condition (52 ms), F(1, 33) = 8.68, p = .006, \(\eta_{p}^{2}\) = 0.21.
Errors
Error rates were affected by RC compatibility, F(1, 33) = 25.85, p < .001, \(\eta_{p}^{2}\) = 0.44, with lower error rates on RC compatible trials (3.6 %) compared to RC incompatible trials (6.8 %). Participants committed fewer errors in Block 2 (4.2 %) compared to Block 1 (6.3 %), F(1, 33) = 19.23, p < .001, \(\eta_{p}^{2}\) = 0.37. The factor context also affected error rates, F(1, 33) = 10.93, p = .002, \(\eta_{p}^{2}\) = 0.25, with fewer errors in the HCF context (4.4 %) than in the LCF context (6.1 %). The factors block, RC compatibility and context were close to interact, F(1, 33) = 3.86, p = .058, \(\eta_{p}^{2}\) = 0.11. In particular, the CSPC effect was more pronounced at the beginning of the experiment (Block 1, 4.0 %) than in the end of the experiment (Block 2, −0.1 %). Analysis revealed a trend for the contextual modulation of the RC compatibility effect, F(1, 33) = 3.00, p = .093, \(\eta_{p}^{2}\) = 0.08, as indicated in a reduced RC compatibility effect in the HCF context (2.3 %) compared to the LCF context (4.2 %). The CSPC effect was not affected by distracter condition F < 1. No further effects were significant, all ps > .124.
Discussion
Results demonstrate a reliable CSPC effect with smaller conflict effects at the HCF context compared to the LCF context. This contextual modulation of the RC compatibility effect was obtained irrespective of distracter condition. Importantly, the presence of task-irrelevant stimuli significantly slowed responses, indicating substantial attentional capture. In this condition, neither context location received a perceptual saliency advantage on the basis of abrupt stimulus onset. Both locations were equally likely of receiving an initial allocation of visual attention. A reliable context-specific adjustment of processing selectivity in these conditions, however, speaks against the assumption that the mere stimulus onset activates the associated attentional control set. Instead we propose that the discrimination between task-relevant and task-irrelevant is needed for the recruitment of the attentional control set that is applied for stimulus processing.
Experiment 2
In Experiment 2, we aimed at replicating findings of Experiment 1 by implementing conditions of increased perceptual saliency for task-irrelevant stimuli. We provided either task-relevant or task-irrelevant stimuli with a temporal head start at the respective location. The head start amounted to 80 ms, which is (a) within the range of allowing for involuntary attentional capture by the abrupt onset of the stimulus and (b) is too short to allow for voluntary and strategic adjustment counteracting the capturing effect of the abrupt onset (e.g., Theeuwes et al., 2000). Thus, in half of the trials task-irrelevant stimuli appeared with an abrupt onset at the irrelevant location before task-relevant stimuli were presented at the opposite location (and vice versa). Again, this manipulation was thought to implement an unpredictable attentional bias to the location of first stimulus appearance, irrespective of task relevance.
If an abrupt stimulus onset at a location is sufficient of triggering the location-associated attentional control set, this should lead to an activation of the incorrect attentional control set by the perceptually salient task-irrelevant stimuli. As a result, CSPC effects should be smaller when task-irrelevant stimuli appear prior to task-relevant stimuli. In contrast, if an attentional control set is activated by processing the stimuli to which the control set is applied to, CSPC effects should be obtained despite the perceptual salience advantage of the task-irrelevant stimuli.
Method
Participants
Twenty students (13 female, 20–33 years, mean age 24.5 years ± 3.46) of the Technische Universität Dresden participated at this experiment, which took about 60 min. Participants received course credits or 5 €.
Stimuli, apparatus and procedure
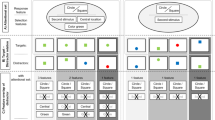
The experimental setup was identical to Experiment 1 except when noted otherwise. In half of the trials, distracter stimuli (D1 and D2) appeared 80 ms earlier than task-relevant digit stimuli (S1 and S2). The onset of D1 between the upper dashes at one location was followed by the onset of D2 between the lower two dashes after 40 ms. After another 40 ms, S1 was presented between the upper dashes at the opposite location to D1. S2 followed S1 by another 40 ms. In the other half of the trials the task-relevant digit stimuli (S1 and S2) instead of the distracter stimuli (D1 and D2) appeared first (see Fig. 2). In contrast to Experiment 1, distracter stimuli were present in each trial.

Experimental procedure in Experiment 2 and 3. S1 and S2 represent digit stimuli for T1 and T2 of the dual task. D1 and D2 represent distracter stimuli, whereas either distracter stimuli or task-relevant digit stimuli received a 80-ms head start of abrupt stimulus onset at a certain location
Design
A 2 (context: HCF; LCF) × 2 (RC compatibility: RC compatible; RC incompatible) × 2 (block: 1st block; 2nd block) × 2 (onset order: task-relevant first, task-irrelevant first) ANOVA with repeated measures on RT1 and error percent was applied.
Results
As in Experiment 1, erroneous trials (4.2 %) and trials that exceeded the outlier criterion (1.1 %) were excluded prior to RT analysis.
RT1
A significant RC compatibility effect was obtained with smaller RT1 in RC compatible (949 ms) compared to RC incompatible trials (1240 ms), F(1, 19) = 47.11, p < .001, \(\eta_{p}^{2}\) = 0.71. Responses grew faster from Block 1 (1143 ms) to Block 2 (1046 ms), F(1, 19) = 16.50, p = .001, \(\eta_{p}^{2}\) = 0.47, and were slightly larger in the HCF context (1105 ms) compared to the LCF context (1084 ms), F(1, 19) = 3.61, p = .073, \(\eta_{p}^{2}\) = 0.16. Most importantly, the context-specific modulation of the RC compatibility effect was reliably found, F(1, 19) = 6.86, p = .017, \(\eta_{p}^{2}\) = 0.27. RC compatibility effects were considerably smaller in the HCF (269 ms) than in the LCF context (313 ms). Furthermore, this CSPC effect was independent of onset order (i.e., relevant versus irrelevant stimulus onset first), F < 1. No further effects were significant, all ps > .167.
Errors
Error rates were affected by RC compatibility, F(1, 19) = 13.97, p = .001, \(\eta_{p}^{2}\) = 0.42, with lower error rates in RC compatible trials (3.0 %) compared to RC incompatible trials (5.4 %). No further effects were significant, all ps > .118.
Discussion
Despite the unpredictability of stimulus onset at either task-relevant or task-irrelevant location a reliable CSPC effect was found. This result further adds to the assumption that the mere low-level perceptual feature of stimulus onset at a location might not be sufficient to trigger the activation of associated attentional control sets. Instead stimulus processing to the point of identifying task relevance might be a precondition for control activation.
Experiment 3
So far we can argue that the mere perceptual event of an abrupt onset of any stimulus is not sufficient to trigger a control set that is linked to the location of stimulus appearance. In both previous experiments stimuli needed to be processed according to their task rule to activate the respective attentional control set. In both experiments, the onset of distracter letter stimuli, however, could have easily been discarded because letter stimuli were never associated with any categorization rule, responses or conflict frequency manipulations. In Experiment 3 we asked whether the onset of (task-irrelevant) stimuli can trigger the activation of an attentional control set, if these stimuli were associated with a categorization rule and the experience of a conflict frequency manipulation. The activation of an attentional control set that is activated by the onset of task-irrelevant stimuli at a certain location should interfere with the activation or implementation of the attentional control set needed to process task-relevant stimuli.
Experiment 3 started with separate letter- and digit categorization practice phases in which a list-wide conflict proportion manipulation was introduced. Importantly, in each list of HCF and LCF, all stimuli were presented at the respective location that served as location context in the subsequent CSPC experiment (e.g., HCF—above, LCF—below, or vice versa). This was thought to establish associative links between context and attentional control settings for both, letter stimuli and digit stimuli, respectively (see also Fischer et al., 2014).
In the letter practice phase, participants were asked to categorize letters (D1 and D2) as either vowels or consonants. Importantly, D1 and D2 also formed RC compatible (i.e., both letters were vowels, or both letters were consonants) and RC incompatible trials (i.e., one letter was a vowel and the other a consonant or vice versa). To reiterate, in the list with HCF all stimuli were presented exclusively at the location that served as HCF context (e.g., above) in the subsequent experiment. In the list with LCF, all stimuli were presented at the opposite location that later served as LCF context (e.g., below). The same practice phase was applied for digit stimuli (Fischer et al., 2014).
In the subsequent experimental blocks, both stimulus types were presented as in Experiment 2. That is, the temporal onset advantage of task-relevant digit stimuli (S1 and S2) and that of task-irrelevant letter stimuli (D1 and D2) at either location alternated unpredictably from trial to trial (see Experiment 2).
Although letter stimuli were again task-irrelevant distracter stimuli, it is conceivable that they still trigger the activation of the associated and formerly relevant attentional control set which then interferes with the attentional control set required for processing task-relevant stimuli. Such an assumption is not trivial, as recent research demonstrated that the repeated presentation of material that is related to previously active but completed intentions is capable of reactivating intention-associated memory contents that interferes with ongoing task performance (e.g., Scullin, Bugg, & McDaniel, 2012; Walser, Fischer, & Goschke, 2012; Walser, Goschke, & Fischer, 2014).
Method
Participants
Thirty-two students (24 female, 19–33 years, mean age 24.2 years ± 3.72) of the Technische Universität Dresden participated at this experiment, which took about 70 min. Participants received course credits or 6 €.
Stimuli, apparatus and procedure
Stimuli, apparatus and procedure were taken from Experiment 2. However, Experiment 3 started with two new practice phases, one for digits and one for letters, respectively. Each practice phase consisted of 16 trials for task familiarization (with centrally presented fixation field) and a list-wide conflict frequency manipulation. In the practice phase for letters, participants were required to categorize letters presented between the upper two dashes of the fixation field (D1) and letters between the lower two dashes (D2) as vowels or consonants. Responses to D1 and D2 were made with the index (vowel) and middle (consonant) finger of the right and left hand, respectively. After the 16 familiarization trials, letter stimuli were presented exclusively at one location (e.g., above) in a mini block of 40 trials. In this block a list-wide conflict frequency manipulation was implemented (e.g., high conflict frequency). In a subsequent mini block of 40 trials, all letter stimuli were presented exclusively at the opposite location (e.g., below) and the mini block contained the opposite list-wide conflict frequency manipulation (e.g., low conflict frequency). This practice phase for letter stimuli was followed by the analogous practice phase for digit stimuli.
Order of practice phases (letter versus digits), first location of mini block trials (above versus below) and location conflict frequency assignment (high conflict frequency above versus below) were counter-balanced across participants. After both practice phases, participants were informed that the letter task is finished and that for the subsequent experiment letter stimuli could be ignored.
Design
A 2 (context: HCF; LCF) × 2 (RC compatibility: RC compatible; RC incompatible) × 2 (block: 1st block; 2nd block) × 2 (onset order: task-relevant first, task-irrelevant first) ANOVA with repeated measures on RT1 and error percent was applied.
Results
RT and error analyses were based on performance in the experimental blocks. For RT analysis errors (5.8 %) and trials that exceeded the outlier criterion (1.1 %) were excluded.
RT1
Participants reacted faster in RC compatible (944 ms) compared to RC incompatible trials (1149 ms), F(1, 31) = 105.50, p < .001, \(\eta_{p}^{2}\) = 0.77. Furthermore, the factor block affected RT1, expressed in higher RTs for Block 1 (1074 ms) compared to Block 2 (1019 ms), F(1, 31) = 29.09, p < .001, \(\eta_{p}^{2}\) = 0.48. Most importantly, there was a context-specific modulation of the RC compatibility effect, F(1, 31) = 7.91, p = .008, \(\eta_{p}^{2}\) = 0.20, denoting a smaller RC compatibility effect at the location with high (181 ms) compared to the location with low conflict frequency (229 ms). Furthermore, this CSPC effect was independent of whether relevant or irrelevant stimuli were presented first, F < 1. No other effects were significant, all p’s > .126.
Errors
Error rates were affected by RC compatibility, F(1, 31) = 6.36, p = .017, \(\eta_{p}^{2}\) = 0.17, with lower error rates on RC compatible trials (4.9 %) compared to RC incompatible trials (6.9 %). No other effects were significant, p’s > .138. All four factors, however, were close to interact, F(1, 31) = 3.90, p = .057, \(\eta_{p}^{2}\) = 0.11. CSPC effects in error rates did not differ between head start conditions in the first block (F < 1), but seemed to grow apart for the second block (4.5 and −2.8 % for onset head start of task-irrelevant and -relevant, respectively), F(1, 31) = 6.41, p = .017, \(\eta_{p}^{2}\) = 0.17. At present, we do not have an explanation for this trend.
Additional analysis
To check whether letter stimuli formed a proportion congruency effect in a list-wide manner during the learning phase, we analyzed RT1 in a 2 (context: HCF-list; LCF-list) × 2 (RC compatibility: RC compatible; RC incompatible) ANOVA with repeated measures. This analysis revealed a highly significant interaction of these two factors, F(1, 31) = 47.86, p < .001, \(\eta_{p}^{2}\) = 0.61. The RC compatibility effect was strongly reduced for the high conflict frequency list (51 ms) compared to the low conflict frequency list (240 ms). Importantly, during the learning phase each list was presented exclusively at one specific location. Thus, it is plausible to assume that the repeated application of an attentional control set at each location formed an association between the location at which stimuli were presented and an attentional control set.
Discussion
Results of the experimental blocks showed a robust CSPC effect with a reduced RC compatibility effect at the high conflict frequency context compared to the low conflict frequency context. This CSPC effect was obtained irrespective of whether the onset of task-irrelevant distracter stimuli preceded the onset of task-relevant stimuli at the opposite location, thus, replicating findings from Experiment 2. The mere disambiguation of a location by the onset of irrelevant stimuli is, therefore, not sufficient to trigger an associated attentional control set. Again, stimuli occurring at a location need to be identified to the degree of determining task relevance to activate the respective attentional control set, associated with this location. In Experiment 3, distracter stimuli were additionally introduced with a categorization rule and received training with list-wide location-specific control requirements in a prior learning phase. Even on trials with prior onset, this did not lead to a reduction of the CSPC effect (e.g., by means of concurrently activating the formerly relevant attentional control set).
It should be noted that the condition of task-irrelevant stimuli appearing first likely includes additional subsequent processes to attentional capture by abrupt onsets than the condition in which task-relevant stimuli appeared first. For the latter, elaborate processing including attentional control set activation can start immediately upon stimulus onset and identification. For the condition of task-irrelevant stimuli being presented first, however, an additional re-orienting to the opposite location is required after the identification of stimuli as task irrelevant. At this point, however, it remains speculative whether, and if so to which extent, these processes might influence the activation of the attentional control set after the onset and identification of stimuli.Footnote 3
General discussion
Previous research has shown that a context (e.g., location) can become associated with a specific attentional control set when this attentional control set is frequently applied for stimulus processing at this context. In the present study, we investigated how an associated attentional control set is activated by the context. Specifically, we tested the hypothesis that low-level visual features, such as an abrupt visual stimulus onset at a certain location is sufficient for activating the established context-control link. We reasoned that an abrupt visual onset at one of two locations might serve as location-disambiguation mechanism, which facilitates attentional control set activation.
Three experiments used a location-based CSPC manipulation in which the abrupt visual onset of task-relevant stimuli at one location was accompanied by the abrupt visual onset of task-irrelevant stimuli at the opposite location. In Experiment 1, we tested whether the presence of irrelevant stimuli affects the CSPC. In half of the trials task-relevant stimuli appeared abruptly at either location whereas in the other half of trials task-irrelevant stimuli appeared simultaneously with task-relevant stimuli but at the opposite location. The presence of irrelevant stimuli substantially slowed responses to task-relevant stimuli. According to Yantis (1998), the simultaneous abrupt onset of relevant as well as irrelevant stimuli necessitates the formation of new object representations for both stimuli. Therefore, involuntary attentional capture applies for relevant and irrelevant stimuli, which results in competition for selection. Even the assumption of a random capture by either stimulus (e.g., Reder, Weber, Shang, & Vanyukov, 2003) would reveal a strong impact on the selection process and slow down responses (cf. Fischer & Hagendorf, 2006). Therefore, we conclude that the presence of task-irrelevant stimuli lead to an involuntary capture of visual attention. Importantly, the CSPC effect in Experiment 1 was not at all affected by the presence or absence of irrelevant stimuli.
While the results of Experiment 1 showed that distracter items presented simultaneously with the task-relevant stimuli involuntary capture visual attention, in Experiments 2 and 3 in half of the trials task-irrelevant stimuli occurred with a head start at the opposite location to the abrupt onset of task-relevant stimuli. The head start of task-irrelevant stimuli was delivered 80 ms prior to the onset of task-relevant stimuli at the other location. This time window is suitable for an abrupt onset to capture attention (Schreij et al., 2008; Theeuwes, 1994, 1995, 2010; Theeuwes et al., 2000; Yantis, 1993) as attentional capture by abrupt onsets is most evident during the first 100 ms (for a review see Theeuwes, 2010). In fact, studying top–down and bottom–up control of visual attention, in a visual search study Theeuwes et al. (2000) showed that the distracter onset had the strongest effect of capturing attention (e.g., disturbing target detection) when it was presented in very close temporal proximity to the target (e.g., less than 100 ms) but not when the temporal proximity between target and distracter exceeded 150 ms. For the latter condition, participants were able to initiate sufficient top–down control to diminish the effect of attention capture by the distracter (for further discussion see Theeuwes, 2010).
In both experiments we found context-specific control adjustments irrespective of different intensities of attention capture modulation and despite the establishment of a strong connection between distracter stimuli and context-control associations (Experiment 3). Therefore, while the temporal delay between task-relevant and task-irrelevant stimuli in Experiments 2 and 3 was suited to trigger involuntary capture of visual attention, the present results converge on the assumption that a perceptual bias by abrupt appearing stimuli is not sufficient to trigger the retrieval of an associated attentional control set.
In the present experiments, task-relevant stimuli (S1 and S2) were accompanied by task-irrelevant distracter stimuli (D1 and D2) at the opposite location. In Fischer et al. (2014) we used the identical experimental setup and trial numbers. In the most comparable Experiment 2, with only task-relevant stimuli abruptly appearing at either location, the CSPC effect did not exceed 50 ms and was thus, virtually identical in size to the ones obtained in the present study (see Table 1). Thus, even in conditions in which participants can perfectly rely on the abrupt onset-induced attentional capture revealing the required location (without any distracter expectation), CSPC effects are not more pronounced than in conditions in which task-irrelevant stimuli might capture attention first.
On the contrary, one could even argue that CSPC effects are easier to establish when both locations contain stimuli than when stimuli are exclusively presented at one location. In the Fischer et al. study (e.g., Experiment 2), CSPC effects developed over time and were only observed in the second half of the experiment. In the present study, CSPC effects were present across the entire experiment. Thus, it is conceivable that the location as context feature is more salient, when both locations are occupied by stimuli. This could enable a referential coding. Stimuli at the upper location, for example, might be more strongly perceived as “upper” when coded in reference to stimuli at the lower location compared to when the lower location is not filled with items. In line with this reasoning, CSPC effects were significantly observed in the first block at least for Experiment 1 (51 ms), F(1, 33) = 4.85, p = .035, \(\eta_{p}^{2}\) = 0.13 and for Experiment 3 (37 ms), F(1, 31) = 5.47, p = .026, \(\eta_{p}^{2}\) = 0.15.Footnote 4 It might thus be easier to activate the context-control link when the location containing relevant stimuli is more salient.
Taken together, our results suggest that the perceptual event of an abrupt stimulus onset is unlikely to serve as single trigger condition for the activation of attentional control sets but that the identification of task relevance is a necessary precondition for the application of context-specific control adjustments. Therefore, while proactive control by means of endogenous cuing of the upcoming location can facilitate CSPC effects (Fischer et al., 2014, Experiment 3), the mere presentation of any stimulus at a certain location does not seem sufficient to activate the location-associated attentional control set.
A central discussion in the study of CSPC effects concerns the question to which extent these effects reflect the context-specific recruitment of cognitive control or may be produced by alternative mechanisms, such as low-level contingency learning (Schmidt, 2013; Schmidt & Besner, 2008). Manipulating the proportion congruence often means that some specific stimulus compounds are presented more often than others, which allows for the prediction of the contingent response. To illustrate, in a proportion congruence Stroop task, the word BLUE, for example, is more often presented in the color blue in high proportion congruence (LCF) blocks. Here, the word BLUE is predictive of the response to the color blue. In a low proportion congruence block (HCF block), the word BLUE might be more often presented in the color green and is thus is prognostic of the response to the color green. Therefore, the contingency between word–color combinations predicts a specific response. In a location-based CSPC paradigm, the word BLUE occurs equally often as congruent and as incongruent compound. However, the combination with a specific location might be predictive for the corresponding response (e.g., at the LCF location, the word BLUE is predictive for the response to the color blue, whereas at the HCF location, the word BLUE is predictive for the response to the color green).
Although we do not deny that this low-level learning might contribute to the occurrence of the CSPC effect to some extent, we think that its influence is rather weak in the present design. First of all, specifically for location-based CSPC paradigms, it has been shown that the congruence proportion manipulation transfers to contingency-unbiased items that are presented with equal congruence probability at each location (Crump & Milliken, 2009), a finding that cannot be explained by contingency learning and thus demonstrates the involvement of cognitive control. Second, contingency learning applies especially when small sets of stimuli are used in an experimental design (Bugg & Hutchison, 2012). According to the contingency learning account by Schmidt and Besner (2008) proportion congruency effects are explained via the predictability of the response based on repetition confounds of stimulus features. For a two-item set, these response predictions for each stimulus–response combination are easy to apply by the participants, therefore, Bugg and Hutchison (2012) argue that smaller set sizes promote the reliance on contingency learning. In addition, they demonstrated that the effect vanishes for larger-item sets as they found no evidence of contingency learning within a four-item set. With regard to the present dual-task paradigm the response predictions for T1 and T2 would have to be made for thirty-two different context-stimulus–response compounds (i.e., a thirty-two-item set). Although, this is not impossible, we think that learning the prediction of two responses by linking the likelihood of two stimuli appearing together at a certain location is unlikely to solely account for the present findings.
It should be noted that the present findings represent a mere first step towards understanding the mechanisms that are or are not responsible for the activation of context-associated control sets. It is still conceivable, for example, that other location-based CSPC paradigms might offer more potent effects of abrupt stimulus onsets. The present paradigm was closely modeled after Fischer et al. (2014). Attentional effects of abrupt onsets might be stronger when, for example, no spatial placeholders and only single stimuli instead of dual tasks are used (e.g., Crump & Milliken, 2009; Lehle & Hübner, 2008).
To conclude, although previous studies demonstrated that endogenous advance allocation of attention towards a context feature (i.e., location) in a CSPC paradigm can facilitate the activation of a context-control link (Fischer et al., 2014), in the present study we could show that the exogenous disambiguation of the context feature by the abrupt onset of task-irrelevant stimuli at the wrong context (i.e., location) does not impact on the occurrence of the CSPC effect. Based on the present results, we suggest that to activate a context-associated attentional control set, the identification of the relevant stimulus at the respective context appears necessary to prompt the activation of the context-associated attentional control set.
Notes
Note, that for the study of RC compatibility effects the performance of Task 1 is the primary dependent measure. Performance in Task 2 is theoretically less important but is typically reported to control for trade-offs between tasks.
In a pilot study with equal font size, participants spontaneously reported that letters appeared larger in size than digits. This might be due to a wider horizontal extension of letters compared to digits. Therefore, we adjusted the size of letters to obtain a visually comparable impression.
It is conceivable, for example, that the requirement of re-orientation leads to a particular strong activation of the correct attentional control set, which might explain an increased CSPC effect in error rates of Block 2 in Experiment 3 when task-irrelevant stimuli appear first. We thank an anonymous reviewer for mentioning this possibility.
For the first block of Experiment 2, the CSPC effect (40 ms) was not significant, F(1, 19) = 1.68, p = .211 η 2p = .08, which might be due to a power problem.
References
Blais, C., Harris, M. B., Guerrero, J. V., & Bunge, S. A. (2012). Rethinking the role of automaticity in cognitive control. The Quarterly Journal of Experimental Psychology, 65(2), 268–276. doi:10.1080/17470211003775234922676200.
Blais, C., Robidoux, S., Risko, E. F., & Besner, D. (2007). Item-specific adaptation and the conflict-monitoring hypothesis: a computational model. Psychological Review, 114(4), 1076–1086. doi:10.1037/0033-295X.114.4.1076.
Bugg, J. M. (2014a). Conflict-triggered top-down control: default mode, last resort, or no such thing? Journal of Experimental Psychology: Learning, Memory, and Cognition, 40(2), 567.
Bugg, J. M. (2014b). Evidence for the sparing of reactive cognitive control with age. Psychology and Aging, 29(1), 115.
Bugg, J. M., & Crump, M. J. (2012). In support of a distinction between voluntary and stimulus-driven control: a review of the literature on proportion congruent effects. Frontiers in Psychology, 3, 367. doi:10.3389/fpsyg.2012.00367.
Bugg, J. M., & Hutchison, K. A. (2012). Converging evidence for control of color-word Stroop Interference at the item level. Journal of Experimental Psychology: Human Perception and Performance,. doi:10.1037/a0029145.
Bugg, J. M., Jacoby, L. L., & Chanani, S. (2011). Why it is too early to lose control in accounts of item-specific proportion congruency effects. Journal of Experimental Psychology: Human Perception and Performance, 37(3), 844. doi:10.1037/a0019957-2010-16967-001.
Bugg, J. M., Jacoby, L. L., & Toth, J. P. (2008). Multiple levels of control in the Stroop task. Memory & Cognition, 36(8), 1484–1494. doi:10.3758/MC.36.8.148436/8/1484.
Cañadas, E., Rodríguez-Bailón, R., Milliken, B., & Lupiáñez, J. (2013). Social categories as a context for the allocation of attentional control. Journal of Experimental Psychology: General, 142(3), 934–943. doi:10.1037/a0029794-2012-22280-001.
Corballis, P. M., & Gratton, G. (2003). Independent control of processing strategies for different locations in the visual field. Biological Psychology, 64(1–2), 191–209.
Crump, M. J., Gong, Z., & Milliken, B. (2006). The context-specific proportion congruent Stroop effect: location as a contextual cue. Psychonomic Bulletin & Review, 13(2), 316–321. doi:10.3758/Bf03193850.
Crump, M. J., & Milliken, B. (2009). The flexibility of context-specific control: evidence for context-driven generalization of item-specific control settings. The Quarterly Journal of Experimental Psychology, 62(8), 1523–1532. doi:10.1080/17470210902752096.
Crump, M. J., Vaquero, J. M., & Milliken, B. (2008). Context-specific learning and control: the roles of awareness, task relevance, and relative salience. Consciousness and Cognition, 17(1), 22–36. doi:10.1016/j.concog.2007.01.004.
Egner, T. (2014). Creatures of habit (and control): a multi-level learning perspective on the modulation of congruency effects. Frontiers in psychology,. doi:10.3389/fpsyg.2014.01247.
Fischer, R., & Dreisbach, G. (2015). Predicting high levels of multitasking reduces between-tasks interactions. Journal of Experimental Psychology: Human Perception and Performance, 41(6), 1482–1487. doi:10.1037/xhp0000157-2015-47211-001.
Fischer, R., Gottschalk, C., & Dreisbach, G. (2014). Context-sensitive adjustment of cognitive control in dual-task performance. Journal of Experimental Psychology. Learning, Memory, and Cognition, 40(2), 399–416. doi:10.1037/a0034310-2013-33714-001.
Fischer, R., & Hagendorf, H. (2006). The control of visual attention and its influence on prioritized processing in a location negative priming paradigm. Psychological Research, 70(4), 317–335. doi:10.1007/s00426-005-0220-2.
Fischer, R., Miller, J., & Schubert, T. (2007). Evidence for parallel semantic memory retrieval in dual tasks. Memory & Cognition, 35(7), 1685–1699.
Ghinescu, R., Schachtman, T. R., Stadler, M. A., Fabiani, M., & Gratton, G. (2010). Strategic behavior without awareness? Effects of implicit learning in the Eriksen flanker paradigm. Memory & Cognition, 38(2), 197–205. doi:10.3758/MC.38.2.19738/2/197.
Heinemann, A., Kunde, W., & Kiesel, A. (2009). Context-specific prime-congruency effects: on the role of conscious stimulus representations for cognitive control. Consciousness and Cognition, 18(4), 966–976. doi:10.1016/j.concog.2009.08.009.
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). The theory of event coding (TEC): a framework for perception and action planning. Behavioral and Brain Sciences, 24(5), 849–877. doi:10.1017/S0140525x01000103.
Jacoby, L. L., Lindsay, D. S., & Hessels, S. (2003). Item-specific control of automatic processes: Stroop process dissociations. Psychonomic Bulletin & Review, 10(3), 638–644. doi:10.3758/Bf03196526.
Kim, M. S., & Cave, K. R. (1999). Top-down and bottom-up attentional control: on the nature of interference from a salient distractor. Perception and Psychophysics, 61(6), 1009–1023.
King, J. A., Korb, F. M., & Egner, T. (2012). Priming of control: implicit contextual cuing of top-down attentional set. Journal of Neuroscience, 32(24), 8192–8200. doi:10.1523/JNEUROSCI.0934-12.2012.
Lehle, C., & Hübner, R. (2008). On-the-fly adaptation of selectivity in the flanker task. Psychonomic Bulletin & Review, 15(4), 814–818. doi:10.3758/PBR.15.4.814.
Lien, M. C., & Proctor, R. W. (2002). Stimulus-response compatibility and psychological refractory period effects: implications for response selection. Psychonomic Bulletin & Review, 9(2), 212–238. doi:10.3758/Bf03196277.
Logan, G. D. (1998). What is learned during automatization? II. Obligatory encoding of spatial location. Journal of Experimental Psychology: Human Perception and Performance, 24(6), 1720. doi:10.1037/0096-1523.24.6.1720.
Logan, G. D., & Schulkind, M. D. (2000). Parallel memory retrieval in dual-task situations: i. Semantic memory. Journal of Experimental Psychology: Human Perception and Performance, 26(3), 1072. doi:10.1037//0096-1523.26.3.1072.
Mayr, U. (1996). Spatial attention and implicit sequence learning: evidence for independent learning of spatial and nonspatial sequences. Journal of Experimental Psychology. Learning, Memory, and Cognition, 22(2), 350. doi:10.1037/0278-7393.22.2.350.
Mayr, U., & Bryck, R. L. (2007). Outsourcing control to the environment: effects of stimulus/response locations on task selection. Psychological Research, 71(1), 107–116. doi:10.1007/s00426-005-0039-x.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24(1), 167–202. doi:10.1146/annurev.neuro.24.1.16724/1/167.
Reder, L. M., Weber, K., Shang, J., & Vanyukov, P. M. (2003). The adaptive character of the attentional system: statistical sensitivity in a target localization task. Journal of Experimental Psychology: Human Perception and Performance, 29(3), 631–649.
Reuss, H., Desender, K., Kiesel, A., & Kunde, W. (2014). Unconscious conflicts in unconscious contexts: the role of awareness and timing in flexible conflict adaptation. Journal of Experimental Psychology: General, 143(4), 1701–1718. doi:10.1037/a0036437-2014-12079-001.
Schmidt, J. R. (2013). Questioning conflict adaptation: proportion congruent and Gratton effects reconsidered. Psychonomic Bulletin & Review, 20(4), 615–630. doi:10.3758/s13423-012-0373-0.
Schmidt, J. R., & Besner, D. (2008). The Stroop effect: why proportion congruent has nothing to do with congruency and everything to do with contingency. Journal of Experimental Psychology. Learning, Memory, and Cognition, 34(3), 514. doi:10.1037/0278-7393.34.3.514.
Schreij, D., Owens, C., & Theeuwes, J. (2008). Abrupt onsets capture attention independent of top-down control settings. Attention, Perception, & Psychophysics, 70(2), 208–218. doi:10.3758/Pp.70.2.208.
Scullin, M. K., Bugg, J. M., & McDaniel, M. A. (2012). Whoops, I did it again: commission errors in prospective memory. Psychology and Aging, 27(1), 46–53. doi:10.1037/a0026112-2011-25887-001.
Theeuwes, J. (1994). Endogenous and exogenous control of visual selection. Perception, 23(4), 429–440. doi:10.1068/P230429.
Theeuwes, J. (1995). Temporal and spatial characteristics of preattentive and attentive processing. Visual Cognition, 2(2–3), 221–233. doi:10.1080/13506289508401732.
Theeuwes, J. (2010). Top-down and bottom-up control of visual selection. Acta Psychologica, 135(2), 77–99. doi:10.1016/j.actpsy.2010.02.006.
Theeuwes, J., Atchley, P., & Kramer, A. F. (2000). On the time course of top-down and bottom-up control of visual attention (pp. 105–124). Control of Cognitive Processes: Attention and Performance XVIII.
Vietze, I., & Wendt, M. (2009). Context specificity of conflict frequency-dependent control. The Quarterly Journal of Experimental Psychology, 62(7), 1391–1400. doi:10.1080/17470210802426908906236510.
Walser, M., Fischer, R., & Goschke, T. (2012). The failure of deactivating intentions: aftereffects of completed intentions in the repeated prospective memory cue paradigm. Journal of Experimental Psychology. Learning, Memory, and Cognition, 38(4), 1030–1044. doi:10.1037/a0027000-2012-02245-001.
Walser, M., Goschke, T., & Fischer, R. (2014). The difficulty of letting go: moderators of the deactivation of completed intentions. Psychological Research, 78(4), 574–583. doi:10.1007/s00426-013-0509-5.
Wendt, M., Kluwe, R. H., & Vietze, I. (2008). Location-specific versus hemisphere-specific adaptation of processing selectivity. Psychonomic Bulletin & Review, 15(1), 135–140.
Wühr, P., & Kunde, W. (2008). Precueing spatial SR correspondence: is there regulation of expected response conflict? Journal of Experimental Psychology: Human Perception and Performance, 34(4), 872.
Yantis, S. (1993). Stimulus-driven attentional capture and attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 19(3), 676–681. doi:10.1037//0096-1523.19.3.676.
Yantis, S. (1998). Control of visual attention. In H. Pashler (Ed.), Attention (pp. 223–256). Hove: Psychology.
Yantis, S., & Hillstrom, A. P. (1994). Stimulus-driven attentional capture: evidence from equiluminant visual objects. Journal of Experimental Psychology: Human Perception and Performance, 20(1), 95. doi:10.1037/0096-1523.20.1.95.
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10(5), 601–621. doi:10.1037/0096-1523.10.5.601.
Yantis, S., & Jonides, J. (1990). Abrupt visual onsets and selective attention: voluntary versus automatic allocation. Journal of Experimental Psychology: Human Perception and Performance, 16(1), 121. doi:10.1037//0096-1523.16.1.121.
Acknowledgments
The authors are grateful to Julia Kleindienst and Marcel Kurtz for assistance in data collection and to Senne Braem and an anonymous reviewer for valuable comments on an earlier version of the manuscript. This research was supported by a Grant of the German Research Foundation (SFB 940, Project A3).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Gottschalk, C., Fischer, R. Activation of context-specific attentional control sets by exogenous allocation of visual attention to the context?. Psychological Research 81, 378–391 (2017). https://doi.org/10.1007/s00426-016-0746-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-016-0746-5