
Abstract
Purpose
In the present study, we examined how a 4-week intervention of maximal intended velocity (MIVRT) and controlled velocity resistance training (CRT)-induced task-specific responses in expert individuals.
Methods
Twenty elite athletes were randomly assigned to either a MIVRT (n = 10) or CRT (n = 10) group, both following the same volume–load training based on the back-squat three times a week but with different intentions in moving load (force–exertion speed). We assessed one-repetition maximum (1RM), mean propulsive velocity (MPV), and mean propulsive power (MPP) using a progressive-loading test before and after the intervention. A linear position transducer was used to monitor propulsive velocity in training and testing sessions.
Results
Both groups significantly increased their 1RM (CRT: + 12.3%, p < 0.001, d = 0.39; MIVRT: + 12.5%, p < 0.001, d = 0.45). Only the MIVRT group showed a significant improvement in MPV (p < 0.01) across different stepping loads, while both groups improved in MPP (MIVRT: + 22.4%, p < 0.001, d = 0.54; CRT: + 8.1%, p = 0.04, d = 0.17).
Conclusions
MIVRT induced significant adaptations in MPV and MPP at various loads (%1RM), underlining its specificity in targeting these parameters. Despite similar enhancements in 1RM, the distinct training protocols suggest that strength gains may stem from either maximal intent in moving loads or longer times under tension. This study highlights the role of execution speed in optimizing power outcomes, emphasizing task specificity as paramount to elicit physiological adaptations in chronically strength-trained individuals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The understanding of how resistance training (RT) impacts the neuromuscular system and its physiological effects on performance, muscle growth, and motor control has gained significant attention (Aagaard et al. 2002; Balshaw et al. 2017; Maffiuletti et al. 2016; Pearcey et al. 2021; Roberts et al. 2023; Škarabot et al. 2021). Adaptations explaining mentioned enhancements are linked to specific changes in motor unit behavior (Del Vecchio et al. 2019) that are at the base of various observations of increases in force and power following a strength training period (Bandy et al. 1990; Lopez et al. 2021; Sale 1988; Sarabia et al. 2017). These improvements have been characterized across different performance metrics, including countermovement jump, isometric leg press, maximal cycling power output, sprinting, and weightlifting (Ojanen et al. 2020; Seitz et al. 2014; Stone et al. 2005). Numerous studies have documented enhancements in one-repetition maximum (1RM), rate of force development (RFD), and mean propulsive velocity (MPV) in back-squat and bench-press exercises (González-Badillo et al. 2014; Morrissey et al. 1998; Pareja-Blanco et al. 2014; Sánchez-Medina et al. 2017). Notably, the speed of execution emerged as a crucial factor influencing force and power adaptations (González-Badillo et al. 2014; Riscart-Lopez et al. 2020; Sergio et al. 2007) in different multi-joint exercises, such as the back squat, Romanian deadlift, and bench press (Jones et al. 2001; Morin & Samozino 2016; Pérez-Castilla & García-Ramos 2020; Weakley et al. 2020). For instance, maximal intended velocity resistance training (MIVRT), consisting of performing tasks with the maximal intention of generating force (as fast as the load allows), has been demonstrated to induce broader adaptations compared to half-maximal velocity training (1RM, 18.2% vs 9.7%) in recreational athletes (González-Badillo et al. 2014; Pareja-Blanco et al. 2014). Accordingly, various studies have reported broader strength increases after a training protocol in the fast repetitions group compared to controlled repetitions (Ingebrigtsen et al. 2009; Maffiuletti & Martin 2001; Munn et al. 2005), whereas others observed no differences between these interventions (Fielding et al. 2002; Morrissey et al. 1998; Sergio et al. 2007). Given the enhancements in strength and power resulting from chronic exposure to a mechanical overload (Aagaard et al. 2002; Del Vecchio et al. 2019; Fielding et al. 2002; Fiorenza et al. 2020; Munn et al. 2005; Roberts et al. 2023), and that mentioned studies reported contrasting findings in recreationally active participants (González-Badillo et al. 2014; Pareja-Blanco et al. 2014), investigating specific adaptations to resistance training at different execution velocities in elite athletes covers the primary interest of the present study. To our knowledge, MIVRT protocols have been studied on elite populations, but no protocols have been done to compare task-dependent effects, which is the main concern in chronically strength-trained individuals (Włodarczyk et al. 2021).
To understand how these specific adaptations occur, we compared MIVRT and CRT adaptations over a 4-week intervention, as it reflects a mesocycle duration in most cases (Fleck 1999; Haugen et al. 2019; A. Turner 2011) as well as to be sufficient to induce significant adaptation of motor units underlying increase in muscle strength (Del Vecchio et al. 2019; Škarabot et al. 2021). We hypothesized that both protocols might induce significant increases in the maximal dynamic force as a result of the prolonged times under tension (TUT) for the CRT and the intention of moving loads for the MIVRT group (Behm & Sale 1993a; Burd et al. 2012; Rheese et al. 2021; Trybulski et al. 2022; Wilk et al. 2021). Based on observations made in recreationally active individuals (González-Badillo et al. 2014; Pareja-Blanco et al. 2014), we expected the MIVRT group to exhibit higher propulsive velocity across various stepping loads (Alcatraz et al. 2021), indicative of increased rate of force development (Del Vecchio et al. 2024). As a result, we awaited enhanced mean propulsive power (MPP) for both groups, reflecting the combined influence of a higher velocity and force production.
Methods
Participants and ethical statement
Twenty top-tier track and field (sprinting disciplines) and weightlifting athletes (males, n = 10; females, n = 10) were enrolled in the study. They competed at world-class (Top-20 world-rank 2021; score 1431–1250, national team athletes) or elite (Top 150 world-rank 2021; score 1250–1135, national team athletes) levels (McKay et al. 2022), with 2 or more years of experience with the back-squat exercise and overall resistance training. The participants were randomly assigned to either an MIVRT (males n = 5, females n = 5, BMI: 21.34 ± 1.96 kg∙m−2) or a CRT (males n = 5, females n = 5, 21.98 ± 4.36 kg∙m−2) group. Female participants were not under contraceptives (Burrows & Peters 2007; Elliott-Sale et al. 2020). All the athletes involved were between the first and second weeks of the general preparatory period for track and field or weightlifting and randomly distributed between the two intervention groups in equal sample size by adopting the block-randomization approach, ensuring equal number per group (Kang et al. 2008). Randomization was done to ensure the reliability of the intervention by minimizing bias and ensuring an equal distribution of male and female participants per group. Participants underwent no surgical intervention or significant injury in 6 months prior to the study that may have impaired the outcome at the testing time. All participants received detailed procedure information and signed a written informed consent. The study protocol observed and followed the Declaration of Helsinki and was approved by the Institutional Review Board of the University of Rome “Foro Italico” (CAR. 40/2020).
Experimental design
A longitudinal pre–post-design was used to compare the effect of two resistance training programs (MIVRT vs CRT) on the load–velocity–power parameters. Participants were asked to be available for 6 weeks, including twelve training and test sessions. In weeks 1 and 6, the 1RM test, MPV, MPP, and load–velocity–power profiles were assessed. Weeks 2–5 were dedicated to completing the training protocol. Before enrolling, participants were involved in the first weeks of the general preparatory period, after they underwent at least 2 weeks of rest from competitions and workouts during the transitional period. All participants were enrolled at the first stage of the macrocycle to minimize sport-specific bias and to guarantee a baseline preparation to avoid influences from previous mesocycles, which could interfere with expected outcomes, as well as to avoid any overreaching status, generally involving 2 weeks of rest to be recovered entirely (Halson & Jeukendrup 2004). In addition to the training protocol, participants performed 2/week aerobic sessions of 20-min cycling at a perceived intensity of RPE12 to RPE13 following existing guidelines (Stangier et al. 2016; Zinoubi et al. 2018) and 3/week core and general strength sessions comprising upper body strength exercises and flexibility. No additional lower-body strength training was performed. According to previous findings, female participants performed the tests during either the ovulatory or mid-luteal phase to reduce neuromuscular activity fluctuations and to avoid a documented general activation decrease attributed to the early follicular phase (Tenan et al. 2013; Weidauer et al. 2020), repeating the test in the same phase as the baseline (Lecce et al. 2024; Piasecki et al. 2024). All testing and training sessions took place under the supervision of the investigators, under constant environmental conditions (20 °C, 60% humidity), and at the same time of the day (± 1 h) for each participant (Racinais et al. 2005).
Force measurements and testing procedures
One week before testing, all athletes were involved in a 2-day familiarization period to prepare participants to perform the stepping load test. Familiarization days were separated by 48 –72 h, one from another. The measurements were performed in the first and sixth weeks within 5 days before the start and 5 days after the training protocol ended. The participants were asked to refrain from strenuous exercise 48 h before the testing sessions for pre- and post-assessment days (Lecce et al. 2023a), and no lower-body training was done to minimize possible residual fatigue. Moreover, the testing sessions were performed at the same time of the day to minimize diurnal variability in muscle force production (Lecce et al. 2023b; Racinais et al. 2005). The testing and training sessions were conducted using a non-guided barbell (free-weight setup) and a rack for the participants' safety. A reliable and validated [MPV: ICC = 0.99, CV = 1.24%; SEM = 0.01 m·s−1 (Martínez-Cava et al. 2020)] linear position transducer [Vitruve, SPEED4LIFTS S.L., Madrid, Spain (Callaghan et al. 2022)] was used to assess barbell propulsive velocity (Fig. 1A). The warm-up was standardized, including 5 min of isometric and dynamic core exercises, 5 min of mobility and dynamic flexibility, and four increasing loads for consecutive double repetitions per set (from RPE8 to RPE16) on back-squat (0 to 90° of knee flexion) (Escamilla & Krzyzewski 2001), with 3 min recovery. Subsequently, an encoder-monitored progressive-loading test was performed on back-squat exercise, setting an initial load of 20 kg and progressively increasing by 10 kg (if MPV > 0.8 m·s−1) or 5 kg (if MPV < 0.8 m·s−1) until the attained MPV was < 0.5 m·s−1, as previously suggested (Pareja-Blanco et al. 2014). Two repetitions were performed per each increasing set, and only two attempts were performed when the MPV > 0.5 m·s−1; otherwise, the test ended. Participants were instructed to 'move as fast as possible' at each repetition. Participants received visual and auditory feedback for execution velocity while performing each repetition and verbal encouragement in all conditions (Fig. 1C). Each stepping load was separated from the other by 3 min of recovery.

Experimental setup comprised a linear position transducer to assess the barbell velocity in each repetition of the back squat exercise (A). The testing sessions were completed before and after 4 weeks of intervention of either a maximal intended velocity (MIVRT) or controlled velocity (CRT) resistance training (3 sessions/week). Both groups performed the same volume load with different execution velocities (B). The progressive-loading test comprised a starting step at 20 kg with an increasing weight of 10 kg (if MPV was greater than 0.8 m∙s−1) or 5 kg (if MPV was lower than 0.8 m∙s−1) until a load considered appropriate to estimate 1RM [< 0.5 m∙s−1 (Pareja-Blanco et al. 2014)]
Data processing and analysis
Encoder data were extrapolated from the software as CSV files, inserted in a dataset, and analyzed. The individual load–velocity–power profiles were obtained, considering the highest MPV value for each load. Subsequently, the 1RM was estimated by linear regression equations (Alcatraz et al. 2021; Sánchez-Medina et al. 2017), setting 0.30 m·s−1 as the minimum velocity threshold as suggested in previous studies (Weakley et al. 2020). The mathematical calculations and the load–velocity–power profile did not consider MPV data obtained at 20 kg. The MPP was calculated by the product of the MPV by the corresponding force output converted stepping load. The normalization of 1RM and the peak MPP was used to calculate the percentage of increase after the intervention and to compare the changes in the MPV at the same %1RM as relative values.
Training protocol
The 4-week protocol comprised twelve supervised training sessions of MIVRT or CRT (based on the group), separated by 48–72 h. The warm-up was standardized for both groups, including 10 min of core exercises, 5 min of upper and lower body joint mobility, and 5 min of dynamic flexibility. Subsequently, four sets of two repetitions with incremental loading (20–70%1RM) were performed, and then the training protocol began. It comprised 2 sets of 5 reps at 60%1RM, 2 sets of 4 reps at 70%1RM, 2 sets of 3 reps at 80%1RM, and 2 sets of 2 reps at 90%1RM with a between-sets recovery of 3 min (Fig. 1B). To better isolate the velocity effect, both protocols were performed with the same volume load [sets x reps x weight (Guy Hornsby et al. 2018; Peterson et al. 2011)] but with different intentions of moving loads: the MIVRT group was asked to move as fast as possible in the concentric phase for every single repetition at any load (with the maximal intent), while the CRT group was asked to push the needed to match the half-maximal velocity of each stepping load (e.g., if participants' velocities at 80%1RM were 0.50 m·s−1, the max-velocity to match would be 0.25 m·s−1); this would ensure substantial differences in execution velocity to possibly induce task-specificity adaptations (González-Badillo et al. 2014). It is crucial to underscore that this also ensured a differential intention in moving load to induce adaptations confined to the speed of force exertion (Behm & Sale 1993a). Training sessions were monitored using the linear position transducer, and all participants received visual and auditory feedback in real time from a screen placed one and a half meters from their eyes. The eccentric phases of each repetition were performed at a controlled velocity for all participants (from 0.30 to 0.50 m·s−1). The between-sets recovery time was 3 min. Since CRT required longer TUT, the training protocol was planned as follows: volume ([2 × 5–2 × 4–2 × 3–2 × 2]/session), intensity (10 × 60%–8 × 70%–6 × 80%–4 × 90%), and recovery (3' between sets) were equal in both training groups (Baz-Valle et al. 2021). All participants successfully performed training protocols.
Statistical analysis
The data distribution normality was assessed using the Shapiro–Wilk test before conducting statistical comparisons. Multiple t tests were performed to account for between and within-group comparisons. Paired sample t tests were then employed to examine the influence of sex on the physiological responses to the different training protocols (pre–post). Since adaptations were not sex-dependent, male and female participants were also examined together to characterize the effect of training. Independent sample t tests were used to compare the age, height, mass, BMI, 1RM, and MPP between groups at baseline. Statistical differences for 1RM, MPP, and absolute MPV were assessed using paired sample t tests. Relative differences for sex-dependent and training-dependent responses were calculated for 1RM and MPP using the formula: ∆χ = [χ2–χ1], where χ–n represents the relative result (%) of a given variable at baseline (1) or after training (2). Relative differences were compared between MIVRT and CRT with independent-sample t tests. Effect sizes were determined using Cohen's d when the result was statistically significant, considering 0.2 = small effect. 0.5 = moderate effect. 0.8 = large effect (Fritz et al. 2012). Changes in load–velocity–power relationships were analyzed by comparing the slopes obtained through univariate linear regressions with one-way ANOVA (Andrade & Estévez-Pérez 2014). To account for the reliability and consistency of comparisons, two-way mixed effects, consistency and single measurement intraclass correlation coefficients (ICC3,1) were computed for 1RM, MPP, and MPV. SPSS, version 25–0 (IBM Corp., Armonk, NY, USA) was used for all statistical analyses. A p < 0.05 was considered statistically significant. Data are presented as the mean ± SD.
Results
Between-group differences
No between-group differences were found in the anthropometric characteristics, one-repetition maximum, and mean power output (p > 0.05). Baseline comparisons are reported in Table 1.
1RM and MPP results
After 4 weeks of training, the 1RM significantly increased for both groups [MIVRT: from 112.90 ± 29.15 kg to 126.80 ± 31.92 kg (+ 12.5%), ICC3,1 = 0.98 [0.93–0.99], p < 0.001, d = 0.45, Fig. 2A; CRT: from 115.50 ± 33.97 kg to 129.90 ± 38.92 kg (+ 12.3%), ICC3,1 = 0.98, [0.94–0.99], p < 0.001, d = 0.39, Fig. 2B]. No significant effects emerged by comparing the relative differences in the 1RM of groups (p = 0.891, Fig. 2C).

Adaptations in 1RM induced by 4 weeks of MIVRT A and CRT B intervention are displayed as bar plots reporting individual results (black circles). The relative comparisons are displayed as relative differences with bar plots (C). Each filled circle represents individual change percentages from the baseline for MIVRT (white bars) and CRT (grey bars). Data are reported as the mean ± SD; p < 0.001 ***
Male and female participants showed a significant increase in 1RM after 4 weeks of MIVRT [M, from 135.80 ± 17.77 kg to 154.40 ± 9.60 kg (+ 14%), p = 0.01, d = 1.30; F: from 90.01 ± 16.87 kg to 99.20 ± 17.40 kg (+ 10%), p = 0.02, d = 0.53] and CRT [M, from 145.40 ± 17.70 kg to 163.60 ± 21.08 kg (+ 12.5%), p = 0.005, d = 0.93; F: from 85.60 ± 6.95 kg to 96.20 ± 11.16 kg (+ 12.3%), p = 0.01, d = 1.14], with similar results in the relative differences (MIVRT, p = 0.403; CRT, p = 0.841), underlining the null-influence of sex for the observed adaptations, Fig. 3.

1RM adaptations for male and female participants are displayed for MIVRT (A) and CRT (C) as bar plots reporting individual values (black circles). Relative differences are reported for both the MIVRT (B) and CRT (D), displaying comparisons between male (white bars) and female (grey bars) results. Each filled circle represents the individual relative change from the baseline. Data are reported as the mean ± SD. p < 0.05 *, p < 0.01 **
The MPP significantly increased for both groups [MIVRT from 534.60 ± 200.40 W to 652.70 ± 232.36 W (+ 22.4%), ICC3,1 = 0.96 [0.88–0.99], p < 0.001, d = 0.54, Fig. 4A; CRT from 544.90 ± 239.87 W to 589.40 ± 270.39 W (+ 8.1%), ICC3,1 = 0.97 [0.92–0.99], p = 0.03, d = 0.17, Fig. 4B]. A significant difference emerged from the between-group comparison of the relative post-training differences (p < 0.001, d = 1.95, Fig. 4C).

Adaptations in MPP induced by 4 weeks of MIVRT (A) and CRT (B) intervention are displayed as bar plots reporting individual results (black circles). The relative comparisons are displayed as relative differences with bar plots (C). Each filled circle represents individual change percentages from the baseline for MIVRT (white bars) and CRT (grey bars). Data are reported as the mean ± SD; p < 0.05 *, p < 0.001 ***
Male and female participants showed a significant increase in MPP in both the MIVRT [M, from 690.82 ± 149.80 W to 831.24 ± 156.90 W (+ 20.5%), p = 0.001, d = 0.91; F: from 378.47 ± 83.29 W to 474.23 ± 131.21 W (+ 24.4%), p = 0.01, d = 0.87] and CRT [M, from 749.42 ± 123.71 W to 818.42 ± 152.98 W (+ 9.2%), p = 0.03, d = 0.49; F: from 340.43 ± 97.35 W to 360.40 ± 99.29 W (+ 5.9%), p = 0.04, d = 0.20], with similar relative differences (MIVRT, p = 0.587; CRT, p = 0.491), underlining the null-influence of sex for the observed adaptations, Fig. 5.

MPP adaptations for male and female participants are displayed for MIVRT (A) and CRT (C) as bar plots reporting individual values (black circles). Relative differences are reported for both the MIVRT (B) and CRT (D), displaying comparisons between male (white bars) and female (grey bars) results. Each filled circle represents the individual relative change from the baseline. Data are reported as the mean ± SD. p < 0.05 *, p < 0.001 ***
MPV results
MPV values were compared at 90%, 80%, 70%, 60%, and 50%1RM (Fig. 6). No differences were observed in the CRT group across all the MPVs analyzed (p > 0.05). Conversely, the MIVRT group showed a significant increase in the MPV at 90%1RM [from 0.42 ± 0.03 m·s−1 to 0.45 ± 0.03 m·s−1 (ICC3,1 = 0.92 [0.75–0.98], p = 0.003, d = 1.01)], 80%1RM [from 0.54 ± 0.06 m·s−1 to 0.59 ± 0.06 m·s−1 (ICC3,1 = 0.92 [0.76–0.98], p = 0.001, d = 0.83)], 70%1RM [from 0.66 ± 0.09 m·s−1 to 0.73 ± 0.09 m·s−1 (ICC3,1 = 0.93 [0.77–0.98], p = 0.001, d = 0.77)], 60%1RM [from 0.78 ± 0.12 m·s−1 to 0.87 ± 0.12 m·s−1 (ICC3,1 = 0.92 [0.75–0.98], p = 0.002, d = 0.74)], and 50%1RM [from 0.90 ± 0.16 m·s−1 to 1.02 ± 0.15 m·s−1 (ICC3,1 = 0.93 [0.77–0.98], p = 0.001, d = 0.77)].

Adaptations in MPV at 50–60–70–80–90% 1RM are displayed as bar plots with individual values (black circles) for MIVRT (A) and CRT (B) groups reporting the baseline (white bars) and post-training (grey bars). Data are reported as the mean ± SD; p < 0.01 **
Load–velocity–power relationships
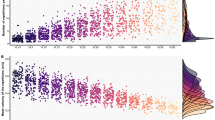
Changes in the load–velocity relationship slope occurred for CRT (p = 0.001) but not MIVRT (p = 0.248, Fig. 7C), with significant differences observed for the intercept in both groups (p < 0.001, Fig. 7A-–D). Changes in the velocity–power relationship slopes occurred for the CRT (p = 0.02) but not the MIVRT (p = 0.143, Fig. 7F) group; both groups showed significant differences in the intercept (p < 0.01, Fig. 7B-–E).

Load–velocity profiles are displayed as scatter plots comparing the baseline to responses induced by MIVRT (A) and CRT (B). Similarly, power–velocity profiles are reported for MIVRT (D) and CRT (E) groups. In each plot, markers for single participants' velocity across the whole load spectrum are expressed as a ratio of PRE. Load interval with the related MPV is 5%RM each starting from zero. Bar plots of the differences in the slope of the load–velocity (C) and power–velocity (F) relationships are displayed as bar plots displaying individual slopes. Data are reported as the mean ± SD; p < 0.05 *, p < 0.01 **
Discussion
In the present study, we investigated the effects of two training protocols that were equal in volume load but different in the speed at which exercises were performed. Both the maximal intended velocity and controlled velocity resistance training protocols induced significant increases in strength and power parameters. We found that male and female participants responded similarly to resistance training, showing increased strength and power to a similar extent (within-group comparison). These findings underlined that the significant effects observed depended not on sex but on task-specific responses. In addition, although significant effects have been highlighted in previous studies concerning MIVRT effects on an elite population, we demonstrated that diverse resistance training protocols induced differential responses in chronically strength-trained individuals according to task specificity.
Various parameters likely influence the similar enhancements observed in the maximal dynamic force. Indeed, controlled velocity resistance training is associated with a longer cumulative time under tension (Pareja-Blanco et al. 2014; Trybulski et al. 2022), which has been demonstrated to significantly affect strength adaptations even in the short-term (4 weeks) (Handford et al. 2022). On the other hand, exerting force with the maximal intended speed has been hypothesized to induce specific neuromuscular responses due to its association with a higher rate of activation compared to controlled velocity contractions across a broad range of intensities (10–90%1RM) (Tøien et al. 2022). These results underlined that both the maximal intent of moving loads and achieving prolonged times under tension may lead to similar increases in force output in chronically strength-trained individuals. Furthermore, achieving longer TUTs could also promote different cellular signaling pathways, inducing broader mitochondrial and protein synthesis (Burd et al. 2012), considering that both prolonged TUTs and high overload are crucial for enhancing muscle strength and hypertrophy (Roberts et al. 2023).
Exerting force at the maximal speed induced significant enhancements in the MPV across various %1RM, confirming previous observations examining the influence of stimulus specificity on the propulsive velocity improvement (González-Badillo et al. 2014). Nevertheless, higher contraction velocities are associated with rapid force increase driven by both a greater neural drive to muscle and rate of muscle activation, which likely explains the specific responses found after the MIVRT (Del Vecchio et al. 2018, 2024; Tøien et al. 2022). Indeed, the distinct stimuli likely led to differential responses as it has been reported how controlled contractions induce modifications in the peripheral membrane properties (e.g., M-wave shape), whereas ballistic contractions affect contractile properties (Maffiuletti & Martin 2001). It is also necessary to consider that the differential intention of moving load (i.e., exerting force) is determined by distinct training protocols supporting previous evidence (non-expert population) in which the intention in moving load covers the paramount drive for specific adaptations, also in chronically strength-trained individuals (Behm & Sale 1993b). In addition, it is known that the actual difference in the intention in moving load may be the pivotal component of such outcomes and that training strategies have been used only to account for these distinctions in intended movement speed (Behm & Sale 1993a; González-Badillo et al. 2014). These results suggest that the increase in the execution velocity may stem from specific adaptations to resistance training, which could be intended as the primary drive for specific adaptations in power training.
As a result of the combined influence of increased velocity and force production, both groups improved their maximal propulsive power in the back squat. Nevertheless, the greater enhancement observed after the maximal-intended velocity resistance training underscores its specificity for power training. This is also evidenced by the increase in the mean propulsive velocity at relative loads, which was absent after the intervention based on controlled velocity contraction. Since a greater speed of motor unit recruitment and activation governs enhancements in the rate of force development as observed following interventions combining ballistic and sustained contractions (Del Vecchio et al. 2018, 2019, 2024; Škarabot et al. 2021), it is possible that exerting force at the maximal speed may have induced a greater adaptation toward both strength and velocity parameters. Again, the broader increase in mechanical power output for the MIVRT group may be attributed to the maximal intention of moving loads as a result of a significant increase in the MPV (Behm & Sale 1993a; González-Badillo et al. 2014; Rheese et al. 2021; Weakley et al. 2023). It is also plausible that both greater contraction velocity and muscle activation associated with maximal intention in moving loads contributed to broader inhibitory effects on antagonist muscles during the concentric phase, potentially explaining the present results (Behm & Sale 1993a; Carolan & Cafarelli 1992; Häkkinen et al. 1998; Tillin et al. 2011).
The observed adaptations led to significant changes in the load–velocity profiles of participants in both groups, with the MIVRT group showing significant effects for intercept and CRT showing significant differences for slope and intercept. This accounts for the association between resistance training specificity and mechanical-related output (Del Vecchio et al. 2024). The MIVRT group significantly improved strength and velocity parameters, whereas the CRT group only for 1RM. As a consequence, a positive shift in mean propulsive velocity was observed solely in the MIVRT group, while the CRT group showed a slight, non-significant backward shift towards lower MPV values. Furthermore, both groups displayed significant changes in the power–velocity profile due to improvements in 1RM (MIVRT and CRT) and MPV (MIVRT), with significant variations of intercept observed in power output for both interventions. However, it is crucial to notice how to manage this information during periodization based on individual necessity (Fleck 1999; A. Turner 2011). By systematically testing athletes, changes occurring in the load–velocity–power profiles could help understand whether specific stimuli accomplish specific needs (Banyard et al. 2018; Pérez-Castilla et al. 2022). Additionally, it is known that load–velocity profiles differ according to strength level (Torrejón et al. 2019); however, we demonstrate that specific stimuli induced expected adaptations (hypothesized based on non-elite data) in elite populations (González-Badillo et al. 2014; Pérez-Castilla et al. 2022). Thus, using specific stimuli may be a helpful tool for enhancing individual athlete performance by assessing load–velocity profile before and after intervention (Banyard et al. 2018; Pérez-Castilla & García-Ramos 2020).
The effects on strength may also depend on the population involved and the periodization stage. Considering their performance maturity and that greater neural effects seem to be attained in the early resistance training phase (4–8 weeks) (Del Vecchio et al. 2024; Hughes et al. 2018; Sale 1988), world-class and elite athletes require more varied stimuli to increase their neuromuscular parameters compared to recreationally active individuals (Hughes et al. 2018; McKay et al. 2022). This differential response is evident when comparing these populations in terms of response timing and performance-planning strategies (Haugen et al. 2019), suggesting that training specificity plays a crucial role in influencing adaptations, as indicated by the similar increase in 1RM in the current results. As expected, by comparing effect sizes to the only study adopting a similar experimental setup as ours, we observed a comparable trend for 1RM and MPV between groups. However, there were effect sizes for 1RM (both medium) but greater effect sizes for MPV (large effect) in recreationally active individuals (see H. M., I. Turner & Bernard, (2006) for different effect size comparisons). No data were provided for MPP to compare.
Indeed, our results support the hypothesis that neural components could significantly contribute to further adaptations to resistance training (Pearcey et al. 2021) and that these are highly sensitive to stimulus-specificity (i.e., speed of force exertion).
In summary, both maximal intended velocity and controlled-velocity resistance training significantly improve muscle strength and power, but with differential adaptations. The increase in 1RM and MPP observed following MIVRT is associated with a concurrent increase in the propulsive velocity. On the other hand, CRT implies prolonged time under tension, which has been observed with diverse mechanisms of strength enhancements supporting consistent execution velocity observed after intervention. These findings demonstrate how specific training protocols can target different aspects of muscle performance and highlight the value of combining methods for optimal strength and power development in athletes. This study could help us understand how different resistance training approaches affect neuromuscular adaptations and performance.
Future research is warranted to explore how the intention of moving loads may influence physiological responses to distinct resistance training approaches, including molecular and cellular mechanisms underlying the differential adaptations induced by MIVRT and CRT. Researchers could also understand the precise adaptation site by systematically assessing participants using advanced molecular biology techniques and biopsies. Such investigations deepen our understanding of muscle adaptation and inform targeted training strategies for optimizing athletic performance and specific rehabilitation protocols in clinical settings.
Practical applications
In the context of periodization, incorporating both maximal intended velocity and controlled-velocity resistance training offers distinct benefits across different training phases. MIVRT enhances muscle strength, maximal power output, and muscle shortening velocity (i.e., MPV), which is crucial for early stage adaptations or specific adaptations required by competition models. On the other hand, CRT, emphasizing prolonged time under tension, promotes hypertrophy and strength gains through mechanisms like mitochondrial and sarcoplasmic protein synthesis. Integrating these protocols sequentially within a periodized program optimizes both acute and delayed muscle responses, enhancing overall athletic performance and muscle adaptation over time.
Data availability
The data that support the findings of this study are available on request from the corresponding author.
Abbreviations
- 1RM:
-
One repetition maximum
- BMI:
-
Body-mass index
- CRT:
-
Controlled velocity resistance training
- MIVRT:
-
Maximal intended velocity resistance training
- MPP:
-
Mean propulsive power
- MPV:
-
Mean propulsive velocity
- RFD:
-
Rate of force development
- RPE:
-
Rate of perceived exertion
- RT:
-
Resistance training
- TUT:
-
Time under tension
References
Aagaard P, Simonsen EB, Andersen JL, Magnusson P, Dyhre-poulsen P, Dyhre-Poulsen P (2002) Increased rate of force development and neural drive of human skeletal muscle following resistance training. J Appl Physiol 93:1318–1326. https://doi.org/10.1152/japplphysiol.00283.2002.-The
Alcatraz J, Parejia-Blanco F, Rodriguez-Lopez C, Navarro-Cruz R, Cornejo-Daza P, Ara I, Alegre L (2021) Comparison of linear, hyperbolic and double-hyperbolic models to assess the force-velocity relationship in multi-joint exercises. Eur J Sport Sci 21(3):359–369
Andrade JM, Estévez-Pérez MG (2014) Statistical comparison of the slopes of two regression lines: A tutorial. In Analytica Chimica Acta 838:1–12. https://doi.org/10.1016/j.aca.2014.04.057
Balshaw TG, Massey GJ, Maden-Wilkinson TM, Morales-Artacho AJ, McKeown A, Appleby CL, Folland JP (2017) Changes in agonist neural drive, hypertrophy and pre-training strength all contribute to the individual strength gains after resistance training. Eur J Appl Physiol 117(4):631–640. https://doi.org/10.1007/s00421-017-3560-x
Bandy WD, Lovelace-Chandler V, Mckitrick-Bandy B (1990) Adaptation of skeletal muscle to resistance training. The J Orthop Sports Phys Ther 12(6):248–255
Banyard HG, Nosaka K, Vernon AD, Gregory Haff G (2018) The reliability of individualized load–velocity profiles. Int J Sports Physiol Performance 13(6):763–769. https://doi.org/10.1123/ijspp.2017-0610
Baz-Valle E, Fontes-Villalba M, Santos-Concejero J (2021) Total number of sets as a training volume quantification method for muscle hypertrophy: a systematic review. In J Strength Condition Res. https://doi.org/10.1519/JSC.0000000000002776
Behm DG, Sale DG (1993a) Intended rather than actual movement velocity determines velocity-specific training response. J Appl Physiol 74(1):359–368. https://doi.org/10.1152/jappl.1993.74.1.359
Behm DG, Sale DG (1993b) Velocity specificity of resistance training. In Sports Med: Evaluations Res Exerc Sci Sports Med 15(6):374–388. https://doi.org/10.2165/00007256-199315060-00003
Burd NA, Andrews RJ, West DWD, Little JP, Cochran AJR, Hector AJ, Cashaback JGA, Gibala MJ, Potvin JR, Baker SK, Phillips SM (2012) Muscle time under tension during resistance exercise stimulates differential muscle protein sub-fractional synthetic responses in men. J Physiol 590(2):351–362. https://doi.org/10.1113/jphysiol.2011.221200
Burrows M, Peters CE (2007) The influence of oral contraceptives on athletic performance in female athletes. In Sports Med 37(7):557–574. https://doi.org/10.2165/00007256-200737070-00001
Callaghan DE, Guy JH, Elsworthy N, Kean C (2022) Validity of the PUSH band 2.0 and Speed4lifts to measure velocity during upper and lower body free-weight resistance exercises. J Sports Sci 40(9):968–975. https://doi.org/10.1080/02640414.2022.2043629
Carolan B, Cafarelli E (1992) Adaptations in coactivation after isometric resistance training. J Appl Physiol 73(3):911–917. https://doi.org/10.1152/jappl.1992.73.3.911
Del Vecchio A, Negro F, Falla D, Bazzucchi I, Farina D, Felici F (2018) Higher muscle fiber conduction velocity and early rate of torque development in chronically strength-trained individuals. J Appl Physiol 125:1218–1226. https://doi.org/10.1152/japplphysiol.00025.2018.-Strength-trained
Del Vecchio A, Casolo A, Negro F, Scorcelletti M, Bazzucchi I, Enoka R, Felici F, Farina D (2019) The increase in muscle force after 4 weeks of strength training is mediated by adaptations in motor unit recruitment and rate coding. J Physiol 597(7):1873–1887
Del Vecchio A, Enoka RM, Farina D (2024) Specificity of early motor unit adaptations with resistive exercise training. J Physiol 602(12):2679–2688. https://doi.org/10.1113/JP282560
Elliott-Sale KJ, McNulty KL, Ansdell P, Goodall S, Hicks KM, Thomas K, Swinton PA, Dolan E (2020) The effects of oral contraceptives on exercise performance in women: a systematic review and meta-analysis. In Sports Med 50(10):1785–1812. https://doi.org/10.1007/s40279-020-01317-5
Escamilla RF, Krzyzewski MW (2001) Knee biomechanics of the dynamic squat exercise. In Med Sci Sports Exerc 33(1):127–141
Fielding RA, LeBrasseur NK, Cuoco A, Bean J, Mizer K, Fiatarone Singh MA (2002) High-velocity resistance training increases skeletal muscle peak power in older women. J Am Geriatr Soc 50(4):655–662. https://doi.org/10.1046/j.1532-5415.2002.50159.x
Fiorenza M, Gliemann L, Brandt N, Bangsbo J (2020) Hormetic modulation of angiogenic factors by exercise-induced mechanical and metabolic stress in human skeletal muscle. Am J Physiol Heart Circ Physiol 319:824–834. https://doi.org/10.1152/ajpheart.00432.2020.-This
Fleck SJ (1999) Periodized strength training: a critical review. J Strength Conditioning Res 13(1). https://doi.org/10.1519/1533-4287(1999)013<0082:PSTACR>2.0.CO;2
Fritz C, Morris P, Richer J (2012) Effect size estimates: current use, calculations, and interpretation. J Exp Psychol Gen 141(1):2–18
González-Badillo JJ, Rodríguez-Rosell D, Sánchez-Medina L, Gorostiaga EM, Pareja-Blanco F (2014) Maximal intended velocity training induces greater gains in bench press performance than deliberately slower half-velocity training. Eur J Sport Sci 14(8):772–781. https://doi.org/10.1080/17461391.2014.905987
Guy Hornsby W, Gentles JA, Comfort P, Suchomel TJ, Mizuguchi S, Stone MH (2018) Resistance training volume load with and without exercise displacement. Sports 6(4):137. https://doi.org/10.3390/sports6040137
Häkkinen K, Kallinen M, Izquierdo M, Jokelainen K, Lassila H, Mälkiä E, Kraemer WJ, Newton RU, Alen M (1998) Changes in agonist-antagonist EMG, muscle CSA, and force during strength training in middle-aged and older people. J Appl Physiol 84(4):1341–1349. https://doi.org/10.1152/jappl.1998.84.4.1341
Halson SL, Jeukendrup AE (2004) Does overtraining exist? An analysis of overreaching and overtraining research. In Sports Med 34(14):967–981. https://doi.org/10.2165/00007256-200434140-00003
Handford MJ, Bright TE, Mundy P, Lake J, Theis N, Hughes JD (2022) The need for eccentric speed: a narrative review of the effects of accelerated eccentric actions during resistance-based training. In Sports Med 52(9):2061–2083. https://doi.org/10.1007/s40279-022-01686-z
Haugen T, Seiler S, Sandbakk Ø, Tønnessen E (2019) The training and development of elite sprint performance: an integration of scientific and best practice literature. In Sports Med - Open. https://doi.org/10.1186/s40798-019-0221-0
Hughes DC, Ellefsen S, Baar K (2018) Adaptations to endurance and strength training. Cold Spring Harbor Perspectives Med 8(6):a029769. https://doi.org/10.1101/cshperspect.a029769
Ingebrigtsen J, Holtermann A, Roeleveld K (2009) Effects of load and contraction velocity during three-week biceps curls training on isometric and isokinetic performance. J Strength Condition Res 23(6):1670–1676
Jones K, Bishop P, Hunter G, Fleisig G (2001) The effects of varying resistance-training loads on intermediate- and high-velocity-specific adaptations. J Strength Cond Res 15(3):349–356
Kang M, Ragan BG, Park JH (2008) Issues in outcomes research: an overview of randomization techniques for clinical trials. In J Athletic Training 43(2):215–221. https://doi.org/10.4085/1062-6050-43.2.215
Lecce E, Nuccio S, Del Vecchio A, Conti A, Nicolò A, Sacchetti M, Felici F, Bazzucchi I (2023a) Sensorimotor integration is affected by acute whole-body vibration: a coherence study. Front Physiol. https://doi.org/10.3389/fphys.2023.1266085
Lecce E, Nuccio S, Del Vecchio A, Conti A, Nicolò A, Sacchetti M, Felici F, Bazzucchi I (2023b) The acute effects of whole-body vibration on motor unit recruitment and discharge properties. Front Physiol. https://doi.org/10.3389/fphys.2023.1124242
Lecce E, Conti A, Nuccio S, Felici F, Bazzucchi I (2024) Characterising sex-related differences in lower- and higher-threshold motor unit behaviour through high-density surface electromyography. Exp Physiol. https://doi.org/10.1113/EP091823
Lopez P, Radaelli R, Taaffe DR, Newton RU, Galvão DA, Trajano GS, Teodoro JL, Kraemer WJ, Häkkinen K, Pinto RS (2021) Resistance training load effects on muscle hypertrophy and strength gain: systematic review and network meta-analysis. In Med Sci Sports Exercise 53(6):1206–1216. https://doi.org/10.1249/MSS.0000000000002585
Maffiuletti NA, Martin A (2001) Progressive versus rapid rate of contraction during 7 wk of isometric resistance training. Med Sci Sports Exerc 33(7):1220–1227. https://doi.org/10.1097/00005768-200107000-00022
Maffiuletti NA, Aagaard P, Blazevich AJ, Folland J, Tillin N, Duchateau J (2016) Rate of force development: physiological and methodological considerations. In Eur J Appl Physiol 11(6):1091–1116. https://doi.org/10.1007/s00421-016-3346-6
Martínez-Cava A, Hernández-Belmonte A, Courel-Ibáñez J, Morán-Navarro R, González-Badillo JJ, Pallarés JG (2020) Reliability of technologies to measure the barbell velocity: Implications for monitoring resistance training. PLoS ONE 15(6):e0232465. https://doi.org/10.1371/journal.pone.0232465
McKay AKA, Stellingwerff T, Smith ES, Martin DT, Mujika I, Goosey-Tolfrey VL, Sheppard J, Burke LM (2022) Defining training and performance caliber: a participant classification framework. Int J Sports Physiol Perform 17(2):317–331. https://doi.org/10.1123/ijspp.2021-0451
Morin JB, Samozino P (2016) Interpreting power-force-velocity profiles for individualized and specific training. In Int J Sports Physiol Performance 11(2):267–272. https://doi.org/10.1123/ijspp.2015-0638
Morrissey MC, Harman EA, Frykman PN, Hoon Han K (1998) Early phase differential effects of slow and fast barbell squat training. The Am J Sports Med 26(2):221–230
Munn J, Herbert RD, Hancock MJ, Gandevia SC (2005) Resistance training for strength: Effect of number of sets and contraction speed. Med Sci Sports Exerc 37(9):1622–1626. https://doi.org/10.1249/01.mss.0000177583.41245.f8
Ojanen T, Kyröläinen H, Kozharskaya E, Häkkinen K (2020) Changes in strength and power performance and serum hormone concentrations during 12 weeks of task-specific or strength training in conscripts. Physiol Rep 8(9):e14422
Pareja-Blanco F, Rodríguez-Rosell D, Sánchez-Medina L, Gorostiaga EM, González-Badillo JJ (2014) Effect of movement velocity during resistance training on neuromuscular performance. Int J Sports Med 35(11):916–924. https://doi.org/10.1055/s-0033-1363985
Pearcey GEP, Alizedah S, Power KE, Button DC (2021) Chronic resistance training: is it time to rethink the time course of neural contributions to strength gain? In Eur J Appl Physiol 121(9):2413–2422. https://doi.org/10.1007/s00421-021-04730-4
Pérez-Castilla A, García-Ramos A (2020) Changes in the load-velocity profile following power and strength-oriented resistance-training programs. Int J Sports Physiol Perform 15(10):1460–1466. https://doi.org/10.1123/ijspp.2019-0840
Pérez-Castilla A, Jukic I, Janicijevic D, Akyildiz Z, Senturk D, García-Ramos A (2022) Load-velocity relationship variables to assess the maximal neuromuscular capacities during the back-squat exercise. Sports Health 14(6):885–893. https://doi.org/10.1177/19417381211064603
Peterson MD, Pistilli E, Haff GG, Hoffman EP, Gordon PM (2011) Progression of volume load and muscular adaptation during resistance exercise. Eur J Appl Physiol 111(6):1063–1071. https://doi.org/10.1007/s00421-010-1735-9
Piasecki J, Škarabot J, Spillane P, Piasecki M, Ansdell P (2024) Sex differences in neuromuscular ageing: the role of sex hormones. Exerc Sport Sci Rev. https://doi.org/10.1249/JES.0000000000000335
Racinais S, Blonc S, Jonville S, Hue O (2005) Time of day influences the environmental effects on muscle force and contractility. Med Sci Sports Exerc 37(2):256–261. https://doi.org/10.1249/01.MSS.0000149885.82163.9F
Rheese M, Drinkwater EJ, Leung H, Andrushko JW, Tober J, Hendy AM (2021) The effects of verbal cueing for high intended movement velocity on power, neuromuscular activation, and performance. Scandinavian J Med Sci Sports 31(5):1115–1125. https://doi.org/10.1111/sms.13926
Riscart-Lopez J, Rendeiro-Pinho G, Mil-Homens P, Soares-daCosta R, Loturco I, Pareja-Blanco F, Léon-Prados JA (2020) Effects of four different velocity-based training programming models on strength gains and physical performance. J Strength Conditioning Res 35(3):596–603
Roberts MD, McCarthy JJ, Hornberger TA, Phillips SM, Mackey AL, Nader GA, Boppart MD, Kavazis AN, Reidy PT, Ogasawara R, Libardi CA, Ugrinowitsch C, Booth FW, Esser KA (2023) Mechanisms of mechanical overload-induced skeletal muscle hypertrophy: current understanding and future directions. In Physiol Rev 103(4):2679–2757. https://doi.org/10.1152/physrev.00039.2022
Sale D (1988) Neural adaptation to resistance training. Med Sci Sports Exerc 20(5):135–145
Sánchez-Medina L, Pallarés J, Pérez C, Morán-Navarro R, González-Badillo J (2017) Estimation of relative load from bar velocity in the full back squat exercise. Sports Med Int Open 01(02):E80–E88. https://doi.org/10.1055/s-0043-102933
Sarabia JM, Moya-Ramón M, Hernández-Davó JL, Fernandez-Fernandez J, Sabido R (2017) The effects of training with loads that maximise power output and individualised repetitions vs. traditional power training. PLoS ONE 12(10):e0186601. https://doi.org/10.1371/journal.pone.0186601
Seitz LB, Reyes A, Tran TT, de Villarreal ES, Haff GG (2014) Increases in lower-body strength transfer positively to sprint performance: a systematic review with meta-analysis. In Sports Med 44(12):1693–1702. https://doi.org/10.1007/s40279-014-0227-1
Sergio P, Gomes C, Inez M, Pereira R (2007) Effects of isotonic resistance training at two movement velocities on strength gains. In Rev Bras Med Esporte 13(2):91–96
Škarabot J, Brownstein CG, Casolo A, Del Vecchio A, Ansdell P (2021) The knowns and unknowns of neural adaptations to resistance training. In Eur J Appl Physiol 121(3):675–685. https://doi.org/10.1007/s00421-020-04567-3
Stangier C, Abel T, Mierau J, Hollmann W, Strüder HK (2016) Effects of cycling versus running training on sprint and endurance capacity in inline speed skating. J Sports Sci Med 15:41–49
Stone MH, Sands WA, Pierce KC, Carlock J, Cardinale M, Newton RU (2005) Relationship of maximum strength to weightlifting performance. Med Sci Sports Exerc 37(6):1037–1043. https://doi.org/10.1249/01.mss.0000171621.45134.10
Tenan MS, Peng YL, Hackney AC, Griffin L (2013) Menstrual cycle mediates vastus medialis and vastus medialis oblique muscle activity. Med Sci Sports Exerc 45(11):2151–2157. https://doi.org/10.1249/MSS.0b013e318299a69d
Tillin NA, Pain MTG, Folland JP (2011) Short-term unilateral resistance training affects the agonist-antagonist but not the force-agonist activation relationship. Muscle Nerve 43(3):375–384. https://doi.org/10.1002/mus.21885
Tøien T, Malmo T, Espedal L, Wang E (2022) Maximal intended velocity enhances strength training-induced neuromuscular stimulation in older adults. Eur J Appl Physiol 122(12):2627–2636. https://doi.org/10.1007/s00421-022-05045-8
Torrejón A, Balsalobre-Fernández C, Haff GG, García-Ramos A (2019) The load-velocity profile differs more between men and women than between individuals with different strength levels. Sports Biomech 18(3):245–255. https://doi.org/10.1080/14763141.2018.1433872
Trybulski R, Gepfert M, Gawel D, Bichowska M, Fostiak K, Wojdala G, Trybek G, Krzysztofik M, Wilk M (2022) Impact of movement tempo on bar velocity and time under tension in resistance exercises with different external loads. Biol Sport 39(3):547–554. https://doi.org/10.5114/biolsport.2022.106160
Turner A (2011) The science and practice of periodization: A brief review. In Strength Conditioning J 33(1):34–46. https://doi.org/10.1519/SSC.0b013e3182079cdf
Turner HM, I., & Bernard, R. M. (2006) Calculating and synthesizing effect sizes. Contemporary Issues Commun Sci Disorders 33(Spring):42–55. https://doi.org/10.1044/cicsd_33_s_42
Weakley J, Mann B, Banyard H, Mclaren S, Scott T, Garcia-Ramos A (2020) Velocity-based training: from theory to application. Strength Conditioning J 43(2):31–49
Weakley J, Cowley N, Schoenfeld BJ, Read DB, Timmins RG, García-Ramos A, McGuckian TB (2023) The effect of feedback on resistance training performance and adaptations: a systematic review and meta-analysis. In Sports Med 53(9):1789–1803. https://doi.org/10.1007/s40279-023-01877-2
Weidauer L, Zwart MB, Clapper J, Albert J, Vukovich M, Specker B (2020) Neuromuscular performance changes throughout the menstrual cycle in physically active females. J Musculoskeletal Neuronal Interact 20(3):314–324
Wilk M, Zajac A, Tufano JJ (2021) The influence of movement tempo during resistance training on muscular strength and hypertrophy responses: a review. In Sports Med 51(8):1629–1650. https://doi.org/10.1007/s40279-021-01465-2
Włodarczyk M, Adamus P, Zieliński J, Kantanista A (2021) Effects of velocity-based training on strength and power in elite athletes—a systematic review. In Int J Environ Res Public Health 18(10):5257. https://doi.org/10.3390/ijerph18105257
Zinoubi B, Zbidi S, Vandewalle H, Chamari K, Driss T (2018) Relationships between rating of perceived exertion, heart rate and blood lactate during continuous and alternated-intensity cycling exercises. Biol Sport 35(1):29–37. https://doi.org/10.5114/biolsport.2018.70749
Funding
The University of Rome “Foro Italico” economically supported this work.
Author information
Authors and Affiliations
Contributions
EL conceived and designed the study, collected and analyzed the data, interpreted results, created figures and drafted the manuscript. RR conceived and designed the study, collected the data, interpreted results and drafted the manuscript. GF collected the data, interpreted results and drafted the manuscript. FF interpreted results and drafted the manuscript. MFP conceived and designed the study, interpreted results and drafted the manuscript. IB conceived and designed the study, interpreted results and drafted the manuscript. All authors participated in contributing to text and content of the manuscript, including revisions and edits. All authors approved the content of the manuscript and agree to be held accountable for the work.
Corresponding author
Ethics declarations
Conflict of interest
The authors declared no conflict of interest.
Additional information
Communicated by William J. Kraemer.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lecce, E., Romagnoli, R., Frinolli, G. et al. Exerting force at the maximal speed drives the increase in power output in elite athletes after 4 weeks of resistance training. Eur J Appl Physiol (2024). https://doi.org/10.1007/s00421-024-05604-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00421-024-05604-1