
Abstract
The purpose of the present study was to determine: (1) if different acute neuromuscular responses occur between constant versus variable external resistance machines, two commonly used resistance machines and (2) whether the potential differences in fatigability between young and older men influence the magnitude of acute response between these resistance machines. Twelve young men (28 ± 5 year) and 13 older men (65.4 ± 4 year) performed 15 × 1 repetition maximum and 5 × 10 repetitions isotonic knee extension resistance loadings with both constant and variable resistance (four loadings in total). Maximum isometric knee extension torque, superimposed twitch, resting twitch torque, maximal M wave properties, electromyograph, and blood lactate concentration measured the effects of loading. Concentric torque reduced to a greater extent during variable 15 × 1 versus constant loading in young men only (P < 0.05). While three out of the four loadings caused decreased voluntary activation in young men, only 15 × 1 using variable resistance caused reductions in older men (P < 0.05). 5 × 10 variable resistance loading significantly increased M wave duration and decreased EMG median frequency, which was not observed following constant resistance loading in both age groups. Acute decreases in force production were significantly greater in young men following all loading protocols (P < 0.05). Both young and older men showed indications of greater fatigue from variable resistance loadings. Differing muscle properties may have led to different magnitudes of fatigue between groups, and older subjects may benefit from specifically tailored training programs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Resistance training increases and/or maintains neuromuscular properties, such as strength and muscle mass, in both young and older adults (Moritani and deVries 1980; Häkkinen et al. 1998; Newton et al. 2002). Resistance machines that provide either constant or variable external resistance are widely available to recreational resistance trainers. Variable resistance machines modify the lever arm distance at certain phases of the range of motion to create greater resistance in accordance with human strength curves (Kulig et al. 1984). Despite the abundant use of these machines, the acute effects of a single resistance training session, hereafter referred to as loading, has not been fully explored, especially in older adults. Previous studies (Häkkinen et al. 1988; Walker et al. 2011) have indicated greater acute fatigue following variable resistance loading in young men, which may suggest a greater potential for long-term adaptation. However, performance measures and use of surface electromyography (EMG) alone as a measure of muscle activation do not provide detailed information regarding the possible causes of fatigue and how these two different machines affect the neuromuscular system.
An increasing amount of scientific attention has been directed to quantifying, and attempting to identify the causes of, age-related differences of loading-induced neuromuscular fatigue (Allman and Rice 2001; Callahan and Kent-Braun 2011; Hunter et al. 2008; Yoon et al. 2008). Despite some contrasting results, attributed to variations in study variables (e.g., contraction velocity, subject population, etc.), most studies have observed reduced fatigability in older subjects (for review see Allman and Rice 2002; Christie et al. 2011).
As a lower magnitude of fatigue in older adults would be expected, this may dampen the expected differences in acute neuromuscular fatigue between constant and variable resistance loadings. This phenomenon may negate the potential benefits of using variable resistance as part of a long-term training strategy for older adults. Therefore, the acute responses to loadings using constant versus variable external resistance in older, as well as young adults should be investigated. Additionally, as maximal strength and muscle size are important qualities in improving/maintaining functional capacity throughout the life span, the present study employed a protocol used in training for maximal strength development and a protocol used in training for muscle hypertrophy (Campos et al. 2002).
The purpose of the present study was to determine: (1) if different acute neuromuscular responses occur between constant versus variable external resistance machines, two commonly used resistance machines and (2) whether the potential differences in fatigability between young and older men influence the magnitude of acute response between these resistance machines. We hypothesized that the overall magnitude of fatigue would be lower in older men, and that, due to a lower magnitude of fatigue, there would be no differences between constant and variable resistance loadings in the older group.
Methods
Subjects
Twelve young men (28 ± 5 year, 180.8 ± 3.6 cm, 78.7 ± 9.8 kg) and 13 older men (65.4 ± 4 year, 175.6 ± 5 cm, 78.6 ± 8.9 kg) volunteered to participate in this study. The subjects were healthy and physically active that took part in low-intensity endurance-based physical activity 2–4 times per week (e.g., jogging, cycling, and cross-country skiing), although none performed systematic resistance training. Before testing, subjects were fully informed of the procedures and risks, and each subject provided written informed consent. Additionally, the older subjects were checked by a physician, including ECG monitoring, to ensure that no foreseeable health complications would arise due to maximal physical effort. The study was approved by the local ethics committee, and was conducted in accordance with the Declaration of Helsinki.
Study design
The subjects reported to the laboratory for a familiarization session during which anthropometric measures (height and weight) and vastus lateralis cross-sectional area, assessed by ultrasound (model SSD-α10, Aloka Co Ltd, Japan) at 50 % of femur length (see Ahtiainen et al. 2010 for protocol details), were determined. Also in this session, the test apparatus was set according to the subject’s individual anthropometry, surface electromyography (EMG) placements were measured and marked by indelible ink tattoos, submaximal muscle and nerve stimulation was performed to familiarize the subjects with stimulation procedures, and the subjects were taught and practiced maximum isometric knee extension and flexion trials. Furthermore, the subjects performed a one repetition maximum test (1RM) using the variable resistance setting to determine the loads used during subsequent loading sessions (a conversion factor based on our pilot testing was used to estimate constant resistance 1RM loads). Subjects performed sets of progressively increasing load (1 × 10 × 70 % estimated 1RM, 1 × 7 × 75 % estimated 1RM, 1 × 5 × 80 % estimated 1RM, 1 × 1 × 90 % estimated 1RM) in order to fully prepare for maximal contractions. Thereafter, single repetitions using 5 kg increments were performed until the subject could no longer lift the load from a knee angle of approx. 60° to full extension (180° knee extension). Three to four repetitions were needed to determine each subject’s 1RM.
Approximately 1 week (7 ± 1 day) after familiarization the subjects returned for the first isotonic loading session. A total of four loadings were completed in a randomized order with 7 (±1) days between loadings. The loadings were: (1) 15 sets of 1 repetition maximum (15 × 1RM) with 3 min rest between sets using constant external resistance, (2) 15 × 1RM using variable resistance, (3) 5 sets of 10 repetitions (5 × 10RM) with an initial load of 80 % 1RM with 2 min rest between sets using constant external resistance and (4) 5 × 10RM using variable resistance.
The subjects refrained from exercise for 48 h before each test session. Each subject had their own test start time during the day, which was maintained throughout testing to control for diurnal variations. Young and older subjects arrived at the laboratory in alternating fashion to ensure no between-group time differences occurred.
Loading protocol
Subjects were instructed to consume 0.5 L water 60 min before they arrived, as hydration status has been shown to affect strength (Judelson et al. 2007). All actions were performed on a David 200 knee extension device (David Health Solutions Ltd, Finland), which was modified with a locking system and strain gauges to allow evaluation of isometric performance. Subjects were secured by a non-elastic strap at the hip and a pad across the knee to prevent extraneous movement. The device was also modified so that it could either produce constant external resistance (i.e., the same resistance throughout the range of motion) or variable resistance through a cam. The cam creates a bell-shaped torque curve and the mean relative increase in external resistance at 100°–140° knee angles in the present study was 24 % in young and 21 % in older men compared to constant resistance (see Häkkinen et al. 1987 for a more detailed quantification of the torque–angle curve). Average concentric torque throughout the range of motion (i.e., 60°–180° knee angle) was measured during the isotonic actions of each loading session by strain gauges and an electric goniometer fitted to the device. Both torque and angle data were sampled at 2000 Hz and filtered by a 20 Hz low-pass filter.
The experimental session began with a warm up of 5 bilateral knee extension repetitions using 50 %1RM followed by two unilateral maximal isometric knee extension and knee flexion contractions (all unilateral actions were performed by the right leg). The knee and hip angles during isometric contractions were 107° and 110°, respectively. Thereafter, the maximal torque response to muscle stimulation of the right quadriceps was determined. Once there was an observed torque plateau, a further three stimulations were delivered during resting conditions. Subjects then performed three maximal unilateral isometric knee extension contractions of 3–5 s duration with superimposed twitch (Merton 1954) evoked at the peak force to assess voluntary activation. Verbal encouragement and visual feedback were provided during all contractions.
Maximum M wave properties of the vastus lateralis and vastus medialis in a standing position were then examined by femoral nerve stimulation. A standing position was chosen to determine M wave properties because, during pilot testing, our measurements in a seated position did not achieve an acceptable level of reliability (possibly due to changes in cathode pressure and position relative to the nerve). Finally, a fingertip blood lactate sample was taken to complete the pre-loading tests.
Subjects then completed one out of the four assigned loading protocols. If the subject completed the desired repetition(s), the load increased slightly to ensure sets of true repetition maximums. If the subjects could no longer complete the desired number of repetitions, they were assisted during the concentric phase by an experienced trainer so that required repetitions could be completed and the resistance was lowered for the following set. Once all sets and repetitions had been completed, post-loading tests were performed immediately and in the same order as pre-loading; namely three resting twitches, three unilateral maximum isometric knee extension with superimposed twitch, standing femoral nerve stimulation, and a blood lactate sample (5 min after loading cessation).
Stimulation procedures
Muscle stimulation was performed by placing four, galvanically paired, self-adhesive electrodes (6.98 cm V-trodes, Mettler Electronics Corp, USA) on the proximal and mid regions of the quadriceps muscle belly. Single 1 ms rectangular pulses were delivered by a constant-current stimulator (Model DS7AH, Digitimer Ltd, UK) until a torque plateau was observed. An additional 25 % of stimulation current was added to the current identified to produce maximum torque. During the unilateral maximum isometric knee extension trials, the same single-pulse stimulation was delivered during the plateau of peak torque and then one more pulse 2 s after contraction cessation to assess voluntary activation. Biceps femoris (BF) EMG showed that our methods did not stimulate antagonist muscles.
M wave properties were assessed by femoral nerve stimulation. Subjects were instructed to stand fully upright with their bodyweight balanced equally between both legs, feet hip width apart. The stimulating cathode (1 cm diameter) was placed firmly into the femoral triangle at the point that gave the strongest response to a weak stimulation current, which was marked on the skin for replacement. The anode (6.98 cm V-trode) was placed on the greater trochanter. Current was increased in 10 mA stages (1 ms single-pulse, 400 V) until there were clear plateaus in the M wave amplitude of both vastus lateralis and vastus medialis. Thereafter, an additional 25 % of stimulation current was applied.
Surface EMG
Bipolar Ag/AgCl electrodes (10 mm diameter, 20 mm inter-electrode distance, common mode rejection ratio >100 dB, input impedance >100 MΩ, baseline noise <1 μV rms) (Department of Biology of Physical Activity, University of Jyväskylä, Finland) were positioned, following shaving and skin abrasion, on the vastus lateralis (VL), vastus medialis (VM), and BF of the right leg according to SENIAM guidelines (Hermens et al. 1999). During the maximum isometric trials with superimposed twitch, surface EMG was sampled at a frequency of 2000 Hz and amplified at a gain of 500 (sampling bandwidth 10–500 Hz). Signals were passed in real time to an AD converter (Micro1401, Cambridge Electronic Design, UK) and recorded by Signal 4.04 software (Cambridge Electronic Design, UK). After testing, EMG signals were band-pass filtered (20–350 Hz) and converted to root mean square (rms) for EMG amplitude or analyzed by fast Fourier transformation (Hamming, 1024 data points) for EMG median frequency over a 500 ms epoche immediately before the superimposed twitch.
Data analysis
Maximal unilateral isometric torque (sampled at 2000 Hz, and filtered by a 20 Hz low-pass filter) was considered as the greatest torque prior to the superimposed twitch. Voluntary activation was assessed from the additional torque produced by the superimposed twitch and the maximum torque of the subsequent resting twitch using the formula of Bigland-Ritchie et al. (1983); activation % = [1−(Pts/Pt)] × 100. Resting twitches were analyzed for maximum torque, maximum rate of twitch torque production (10 ms epoche) and half-relaxation time. Maximum M wave properties were analyzed for peak-to-peak amplitude and peak-to-peak duration. Fingertip blood samples were collected into 20 μL capillary tube and were analyzed following the manufacturers’ instructions (EKF diagnostic, Biosen, Germany).
Statistical analysis
Conventional statistical methods were used to obtain mean values, standard deviations, and correlation coefficients. Normal distribution was determined through the Shapiro–Wilk test. Baseline neuromuscular properties were assessed by independent t test. Repeated measures ANCOVA (2 group × 4 loading × 2 time) was used to determine significant main effects in torque variables (baseline values were applied as the covariate). Repeated measures ANOVA (2 group × 4 loading × 2 time) was used to determine significant main effects in median frequency and M wave variables. Bonferroni adjustments were applied as post hoc tests for both ANCOVA and ANOVA assessments. EMG amplitude was analyzed by paired t test (within group; pre- vs. post-loading). Differences in relative changes pre- to post-loading were assessed by one-way ANOVA. The alpha level was set at 0.05.
Inter-session intra-class correlation coefficients and CV %, assessed from the pre-loading values obtained for each loading session, were at acceptable levels; isometric torque = 0.981 and 3.4 %, EMG amplitude = 0.918 and 7.2 %, median frequency = 0.957 and 6.8 %, maximum twitch torque = 0.994 and 1.3 %, maximum rate of twitch torque development = 0.997 and 3.2 %, half-relaxation time = 0.96 and 7.6 %, M wave amplitude = 0.92 and 7.4 %, M wave duration = 0.77 and 11 %, and calculated voluntary activation = 0.732 and 1.9 %.
Results
At baseline (pre-loading), the young subjects were stronger, had greater VL cross-sectional area, and demonstrated greater twitch torque (both maximum and rate of torque development) compared to the older subjects (P < 0.05; Table 1). Both VL cross-sectional area (r = 0.42, P = 0.04, n = 24) and rate of twitch torque development (r = 0.69, P = 0.01, n = 24) were significantly related to maximum unilateral isometric torque when the groups were combined. When the data were normalized based on VL cross-sectional area, the differences observed in voluntary (young = 9.2 ± 2.5 Nm cm−2 vs. older = 8.6 ± 1.3 Nm cm−2) and electrically evoked torque (e.g., maximum twitch torque; young = 3.1 ± 0.7 Nm cm−2 vs. older = 2.9 ± 0.5 Nm cm−2) were no longer significant. There were no differences in voluntary activation level between young and older men (Table 1).
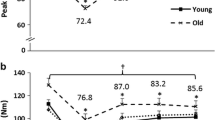
Significant main effects for time (P < 0.001) and age group (P < 0.01) were found for concentric torque. Table 2 shows that the relative change in concentric torque between variable versus constant loading in young men was statistically significant during 15 × 1RM (P < 0.05; Table 2). Also, young men produced significantly greater concentric torque compared to older men during 15 × 1RM (P < 0.05; Table 2). Significant main effects for time (P < 0.001), age group (P < 0.001) and interactions (time × age P < 0.001, time × loading P < 0.001) for maximum isometric torque were observed. Post hoc analysis revealed significant decreases in maximum isometric torque were observed after all four loadings (P < 0.05). Furthermore, there was a trend that variable resistance loading caused a greater decrease in isometric torque following 5 × 10RM (−44 ± 11 % vs. −50 ± 15 %, P = 0.058, Fig. 1a) in young men. Age influenced the magnitude of loading-induced fatigue in that the older men maintained a higher percentage of their pre-loading performance compared to the young men during all loadings (Fig. 1a).

Relative changes pre- to post-loading in unilateral maximum isometric torque (a) and post-loading blood lactate concentrations (b) in young and older men (mean ± SD). Symbols inside the bars depict significant within-group difference from pre-loading, *P < 0.05, **P < 0.01. O depicts significant difference versus older men, O = P < 0.05. †Trend between constant and variable resistance loading, P = 0.058
Significant main effects for blood lactate were observed for time (P < 0.001), age group (P < 0.001) and interactions (time × age P < 0.001, time × loading P < 0.001). Blood lactate was increased in both groups only after both 5 × 10RM protocols. However, young men had higher blood lactate concentrations than older men post-loading (Fig. 1b; P < 0.01), although there were no differences when comparing variable versus constant resistance loadings.
Post-loading EMG amplitude was reduced following the 15 × 1RM protocols only (P < 0.05), especially in young men (Fig. 2). In older men, VM activity was reduced (−20 ± 24 %, P = 0.018, Fig. 2) following 15 × 1RM using variable resistance. In young men, significant reductions in EMG amplitude were observed in both VL and VM muscles following variable resistance loading (VL = −18 ± 20 %, P = 0.041; VM = −30 ± 22 %, P = 0.005) and constant resistance loading (VL = −29 ± 26 %, P = 0.012; VM = −22 ± 24 %, P = 0.012) (Fig. 2).

Relative changes pre- to post-loading in isometric EMG amplitude of the vastus lateralis and medialis muscles in the older (a) and young (b) men (mean ± SD). Symbols inside the bars depict significant within-group difference from pre-loading, *P < 0.05, **P < 0.01
A significant main effect for time (P < 0.001) was observed for voluntary activation level. Both 15 × 1RM loading protocols caused decreased voluntary activation levels in young men (constant and variable resistance, Fig. 3a, b, respectively). However, only the 15 × 1RM loading with variable resistance caused changes in older men (Fig. 3b). Following 5 × 10RM loading, only young men showed significant decreases in voluntary activation level when using variable resistance (Fig. 3d; P < 0.05).

Voluntary activation level (mean ± SD) in young and older men during 15 × 1RM using constant resistance (a), 15 × 1RM using variable resistance (b), 5 × 10RM using constant resistance (c), and 5 × 10RM using variable resistance (d). Symbols inside the bars depict significant within-group difference from pre-loading, *P < 0.05, **P < 0.01
When muscle stimulation was delivered on the resting quadriceps, significant main effects for time (P < 0.001), age group (P = 0.012), and interactions (time × age P < 0.001, time × loading P < 0.001, time × age × loading P = 0.03,) were observed in maximum twitch torque. The maximum twitch torque was reduced in both young and older men following 15 × 1RM using variable resistance loading and following both 5 × 10RM protocols (Fig. 4). Comparing young and older men, there were greater reductions (P < 0.05) in maximum twitch torque following 5 × 10RM using variable resistance in the young men (Fig. 4). Results were similar for maximum rate of twitch torque development but no changes were observed for half-relaxation time.

Relative changes pre- to post-loading in maximum twitch torque (mean ± SD). Symbols inside the bars depict significant within-group difference from pre-loading, *P < 0.05, **P < 0.01
Significant main effects for time (VL P = 0.008, VM P < 0.001), age group (VL P = 0.011, VM P = 0.03) and time × loading interaction (VL P = 0.01, VM P = 0.002) were found in peak-to-peak M wave duration. A significant increase was observed in VL M wave duration in older men following 5 × 10RM using variable resistance only (Fig. 5a). In young men, both the VL and the VM muscles showed increased M wave duration following 5 × 10RM using variable resistance, while the VL muscle had increased M wave duration following 5 × 10RM using constant resistance (Fig. 5b). There were no loading-induced changes in M wave amplitude for the VL and VM muscles.

Peak-to-peak M wave duration (mean ± SD) during 5 × 10RM loadings in older (a) and young (b) men. Symbols inside the bars depict significant within-group difference from pre-loading. Values within the bars depict the mean relative change from pre- to post-loading, *P < 0.05
During maximum unilateral isometric knee extension, EMG median frequency showed a significant main effect for time loading interaction (P < 0.001). Post hoc revealed that median frequency was reduced following variable resistance 5 × 10RM loading in the older (VL, −14.1 ± 10.4 %, P = 0.004) and young (VM, −11.8 ± 12.7 %, P = 0.019) men. Also, this significant difference was observed in both groups when the VL and VM muscles were combined and averaged (P < 0.05). No loading-induced changes were observed in either group during any other loading.
Discussion
The magnitudes of neuromuscular fatigue induced by two typical, but very different, resistance loading protocols were greater in young men compared to older men. In agreement with our hypothesis, only young men had a greater acute reduction in force production following variable versus constant resistance loadings. However, the use of variable resistance loading caused more pronounced decreases in voluntary activation level, M wave duration, and EMG variables in both young and older men compared to constant external resistance. The differences between the responses of young and older men may be largely central, as observed following both loadings, but also peripheral in origin following 5 × 10 loading in the present study.
At baseline, the young men demonstrated greater isometric torque, larger cross-sectional area of the vastus lateralis, and greater resting twitch torque and rate of twitch torque development. There was no difference, however, in the voluntary activation level between groups. The observed group differences are likely to be due to muscular properties as there were no between-group differences when torque was normalized to VL CSA. These findings suggest that healthy (physically active) aging may maintain the ability to activate muscles, but that changes within the muscle itself likely account for differences in performance. Furthermore, the larger muscle mass and possibly more fast-twitch muscle properties (not examined in the present study) in young men may help to explain some of the observed differences in loading-induced fatigue, blood lactate concentration, resting twitch torque, and M wave duration between the young and older men during 5 × 10RM loadings.
Although we found no significant differences between the two age groups, it must be acknowledged that some studies have shown an age-related decline in voluntary activation (e.g., Stackhouse et al. 2001; Yoon et al. 2008) and that the stimulation methodology of the present study has been suggested to lack sensitivity (Herbert and Gandevia 1999) in distinguishing voluntary activation level between groups. However, it should be pointed out that the mean age of the older group was “only” 65 years, they were physically active and that similar findings to the present study have been previously shown in subjects of 65–86 years (Allman and Rice 2001, 2003; Callahan et al. 2009; Dalton et al. 2012).
The loading-induced reductions in isometric torque are in agreement with what would be expected during these isotonic loading protocols in young men (Walker et al. 2012). In women (Häkkinen 1995), there was a significant difference in loading-induced fatigue between young and older subjects (P < 0.01) following the same 5 × 10RM leg press loading as the present study. However, in another study (Häkkinen and Pakarinen 1995), there were no significant differences in isometric leg extension force reductions post-loading between young (mean change −24 %) and older subjects (mean change −20 %). Perhaps, the equivalent results in the study of Häkkinen & Pakarinen (1995) were due to a lower than expected reduction in maximal isometric force (−24 % in Häkkinen & Pakarinen vs. −44 % in the present study) minimizing the between-group differences.
In the present study, it was possible to quantify potential mechanisms of fatigue when comparing constant and variable resistance loadings in young and old men. Firstly, considering the 15 × 1RM protocol with constant external resistance, young men showed decreases in maximum isometric torque that were accompanied by decreased voluntary activation and reduced EMG amplitude in both VL and VM muscles, suggesting that there was a decrease in the ability to activate the muscles (i.e., central fatigue). Conversely, older men showed no change in voluntary activation or EMG amplitude. When variable resistance was used, the older men demonstrated similar neuromuscular changes as young men, specifically, significant reductions in voluntary activation and VM EMG amplitude. Both young and older men demonstrated reduced resting twitch torque following the 15 × 1RM protocol with variable external resistance only. This suggests that peripheral fatigue also occurred, which was independent of changes in blood lactate concentration, possibly as a consequence of higher resistance during the mid-range of motion (Häkkinen et al. 1987).
Following 5 × 10RM loadings, it appeared that variable resistance loading influenced central and peripheral parts of the neuromuscular system to a greater extent in both age groups compared to constant resistance loading. In the young, significant reductions in voluntary activation and EMG median frequency were observed, as well as greater M wave duration in both VL and VM muscles. The combined reduction in voluntary activation and median frequency data may indicate that variable resistance loading impaired the activation of Type II muscle fibers more than following constant resistance, which may have been expected when comparing young and older subjects (Yamada et al. 2000). However, during 5 × 10RM variable resistance loading significant changes in M wave duration and EMG median frequency also occurred in the older men, which were not observed following constant resistance loading. As there were no differences in blood lactate concentration or changes in fluid volume (unpublished observation) between constant and variable resistance loadings, one possible explanation for the prolonged M wave duration in both young and older men could be greater damage to the sarcolemma influencing Na+/K+ concentrations (Juel 1988; McBride et al. 2000). This may be due to the higher resistances, as muscle strain has been shown to influence the magnitude of muscle damage (Lieber and Friden 1993).
It is noteworthy that age differences were observed in peripheral fatigue (resting twitch torque data), as well as central fatigue (reduced maximum voluntary activation level), during the 5 × 10RM loadings. It is possible that greater impairments in conduction velocity may have occurred in the young subjects, which would accompany the observed larger blood lactate concentrations (Juel 1988; Brody et al. 1991), as indicated by significant increases in M wave duration during both constant and variable resistance loadings. However, M wave duration was significantly prolonged in the VL of older men also during 5 × 10RM variable resistance loading, suggesting that impairments to sarcolemma function may have been similar between the groups. Another explanation is that greater anaerobic metabolism (Lanza et al. 2005) and depletion of ATP-PC stores (Gorostiaga et al. 2010) may have occurred in young men to cause greater losses in twitch torque, as a consequence of greater muscle mass and/or possibly greater proportion of fast-twitch fibers. This would be associated with higher anaerobic metabolism (Lanza et al. 2005) and the observed higher blood lactate concentrations.
The more readily observed decreases in voluntary activation level and EMG amplitude in young men may suggest that older men were more resistant to central fatigue during the present resistance loadings. Possible mechanisms for this are unclear. Nevertheless, these findings may suggest that the elderly may benefit from different resistance training protocols than are usually prescribed for young subjects. For example, a higher training volume has recently been suggested to be beneficial in the elderly (Peterson et al. 2011).
Most previous studies have observed greater acute neuromuscular fatigue in young versus older adults (e.g., Häkkinen 1995; Lanza et al. 2004; Baudry et al. 2007). However, of the studies showing the opposite trend, one common factor seems to be the use of high velocity dynamic contractions (Petrella et al. 2005; McNeil and Rice 2007; Callahan and Kent-Braun 2011; Dalton et al. 2012). The present study used relatively slow contractions speeds (approx. 2–5 s concentric phase), as is typical of these types of loading, which may help to explain different findings between studies. Collectively, these may be very important findings, as the need to maintain power in old age is highlighted by the greater loss in explosive force capacity compared with maximum force capacity (Häkkinen et al. 1998).
Conclusions
Greater indications of fatigue were observed when using variable resistance compared to constant resistance in both groups, but especially in the young men. Variable resistance may induce greater adaptations in young, and perhaps also older, men if used during long-term training. The different magnitude of peripheral fatigue following 5 × 10RM loading may have been influenced by different muscle properties of the groups. However, it seems that older men are more resistant to central fatigue during typical maximal strength or hypertrophic loading protocols than young men. Based on the results, it is suggested that loading protocols be specifically tailored for different age groups, and evaluated through training studies, in order to optimize training adaptations.
References
Ahtiainen JP, Hoffren M, Hulmi JJ, Pietikäinen M, Mero AA, Avela J, Häkkinen K (2010) Panoramic ultrasonography is a valid method to measure changes in skeletal muscle cross-sectional area. Eur J Appl Physiol 108(2):273–279
Allman BL, Rice CL (2001) Incomplete recovery of voluntary isometric force after fatigue is not affected by old age. Muscle Nerve 24(9):1156–1167
Allman BL, Rice CL (2002) Neuromuscular fatigue and aging: central and peripheral factors. Muscle Nerve 25(6):785–796
Allman BL, Rice CL (2003) Perceived exertion is elevated in old age during an isometric fatigue task. Eur J Appl Physiol 89(2):191–197
Baudry S, Klass M, Pasquet B, Duchateau J (2007) Age-related fatigability of the ankle dorsiflexor muscles during concentric and eccentric contractions. Eur J Appl Physiol 100(5):515–525
Bigland-Ritchie B, Johansson R, Lippold OC, Woods JJ (1983) Contractile speed and EMG changes during fatigue of sustained maximal voluntary contractions. J Neurophysiol 50(1):313–324
Brody LR, Pollock MT, Roy SH, De Luca CJ, Celli B (1991) pH-induced effects on median frequency and conduction velocity of the myoelectric signal. J Appl Physiol 71(5):1878–1885
Callahan DM, Kent-Braun JA (2011) Effect of old age on human skeletal muscle force-velocity and fatigue properties. J Appl Physiol 111(5):1345–1352
Callahan DM, Foulis SA, Kent-Braun JA (2009) Age-related fatigue resistance in the knee extensor muscles is specific to contraction mode. Muscle Nerve 39(5):692–702
Campos GE, Luecke TJ, Wendeln HK et al (2002) Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. Eur J Appl Physiol 88(1–2):50–60
Christie A, Snook EM, Kent-Braun JA (2011) Systematic review and meta-analysis of skeletal muscle fatigue in old age. Med Sci Sports Exerc 43(4):568–577
Dalton BH, Power GA, Vandervoort AA, Rice CL (2012) The age-related slowing of voluntary shortening velocity exacerbates power loss during repeated fast knee extensions. Exp Gerontol 47(1):85–92
Gorostiaga EM, Navarro-Amezqueta I, Cusso R et al (2010) Anaerobic energy expenditure and mechanical efficiency during exhaustive leg press exercise. PlosOne 5(10):e13486
Häkkinen K (1995) Neuromuscular fatigue and recovery in women at different ages during heavy resistance loading. Electromyogr Clin Neurophysiol 35(7):403–413
Häkkinen K, Pakarinen A (1995) Acute hormonal responses to heavy resistance exercise in men and women at different ages. Int J Sports Med 16(8):507–513
Häkkinen K, Komi PV, Kauhanen H (1987) Scientific evaluation of specific loading of the knee extensors with variable resistance, “isokinetic” and barbell exercises. Med Sport Sci 26:224–237
Häkkinen K, Kauhanen H, Komi PV (1988) Effects of fatiguing loading with variable resistance equipment on neural activation and force production of the knee extensor muscles. Electromyogr Clin Neurophysiol 28:79–87
Häkkinen K, Newton RU, Gordon SE et al (1998) Changes in muscle morphology, electromyographic activity, and force production characteristics during progressive strength training in young and older men. J Gerontol A Biol Sci Med Sci 53(6):B415–B423
Herbert RD, Gandevia SC (1999) Twitch interpolation in human muscles: mechanisms and implications for measurement of voluntary activation. J Neurophysiol 82(5):2271–2283
Hermens HJ, Freriks B, Merletti R et al (1999) European recommendations for surface electromyography, results of the SENIAM project 1999. Roessingh Research and Development, Enschede, pp 44–46
Hunter SK, Todd G, Butler JE, Gandevia SC, Taylor JL (2008) Recovery from supraspinal fatigue is slowed in old adults after fatiguing maximal isometric contractions. J Appl Physiol 105(4):1199–1209
Judelson DA, Maresh CM, Farrell MJ et al (2007) Effect of hydration state on strength, power, and resistance exercise performance. Med Sci Sports Exerc 39(10):1817–1824
Juel C (1988) Muscle action potential propagation velocity changes during activity. Muscle Nerve 11(7):714–719
Kulig K, Andrews JG, Hay JG (1984) Human strength curves. Exerc Sport Sci Rev 12:417–466
Lanza IR, Russ DW, Kent-Braun JA (2004) Age-related enhancement of fatigue resistance is evident in men during both isometric and dynamic tasks. J Appl Physiol 97(3):967–975
Lanza IR, Befroy DE, Kent-Braun JA (2005) Age-related changes in ATP-producing pathways in human skeletal muscle in vivo. J Appl Physiol 99(5):1736–1744
Lieber RL, Friden J (1993) Muscle damage is not a function of muscle force but active muscle strain. J Appl Physiol 74(2):520–526
McBride TA, Stockert BW, Gorin FA, Carlsen RC (2000) Stretch-activated ion channels contribute to membrane depolarization after eccentric contractions. J Appl Physiol 88(1):91–101
McNeil CJ, Rice CL (2007) Fatigability is increased with age during velocity-dependent contractions of the dorsiflexors. J Gerontol A Biol Sci Med Sci 62(6):624–629
Merton PA (1954) Voluntary strength and fatigue. J Physiol 123(3):553–564
Moritani T, deVries HA (1980) Potential for gross muscle hypertrophy in older men. J Gerontol. 35(5):672–682
Newton RU, Häkkinen K, Häkkinen A, McCormick M, Volek J, Kraemer WJ (2002) Mixed-methods resistance training increases power and strength of young and old men. Med Sci Sports Exerc 34(8):1367–1375
Peterson MD, Sen A, Gordon PM (2011) Influence of resistance exercise on lean body mass in aging adults: a meta-analysis. Med Sci Sports Exerc 43(2):249–258
Petrella JK, Kim J-S, Tuggle SC, Hall SR, Bamman MM (2005) Age differences in knee extension power, contractile velocity, and fatigability. J Appl Physiol 98(1):211–220
Stackhouse SK, Stevens JE, Lee SC, Pearce KM, Snyder-Mackler L, Binder-Macleod SA (2001) Maximum voluntary activation in nonfatigued and fatigued muscle of young and elderly individuals. Phys Ther 81(5):1102–1109
Walker S, Taipale RS, Nyman K, Kraemer WJ, Häkkinen K (2011) Neuromuscular and hormonal responses to constant and variable resistance loadings. Med Sci Sports Exerc 43(1):26–33
Walker S, Davis L, Avela J, Häkkinen K (2012) Neuromuscular fatigue during dynamic maximal strength and hypertrophic resistance loadings. J Electromyogr Kinesiol 22(3):356–362
Yamada H, Okada M, Oda T, Nemoto S, Shiozaki T, Kizuka T, Kuno S, Masuda T (2000) Effects of aging on EMG variables during fatiguing isometric contractions. J Hum Ergol 29(1–2):7–14
Yoon T, Schlinder De-Lap B, Griffith EE, Hunter SK (2008) Age-related muscle fatigue after a low-force fatiguing contraction is explained by central fatigue. Muscle Nerve 37(4):457–466
Acknowledgments
The authors wish to thank the subjects for their efforts during this study. We also acknowledge the contribution of Pirkko Puttonen during data analysis. This work was funded in part by the Department of Biology of Physical Activity, University of Jyväskylä and by personal grants to Simon Walker from The National Doctoral Program of Musculoskeletal Disorders and Biomaterials, Finland and the Rector of the University of Jyväskylä, Finland.
Conflict of interest
All authors declare no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Toshio Moritani.
Rights and permissions
About this article
Cite this article
Walker, S., Peltonen, H., Avela, J. et al. Neuromuscular fatigue in young and older men using constant or variable resistance. Eur J Appl Physiol 113, 1069–1079 (2013). https://doi.org/10.1007/s00421-012-2526-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-012-2526-2