
Abstract
The influence of different work-to-rest (W:R) ratios during fatigue induced by maximal eccentric contractions is unknown. The present study sought to expand the understanding of the task-dependent nature of eccentric contractions, and the associated fatigue, during exercise and acute as well as extended recovery periods. Using a Biodex multi-joint dynamometer, the ankle dorsiflexors of eight men [26 (4) years] were fatigued with 150 maximal eccentric contractions. Set structure was manipulated such that one leg performed 3 sets of 50 repetitions (short rest protocol, SRP), and the other leg performed 15 sets of 10 repetitions (long rest protocol, LRP). A 1-min rest interval separated each set, which resulted in 2 and 14 min of total rest for the SRP and the LRP, respectively. At fatigue, the SRP demonstrated a marginally greater loss of average peak eccentric torque than the LRP (P<0.05). In the acute period following fatigue, isometric peak torque loss and the degree of low-frequency fatigue (LFF) were not recovered (P<0.05) and were equivalent for both protocols. Significant impairment of both eccentric and isometric torque was persistent and equal for each protocol at 96 h of recovery (P<0.05). These findings suggest that the W:R ratio has a modest influence on the fatigue (torque loss) induced by maximal eccentric contractions, but maximal isometric torque during recovery and LFF are insensitive to changes in total rest time.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Eccentric contractions are unique because large muscle forces are generated at relatively low metabolic cost (Newham et al. 1983), and, unlike isometric and concentric contractions, repetitive eccentric contractions induce substantial muscle damage (Clarkson and Hubal 2002; Lieber and Friden 2002; Warren et al. 2002). Similar to the other contraction types, performing consecutive eccentric contractions results in a loss of torque over time. However, it remains unclear whether the loss of torque associated with performing eccentric contractions is due entirely to persistent structural impairment (muscle damage), or whether the transient metabolic factors of muscle fatigue (conventional fatigue) commonly associated with isometric and concentric fatigue protocols also mediate the torque loss. Conventional fatigue mechanisms have been assumed to have little or no influence during eccentric fatigue protocols (Warren et al. 1993; Teague and Schwane 1995) but this has not been evaluated thoroughly.
The collective results of isometric and concentric fatigue protocols suggest that the performance impairment and the associated mechanisms are largely influenced by the task performed (Enoka and Stuart 1992; Bigland-Ritchie et al. 1995). In particular, the intensity (% of maximal effort) and number of contractions, as well as the rest periods provided between the fatiguing contractions (work-to-rest ratio, W:R), appear to be key factors in the task-dependent nature of muscle fatigue. Despite the limited number of studies on eccentric muscle fatigue, it seems clear that eccentric torque loss is exacerbated by increasing the contraction intensity or the number of contractions (Clarkson and Tremblay 1988; Warren et al. 1993; Brown et al. 1997; Nosaka et al. 2001). However, the extent to which eccentric torque loss at fatigue is associated with the W:R ratio is not well understood. Manipulation of the W:R ratio of eccentric protocols is an effective way to investigate the interaction between conventional fatigue and the structural impairment associated with the loss of torque following repetitive eccentric contractions.
The most widely proposed mechanism of impairment following eccentric contractions is a mechanical disruption of the link between the t-tubule and the sarcoplasmic reticulum (SR), causing dysfunction of the excitation-contraction (E-C) coupling mechanism (Lannergren et al. 1996; Ingalls et al. 1998; Warren et al. 2002; Westerblad and Allen 2002). This suggestion is supported by the persistent and preferential impairment at low compared to high frequencies of stimulation (low-frequency fatigue, LFF) that follows eccentric fatigue protocols (Jones 1981, 1996). Although a greater degree of LFF has been shown following eccentric versus concentric contractions (Davies and White 1981; Edwards et al. 1981; Newham et al. 1983), it is unknown whether an equivalent number of maximal eccentric contractions will induce the same degree of LFF despite large differences in the W:R ratios of the eccentric fatigue protocols.
A protocol with very limited rest during exercise (high W:R ratio) likely would, through the influence of conventional fatigue, cause a greater decline in torque than a protocol with a lower W:R ratio because, in the latter protocol, conventional fatigue would be effectively removed as an influence on torque loss. Consequently, conventional fatigue would cause the total torque production to be less in the high W:R ratio protocol resulting in less muscle damage. Because muscle damage would be minimized, torque loss in the later stages of acute recovery and in the subsequent days would be greater in the low versus the high W:R ratio protocol. This difference in muscle damage would also be reflected in the degree of LFF, in which the low W:R ratio protocol would have substantially greater LFF during the acute recovery period.
One study has examined the effect of the W:R ratio on repetitive eccentric contractions, and, although the role in eccentric fatigue was not quantified, it was suggested that the associated symptoms of muscle damage were influenced marginally by varying the W:R ratio (Teague and Schwane 1995). However, the exercise protocol consisted of a small number (10) of eccentric contractions performed at a low intensity (<50% eccentric maximal strength) and caused a limited loss of torque. Therefore, it remains to be determined whether manipulation of the W:R ratio is a principal factor in the task-dependency rubric of eccentric muscle fatigue when a large number of repetitive contractions are performed at maximal intensity to induce substantial fatigue.
Thus, the purpose of this study was to investigate whether eccentric torque loss during a high volume of repetitive maximal eccentric contractions is sensitive to considerable variation in the W:R ratio of the fatigue task. In addition, we examined the effect of total rest time on the isometric torque loss and LFF present in the acute recovery period. Based upon the assumption that the role of conventional fatigue is limited in eccentric contractions, we hypothesized that, despite large differences in the total rest time, two eccentric protocols with the same number of contractions would have equivalent torque loss during exercise and recovery. Moreover, the LFF that accompanies eccentric contractions would also be similar between the two protocols.
We studied the ankle dorsiflexors because recent studies have demonstrated that this muscle group is susceptible to eccentric torque loss during fatiguing contractions (Pasquet et al. 2000; Warren et al. 2000), with an appreciable degree of LFF persistent in the recovery period (Birtles et al. 2003). Furthermore, the dorsiflexors require a high volume of repeated eccentric contractions to induce substantial torque loss, thus this muscle group provides a model to create two protocols with very different W:R ratios in order to separate the influence of conventional fatigue from muscle damage during the eccentric exercise.
Methods
Subjects
Eight healthy men from the university environment volunteered for this study. Subjects were moderately active but not involved in any extreme lower body physical activities. The mean (±SD) age, height, and weight of the subjects were 26.3 (3.5) years, 177.2 (10.0) cm, and 73.9 (10.9) kg, respectively. The study was conducted in accordance with the guidelines for experimentation on human subjects established by the local university’s ethics review board, and informed written consent was obtained from each of the eight subjects. Subjects were tested on three separate sessions. The first session consisted of baseline measurements followed by the fatiguing protocols and an acute recovery of 30 min. Sessions two and three involved only the baseline measures and took place 48 and 96 h after the initial session. In session one, the leg to be tested first and the protocol that the leg would perform were selected randomly. Once determined, this testing order was maintained in the latter two sessions. The following procedures were performed on one leg and, after a short rest, were conducted again on the subject’s other leg.
Experimental set-up
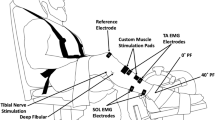
All sessions were performed on a Biodex multi-joint dynamometer (System 3, Biodex Medical Systems, Shirley, New York). Subjects sat in a reclined position with angles of approximately 90° and 120° at the hip and knee respectively, and their leg was aligned parallel to the ground (Fig. 1). This orientation provided access to the peroneal nerve for electrical stimulation. A Velcro strap across the thigh, an adjustable seat belt across the waist, and shoulder straps minimized hip flexion and upper body movement. Velcro straps across the toes and the top of the foot secured the subject to the dynamometer footplate (Fig. 1). During isometric contractions the ankle was positioned at 30° of plantar flexion.

Schematic of the experimental set up. Dynamometer settings for each subject were recorded during session one and then repeated for sessions two and three
A computer-triggered stimulator (model DS7A, Digitimer, Welwyn Garden City, Hertfordshire, UK) provided the electrical stimulation at a pulse width of 50 μs, 400 V, and a current level ranging from 35 to 55 mA. To elicit contractions of primarily the tibialis anterior (TA) muscle, the cathode (1.5 cm diameter; Silver Mactrode, Marquette Medical Systems, Jupiter, Fla., USA) was fixed and held in place by the operator over the common peroneal nerve posterior to the fibular head, while the anode (1.0 cm diameter; Soft-E, Kendall, Chicopee, Mass., USA) was placed proximally over the TA muscle belly. The ground electrode was positioned at the distal portion of the TA. Electrode placements were marked with permanent ink to ensure accurate placement in the subsequent two visits.
Baseline experimental procedures
Each session began with the determination of the supramaximal level of stimulation to be used for the session. The isometric torque amplitude of a 1-s, 50-Hz train was monitored as the current intensity was increased incrementally. Once the 50 Hz torque level reached a plateau, the current was increased 15% in order to ensure that all further stimulation was supramaximal. Subsequently, a combined test of an isometric maximal voluntary contraction (MVC) of the dorsiflexors and a computer-regulated stimulation sequence was performed to establish baseline measures of maximal voluntary and electrically evoked torque amplitudes. To ensure accurate baseline torques, this isometric test was performed three times, with 3 min of rest separating each sequence. In addition to establishment of the baseline values, the test was used to assess the level of fatigue during exercise and recovery, and will be referred to as the fatigue test (FT). The FT consisted of an isometric MVC with an interpolated doublet (two pulses at 100 Hz), a doublet 5 s after the MVC, followed by a twitch and 1-s trains of 10, 20 and 50 Hz with 1 s of rest between each frequency (Fig. 2). Subjects held the MVC for 5 s, during which time they were provided with visual torque feedback via an oscilloscope and received strong verbal encouragement. The torque amplitudes of the interpolated doublet (T s) at the midpoint of the MVC and the subsequent doublet at rest (T r) were compared to assess voluntary activation {% activation=[1−(T s/T r)]x100%}.

Comparison of stimulated torque responses of one subject at rest (Pre-fatigue) and at 30 min of recovery (Fatigue). Each pulse or train was delivered for 1 s, with a 1-s interval between them. Low-frequency fatigue was assessed throughout exercise and recovery using the 10:50 Hz ratio at each fatigue test as a percentage of the pre-fatigue 10:50 Hz ratio
Three minutes after completion of the last baseline FT, maximal eccentric torque of the dorsiflexors was determined via five consecutive maximal eccentric contractions. The ankle began at 0° of plantar flexion and the subject maximally resisted the dynamometer lever arm as it moved at 60° s−1 to an end point of 30° of plantar flexion. Incorporating the time for the dynamometer to engage upon initiation of the contraction, each contraction lasted for 1 s and was followed by 1 s of rest (0.5 duty cycle). The 1-s rest between contractions allowed for a brief relaxation of the dorsiflexors and time for the investigator to manually return the footplate to the starting position for the subsequent eccentric contraction.
Fatigue protocols
Sessions two and three concluded with the completion of the baseline measures, whereas in session one, following a 3-min rest, the first fatigue protocol was performed. Each leg performed a protocol consisting of 150 maximal voluntary eccentric dorsiflexions. To ensure dramatically different W:R ratios, one leg performed 3 sets of 50 contractions (short rest protocol, SRP), while the other performed 15 sets of 10 contractions (long rest protocol, LRP). Regardless of the protocol, a rest period of 1 min was given between each set of contractions. As a result, the SRP had only 2 min of total rest, while the LRP had 14 min of rest. The duty cycle and range of motion for the contractions during the fatigue protocol were identical to that described for the five contractions used in the determination of maximal eccentric dorsiflexion strength. The FT was administered after the 50th(S1), 100th (S2) and 150th (S3) contraction and during the acute recovery at 1, 3, 5, 10, 15, 20, 25, and 30 min.
Data reduction and statistics
Torque data were sampled on-line using Spike2 (version 4.13, Cambridge Electronic Design, Cambridge, UK) software. Using a 12-bit A/D converter (model 1401 Plus, Cambridge Electronic Design, Cambridge, UK), the torque data were sampled at 100 Hz. During off-line analysis, Spike2 software was used to determine eccentric as well as isometric torque, electrically evoked torques, and 50 Hz half-relaxation time (50 Hz-HRT). With the exception of eccentric torque, all measures were normalized to the greatest value generated during the three fatigue tests performed prior to the fatigue protocol (PRE). A comparison of the peak torque amplitudes of the 10 Hz (T 10) and 50 Hz (T 50) responses [10:50 Hz ratio=(T 10/T50)×100%] were performed to assess LFF. To quantify LFF throughout exercise and recovery, the 10:50 Hz ratio at each FT was normalized to the pre-fatigue value (Fig. 2). The PRE value for the 10:50 Hz ratio was determined from the largest 10 Hz and 50 Hz torques. Pre-fatigue maximal eccentric torque was calculated as the average of the middle three of the five contractions performed. Eccentric torque during the fatigue protocol was calculated as the average of the 48–50th, 98–100th and 148–150th contractions. All eight subjects were used in the analysis of each measure with the exception of the 50 Hz-HRT, for which only seven subjects were used due to a technical problem during the protocol.
Normalized measures of eccentric and isometric MVCs, 10, 20, and 50 Hz, 10:50 Hz ratio, and 50 Hz-HRT each were analyzed using a two-way repeated measures analysis of variance, with protocol as one factor for comparison and time as the other. For each variable, separate analyses were performed to compare pre-fatigue versus exercise time points and pre-fatigue versus recovery time points. The exercise phase ended with the FT performed after the 150th contraction (S3), while the recovery component began with the FT measured 1 min after completion of the fatigue protocol (R1) and ended at the appropriate time point (R30 or 96 h) for each variable. Values for all data are presented in the text and figures as means with their standard error. The level of significance was set at P<0.05. When a significant main effect or an interaction was found, Tukey’s HSD post hoc tests were performed to indicate where significant differences existed.
Results
Pre-fatigue measures
There were no pre-fatigue differences in the voluntary and electrically stimulated muscle properties between protocols (Table 1). Average peak voluntary eccentric torque was approximately 140% of the peak voluntary isometric torque. Stimulation of the muscle at 50 Hz produced a torque level that was approximately 68% of the isometric MVC.
Voluntary torque
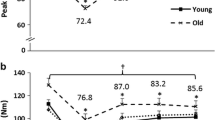
During exercise, both protocols resulted in a significant decrease in eccentric torque over time (Fig. 3). The loss of torque was non-linear, demonstrating a rapid 22% deficit relative to PRE after 50 contractions (S1), but only a further 10–14% loss over the final 100 contractions. Eccentric torque loss was similar for both protocols until the end of exercise when the loss for the SRP was 8% greater than that for the LRP. At 96 h post-exercise, the protocols had a similar and significant reduction of eccentric torque from PRE (8% SRP, 13% LRP). Although not statistically significant, isometric torque loss also tended to be greater in the SRP than LRP at the end of exercise (32% SRP, 27% LRP; Fig. 4). The acute recovery of isometric torque was similar for both protocols, but a significant deficit from PRE was still evident 30 min after the end of exercise (16% SRP, 17% LRP; Fig. 4). Recovery of isometric torque was still incomplete in both protocols at 96 h of recovery (10% torque deficit for SRP and LRP; Fig. 4). Voluntary activation, as measured by the interpolated twitch technique, was unchanged (>95%) at any point during exercise or recovery for either protocol.

Fatigue-induced changes in normalized average peak eccentric torque during exercise and at 48 and 96 h of recovery. Torque loss was significantly greater for the short rest protocol (SRP) than the long rest protocol (LRP) at the completion of exercise (S3) (+ P<0.05). All time points were significantly reduced from the pre-fatigue value (PRE) for both protocols (P<0.05). Data points of the SRP are filled, the LRP are open

Fatigue-induced changes in normalized isometric peak torque during exercise and acute as well as extended recovery. There were no significant differences between the protocols, and all time points were significantly reduced from the pre-fatigue value (PRE) for both protocols (P<0.05). Data points of the short rest protocol are filled, the long rest protocol are open
Electrically stimulated properties
The loss of stimulated 50 Hz torque paralleled the voluntary isometric torque loss at the end of exercise (SRP: 33% 50 Hz vs. 32% MVC; LRP: 24% 50 Hz vs. 27% MVC). The similarity of the decline of torque between 50 Hz and isometric MVC is another indicator that activation was unchanged in either protocol. Substantial LFF was evident at the completion of the fatiguing task, but was not different between protocols (32% SRP, 33% LRP; Fig. 5). As seen in Fig. 2, the drop in the 10:50 Hz ratio was due to a preferentially greater loss of torque at the lower frequency (10 Hz) compared to the higher frequency (50 Hz). Unlike the modest recovery of isometric torque, LFF was exacerbated throughout the 30 min acute recovery period. Thirty minutes post-exercise the 10:50 Hz ratio was further reduced to 55% and 56% of its initial value in the SRP and LRP respectively (Fig. 5). The 10:50 Hz ratio remained significantly lower than the pre-fatigue value at 96 h for both protocols (7% SRP, 9% LRP; Fig. 5).

Fatigue-induced changes in low-frequency fatigue (10:50 Hz ratio) during exercise and acute as well as extended recovery. There were no significant differences between the protocols, and all time points were significantly reduced from the pre-fatigue value (PRE) for both protocols (P<0.05). Data points of the short rest protocol are filled, the long rest protocol are open
The minimal influence of the W:R ratio was evident in the 50 Hz-HRT. With sufficient rest time during the protocol, the 50 Hz-HRT of the LRP was significantly faster than that of the SRP after 50 (S1) and 100 (S2) contractions (Fig. 6). Initially, the 50 Hz-HRT of the SRP was resistant to the decrease from the pre-fatigue HRT (faster relaxation) that occurred immediately in the LRP, but the difference between protocols became smaller and non-significant by the end of exercise. Half-relaxation time was unchanged throughout the acute recovery with both protocols leading to values that were significantly lower than the pre-fatigue value at 30 min (26% SRP, 23% LRP; Fig. 6).

Fatigue-induced changes in the 50 Hz half-relaxation (50 Hz-HRT) time during exercise and acute recovery. The long rest protocol (LRP) had a significantly faster 50 Hz-HRT in the first stages of exercise (S1 and S2) compared to the short rest protocol (SRP) (P<0.05). All recovery time points were significantly reduced from the pre-fatigue value (PRE) for both protocols (P<0.05). Data points of the SRP are filled, the LRP are open
Discussion
Contrary to our hypothesis, manipulation of the W:R ratio demonstrated a marginal effect on eccentric torque loss in high volume maximal eccentric protocols, which suggests that conventional muscle fatigue does contribute, albeit minimally, to the loss of torque. Thus, the torque loss following repetitive eccentric contractions is not solely the result of muscle damage (structural impairment). However, in accordance with our hypothesis, the LFF and the recovery of isometric MVC were insensitive to differences in total rest time (2 vs. 14 min), indicating that the persistent muscular impairment is independent of the set and repetition structure of the eccentric fatigue protocol, providing that the total contraction number and intensity are the same. Ultimately, the importance of the W:R ratio in the task-dependency model of eccentric contractions is considerably less than it is for the other contraction types.
Similar to previous investigations on the dorsiflexors (Pasquet et al. 2000; Warren et al. 2000; Birtles et al. 2003), the pre-fatigue maximal eccentric torque was 140% of the isometric torque, and there was a relatively small loss of eccentric torque (~30%) following the high volume of eccentric contractions. Compared with the dorsiflexors, repetitive eccentric contractions of the elbow flexors often results in a considerably larger loss of eccentric torque (40–50%) with fewer total contractions (Clarkson and Tremblay 1988; Nosaka et al. 2001). In the present study, the majority of the torque loss occurred within the first 50 contractions of both protocols. This greater relative decline in eccentric torque early in the protocols is consistent with the suggestion that a unique structural alteration occurs during repetitive eccentric contractions (Teague and Schwane 1995; Pasquet et al. 2000; Warren et al. 2000), as opposed to a fatigue-related torque loss induced by conventional fatigue.
Warren and colleagues (2000) found that, unlike a concentric contraction protocol that showed no concentric torque loss, repetitive maximal eccentric contractions separated by a substantial rest period (30 s) resulted in a significant reduction in eccentric torque. Because the rest period was sufficient to avoid the deleterious effects of conventional fatigue during the concentric protocol, it is reasonable to suggest that the torque loss during the eccentric protocol was entirely the result of a structural impairment (Warren et al. 2000). However, in the present study, the protocol with the greater W:R ratio (SRP) had a marginally greater loss of average peak eccentric torque than the less demanding protocol (LRP) at fatigue (S3). This finding indicates that torque loss was modestly affected by manipulation of the W:R ratio (i.e., conventional metabolic fatigue), and thus a structural mechanism was not solely responsible for the impairment. Although the effect of the W:R ratio on eccentric fatigue was not quantified, Teague and Schwane (1995) found a marginal effect of the W:R ratio on the symptoms of muscle injury following ten submaximal eccentric contractions of the elbow flexors.
The minimal impact of the W:R ratio on torque loss is underscored by the equivalent incomplete acute recoveries of isometric MVC in both protocols and the shift to a faster 50 Hz-HRT during exercise and acute recovery in both protocols. The similar persistent torque loss during acute recovery for the SRP and LRP suggests that the impairment of torque at this time is likely due to a limitation created by muscle damage, and that this damage is equivalent for both protocols (Pasquet et al. 2000). Although damage was not measured directly, previous research has indicated that the impairment of MVC torque following eccentric contractions is the best indirect measurement tool to quantify muscle damage (Warren et al. 1999).
Slowing of contractile properties (e.g., increased 50 Hz-HRT) is a common finding during high-intensity isometric and concentric fatigue protocols when limited rest is provided between contractions. This contractile slowing is often ascribed to a fatigue mechanism involving an accumulation of metabolites (Fitts 1994). The high-intensity eccentric contractions of this study resulted in a 50 Hz-HRT that was reduced (faster relaxation) in the acute recovery period compared to the pre-fatigue value in both protocols. There is presently no metabolic explanation for a decrease in HRT, but a suggested cause involves an increased rate of ATP utilization at either the actin-myosin cross bridges or at the calcium pumps of the sarcoplasmic reticulum (Vollestad et al. 1997). Regardless of the mechanism(s), the role of conventional fatigue in eccentric exercise is clearly limited because even the muscles of the SRP, designed for metabolite accumulation, shifted to a faster relaxation. Ultimately, the equivalent recovery profiles of the isometric MVC and the 50 Hz-HRT for the SRP and the LRP suggest that the mechanisms responsible for the torque loss during each protocol were similar, despite the dramatic differences in the W:R ratio.
Low-frequency fatigue has often been observed following eccentric contractions (Jones 1981, 1996), but this study was the first to examine the influence of the W:R ratio. The equivalent decline in the normalized 10:50 Hz ratio following the SRP and LRP seen in this study suggests that the LFF was insensitive to manipulation of the W:R ratio. Previous studies have identified that eccentric contractions induce considerably more LFF than concentric contractions (Davies and White 1981; Edwards et al. 1981; Newham et al. 1983), and the degree of LFF induced is largely related to the number of eccentric contractions performed (Chin et al. 1997; Willems and Stauber 2003). The development of LFF is often ascribed to a dysfunction of E-C coupling, likely due to a disruption of the t-tubule and SR link (Lannergren et al. 1996; Ingalls et al. 1998; Warren et al. 2002; Westerblad and Allen 2002). Given that an equivalent degree of LFF was induced in both the SRP and LRP, the greater decline in torque at fatigue in the SRP versus the LRP was not sufficient to minimize the muscle damage and thus alter the associated E-C coupling failure. Thus, the E-C coupling failure that accompanies repetitive eccentric contractions is more likely related to the intensity and number of contractions performed, rather than manipulation of the set and repetition structure (W:R ratio) of the fatigue protocol.
Low-frequency fatigue can be due solely to reduced torques at the lower frequencies with no change in the response at high frequencies of stimulation (Chin et al. 1997). This was not the case in the present study because the 50 Hz as well as the 10 Hz stimulated torque were significantly impaired at 96 h, which suggests the contribution of a persistent structural impairment to LFF. Additionally, irrespective of the protocol, isometric peak torque as well as eccentric average peak torque remained reduced after 96 h of recovery. These sustained impairments of MVC and 50 Hz stimulated torque suggest that the SRP and LRP have significant structural damage beyond that indicated by the decrease in the 10:50 Hz ratio. The repair and regeneration of this damage to the contractile elements or the SR is a slow process, set on a time course of protein turnover rather than metabolite resynthesis (Clarkson and Tremblay 1988; Jones 1996).
In conclusion, the present study contributes two novel findings concerning the influence of the W:R ratio on eccentric contractions and the associated fatigue. Firstly, as postulated, the degree of LFF and the recovery of isometric MVC are insensitive to total rest time provided during moderately fatiguing eccentric exercise. Secondly, in contrast to our hypothesis, the task-dependency paradigm of eccentric contractions is marginally associated with the W:R ratio. Although only indirect measures of muscle damage and metabolic changes were made in the present study, torque loss appears to be influenced mainly by muscle damage but does involve a metabolic component associated with conventional fatigue. This marginal influence of conventional fatigue represents an important difference from the task-dependent nature of concentric and isometric contractions and is another unique feature of eccentric contractions.
References
Bigland-Ritchie B, Rice CL, Garland SJ, Walsh ML (1995) Task-dependent factors in fatigue of human voluntary contractions. In: Gandevia SC, Enoka RM, McComas AJ, Stuart DG, Thomas CK (eds) Fatigue. Plenum, New York, pp 361–380
Birtles DB, Rayson MP, Jones DA, Padhiar N, Casey A, Newham DJ (2003) Effect of eccentric exercise on patients with chronic exertional compartment syndrome. Eur J Appl Physiol 88:565–571
Brown SJ, Child RB, Day SH, Donnelly AE (1997) Exercise-induced skeletal muscle damage and adaptation following repeated bouts of eccentric muscle contractions. J Sports Sci 15:215–222
Chin ER, Balnave CD, Allen DG (1997) Role of intracellular calcium and metabolites in low-frequency fatigue of mouse skeletal muscle. Am J Physiol 41:C550–C559
Clarkson PM, Tremblay I (1988) Exercise-induced muscle damage, repair, and adaptation in humans. J Appl Physiol 65:1–6
Clarkson PM, Hubal MJ (2002) Exercise-induced muscle damage in humans. Am J Phys Med Rehabil 81:S52–S69
Davies CTM, White MJ (1981) Muscle weakness following eccentric work in man. Pflugers Arch 392:168–171
Edwards RHT, Mills KR, Newham DJ (1981) Greater low frequency fatigue produced by eccentric than concentric muscle contractions. J Physiol (Lond) 317:17P
Enoka RM, Stuart DG (1992) Neurobiology of muscle fatigue. J Appl Physiol 72:1631–1648
Fitts RH (1994) Cellular mechanisms of muscle fatigue. Physiol Rev 74:49–94
Ingalls CP, Warren GL, Williams JH, Ward CW, Armstrong RB (1998) E-C coupling failure in mouse EDL muscle after in vivo eccentric contractions. J Appl Physiol 85:58–67
Jones DA (1981) Muscle fatigue due to changes beyond the neuromuscular junction. Ciba Found Symp 82:178–196
Jones DA (1996) High-and low-frequency fatigue revisited. Acta Physiol Scand 156:265–70
Lannergren J, Westerblad H, Bruton JD (1996) Slow recovery of force in single skeletal muscle fibres. Acta Physiol Scand 156:193–202
Lieber RL, Friden J (2002) Mechanisms of muscle injury gleaned from animal models. Am J Phys Med Rehabil 81:S70–S79
Newham DJ, Mills KR, Quigley BM, Edwards RH (1983) Pain and fatigue after concentric and eccentric muscle contractions. Clin Sci 64:55–62
Nosaka K, Sakamoto K, Newton M, Sacco P (2001) The repeated bout effect of reduced-load eccentric exercise on elbow flexor muscle damage. Eur J Appl Physiol 85:34–40
Pasquet B, Carpentier A, Duchateau J, Hainaut K (2000) Muscle fatigue during concentric and eccentric contractions. Muscle Nerve 23:1727–1735
Teague BN, Schwane JA (1995) Effect of intermittent eccentric contractions on symptoms of muscle microinjury. Med Sci Sports Exerc 27:1378–1384
Vollestad NK, Sejersted I, Saugen E (1997) Mechanical behaviour of skeletal muscle during intermittent voluntary isometric contractions in humans. J Appl Physiol 83:1557–1565
Warren GL, Hayes DA, Lowe DA, Armstrong RB (1993) Mechanical factors in the initiation of eccentric contraction-induced injury in rat soleus muscle. J Physiol (Lond) 464:457–475
Warren GL, Lowe DA, Armstrong RB (1999) Measurement tools used in the study of eccentric contraction-induced injury. Sports Med 27:43–59
Warren GL, Hermann KM, Ingalls CP, Masselli MR, Armstrong RB (2000) Decreased EMG median frequency during a second bout of eccentric contractions. Med Sci Sports Exerc 32:820–829
Warren GL, Ingalls CP, Lowe DA, Armstrong RB (2002) What mechanisms contribute to the strength loss that occurs during and in the recovery from skeletal muscle injury? J Orthop Sports Phys Ther 32:58–64
Westerblad H, Allen DG (2002) Recent advances in the understanding of skeletal muscle fatigue. Curr Opin Rheumatol 14:648–652
Willems ME, Stauber WT (2003) Low-frequency fatigue increases with the number of stretches of activated rat skeletal muscles [abstract]. Med Sci Sports Exerc 35:S144
Acknowledgements
This work is supported in part by the Natural Sciences and Engineering Research Council of Canada, the Canadian Institutes of Health Research, and the Canada Foundation for Innovation. The experiments performed comply with the current laws of Canada regarding experimentation on human subjects.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
McNeil, C.J., Allman, B.L., Symons, T.B. et al. Torque loss induced by repetitive maximal eccentric contractions is marginally influenced by work-to-rest ratio. Eur J Appl Physiol 91, 579–585 (2004). https://doi.org/10.1007/s00421-003-0996-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-003-0996-y