
Abstract
Focal cortical dysplasias (FCD) represent a frequent finding in patients with chronic intractable epilepsy. Neuropathological hallmarks include localized dyslamination of the neocortex and neuronal heterotopias in white matter. Balloon cells, similar to those occurring in cortical tubers of patients with tuberous sclerosis (TSC) are observed in numerous patients. These lesions were classified as FCD type IIb (FCD IIb). Recent findings indicate an accumulation of TSC1 polymorphisms as well as loss of heterozygosity (LOH) and/or microsatellite instability (MSI) at the TSC1 locus on chromosome 9q in FCD IIb. Here, we tested the hypothesis of whether chromosomal instability constitutes a genome-wide phenomenon in this patient cohort. Seven microsatellite markers based on a reference panel recommended by the international workshop on microsatellite instability were analyzed in 14 surgical FCD IIb specimens. DNA from single laser-microdissected cells, i.e., balloon cells versus control neurons obtained from adjacent cortex was harvested for PCR amplification and subsequent fluorescent fragment length gel electrophoresis. Our analysis revealed only rare instances of LOH and MSI at genomic loci on 2p and 17q, whereas no alterations were found at informative markers on chromosomes 1p, 5q and 18q. In addition, no loss of repair protein expression (MSH2 or MLH1) has been identified in balloon cell nuclei of FCD IIb specimens. The present data suggest solitary LOH and MSI events at genomic localizations others than the TSC1 locus to occur in FCD IIb. Our findings lend further support to the hypothesis that the molecular pathogenesis of FCD IIb is associated with TSC1.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cortical dysplasias display a broad spectrum of structural alterations, which appear to result from aberrant proliferation, migration, differentiation and apoptosis of neural precursors and neuroglial cells during cortical development [13, 29]. Due to distinct clinical and phenotypic characteristics, a subtype linked to chronic intractable epilepsy has been determined as focal cortical dysplasia of Taylor’s balloon cell type [31]. In a recent classification system of epilepsy-associated cortical malformations, this variant was referred to as focal cortical dysplasia (FCD) type IIb [25]. In our series, FCD IIb was identified by a transmantle-like hyperintensive lesion on magnetic resonance images, characteristic histopathological findings and favorable clinical outcome [33]. Striking histomorphological similarities between FCD IIb and tuberous sclerosis (TSC)-associated cortical tubers, i.e., dysplastic neurons and balloon cells, have suggested that the TSC1 and TSC2 genes are involved in the molecular pathogenesis of FCD IIb.
Previous loss of heterozygosity (LOH) studies revealed allelic losses at the TSC1 (hamartin; chromosome 9q34) and TSC2 (tuberin, chromosome 16p13.3) loci in lesions of TSC patients and in sporadic tumors of individuals not suffering from TSC. Hamartin and tuberin have been suggested to constitute a tumor suppressor mechanism [14, 16, 34]. Recently, we have observed a significant increase in the frequency of TSC1 polymorphisms and chromosomal instability [LOH and microsatellite instability (MSI)] at the TSC1 locus in FCD IIb of patients with chronic pharmaco-resistant epilepsy [5]. The combination of LOH at the TSC1 locus and coding polymorphisms of the second allele in FCD IIb represented an intriguing finding with respect to the two-hit hypothesis for the inactivation of tumor suppressor genes [21]. Other studies pointed towards differences between FCD IIb and TSC in the expression of excitatory and inhibitory neurotransmitter pathways [10, 11, 36].
A further, novel observation was the instability of individual microsatellite markers at the TSC1 locus in balloon cells obtained from surgical specimens of FCD IIb patients [5]. Microsatellites constitute short, tandemly repeated nucleotide sequences widely distributed throughout the genome, which appear particularly prone to replication errors [20]. Nucleotide repeat length alterations are used as diagnostic criteria for the replication error phenotype in tumor DNA and referred to as MSI [6, 32]. MSI is most commonly caused by a deficiency of DNA mismatch repair (MMR) genes, i.e., MLH1 or MSH2 [2]. MSI represents a hallmark of the hereditary nonpolyposis colorectal carcinoma (HNPCC) syndrome with a high incidence of colorectal tumors [1]. A subpopulation of HNPCC patients develop gliomas. Several studies have addressed MSI in gliomas using different panels of microsatellite loci. Variable prevalences of the defective repair phenotype have been reported in these studies [12, 30, 38]. Referring to National Cancer Institute consensus microsatellite markers [6], MSI has been detected in a substantial fraction of highly differentiated, epilepsy-associated glio-neuronal neoplasms, i.e., gangliogliomas [2]. This appeared particularly remarkable since MSI has been generally regarded as a phenomenon primarily related to malignant neoplasms.
To answer the question of whether chromosomal instability in FCD IIb is a genome-wide phenomenon, as has been demonstrated in gangliogliomas, or a genomic alteration restricted to the region of the TSC1 locus [9], we have analyzed a set of seven microsatellite markers of different genomic localizations as well as the expression of the MMR proteins MLH1 and MSH2 in 14 FCD IIb lesions of patients with intractable epilepsy.
Materials and methods
Surgical specimens
Biopsy samples were obtained from 14 patients with chronic pharmaco-resistant epilepsy, who underwent surgical treatment in the Epilepsy Surgery Programs at the University of Bonn Medical Center and the Epilepsy Center at the Bethel Hospital. In all patients, surgical removal of the FCD IIb was necessary to achieve seizure control after presurgical evaluation [22]. Informed and written consent was obtained from all patients for additional studies. Two of the patients presented with multiple stigmata of TSC, i.e., skin lesions (facial angiofibromas, hypomelanotic macules), retinal hamartomas or cardiac rhabdomyoma, and were therefore classified as manifest TSC [27]. All procedures were conducted in accordance with the Declaration of Helsinki and approved by the ethics committees of the respective institutions. Surgical specimens were fixed in formaldehyde overnight and embedded into paraffin. All FCD IIb were reviewed by experienced neuropathologists at the Bonn or Bethel epilepsy centers. The FCD IIb specimens have been previously analyzed for genomic alterations at the TSC1 locus [5].
Laser-assisted microdissection and PCR
UV laser microbeam technology based on a nitrogen laser [28] (PALM, Bernried) was applied for microdissection and harvesting of balloon cells of the FCD IIb lesion and adjacent normal brain neuronal cellular elements. Balloon cells could be differentiated according to their nuclear and cellular morphologies after hematoxylin-eosin staining of 10-µm paraffin sections. Ten cells were harvested and sampled for each PCR reaction. Cells from normal cortex outside the FCD IIb served as controls for SSCP analysis. Harvested samples were further processed as recommended by the manufacturer. The DNA isolation was carried out according to the manufacturer’s protocol of the QIAampDNA Mini Kit (Qiagen, Hilden, Germany).
PCR/microsatellite marker analysis
PCRs were carried out separately for DNA obtained from balloon cells vs control cell samples. LOH/MSI analysis was performed using the seven microsatellite markers MYCL1 (1p32), D2S123 (2p16), BAT26 (2p16.3-p2), BAT25 (4q12), D5S346 (5q21/22), D17S250 (17q11.2-q12) and D18S58 (18q22.3) [23]. A total reaction volume of 20 µl containing the previously extracted DNA from laser-microdissected cellular components, 10× PCR buffer (−MgCl2), 0.2 µM dNTPs, 0.075 U Taq polymerase, 1.5 mM MgCl2 and 1 pmol of each primer was used. Primer combinations were applied as outlined in Table 1. Amplification was performed in an automated thermocycler (GeneAmp 2700, Applied Biosystems) for 40 cycles at 94°C for 30 s, annealing for 30 s at 50/55/60°C, and 72°C for 1 min. An initial 2-min step at 94°C and a final 10-min step at 72°C were added. For DNA fragment analysis using the PRISM 310 (ABI, Foster City, CA), 1 μl of each PCR product was mixed with 12 μl formamide and 1 μl GeneScan-350 size standard to a total volume of 18 μl and processed as described above [23]. The status for each microsatellite marker in an individual FCD IIb was determined by at least two replicates starting at the laser-microdissection level. The Chi-square test was used for statistical comparison of MSI frequencies in informative cases of microsatellites at chromosomes 1, 2, 5, 17 and 18 versus the previous data from our group on MSI for markers D9S302, D9S303 and D9S319 at the TSC1 locus [5].
Immunohistochemistry
Detection of the mismatch repair proteins MSH2 and MLH1 was carried out with mouse anti-human monoclonal antibodies (MSH2 clone, G219-1129; MLH1 clone, G618-15, PharMingen, San Diego, CA) as described in detail previously [23]. In brief, paraffin sections of FCD IIb (n=11) were deparaffinized in xylene, rehydrated in graded alcohols, and washed in TRIS buffer. Heat-induced epitope retrieval (600 W microwave treatment twice for 15 min in prewarmed 10 mM sodium citrate buffer, pH 6) was applied during MSH2 and MLH1 staining procedures. Primary antibodies were added (dilution: MSH2 1:50, MLH1 1:75) before incubation of slides overnight at 4°C, which were subsequently processed on an immunostainer (TechMate 500, Dako, Hamburg, Germany) and visualized using the avidin-biotin-complex method. Specificity of the antibodies has been tested previously [23]. Hematoxylin-counterstained sections were mounted in aqueous mounting media and analyzed by standard light microscopy. Neurons, glia and endothelial cells in normal CNS tissue adjacent to the FCD IIb served as internal controls. FCD IIb cellular components were considered to lack repair protein expression if nuclei were deficient of immunostaining for MSH2 or MLH1 in contrast to positive internal control nuclei.
Results
The microsatellite allelic mobility shift analysis of laser-microdissected cellular components from FCD IIb of 14 individuals revealed the microsatellite marker status summarized in Fig. 1. Sequence alterations were found in three patients: MSI was detected at D17S250 in the FCD IIb of patient 8 (17q11.2-q12). LOH events at D2S123 (2p16) were observed in two FCD IIb (patients 3 and 5). In the FCD IIb of patient 5, the LOH at D2S123 was found in concert with MSI at D17S250. This was the only case in which such co-occurrence at two loci was noted in a single FCD IIb in the present series. Taking previously reported microsatellite analysis at the TSC1 locus into account [5], this was one of only three FCD IIb cases with MSI for more than one microsatellite marker at 9q. Two patients with manifest TSC showed no genomic alterations with the present microsatellite markers (Fig. 1). The comparison of genomic instability (LOH and MSI) at the informative markers in the present study with previously reported results of our group for microsatellites at the TSC1 locus [5] demonstrated that chromosomal alterations were most frequently observed at chromosome 9q (Chi-square test P<0.001; Fig. 2).

MSI and LOH status obtained from 14 epileptic patients with FCD IIb using microsatellite markers MYCL1, D2S123, BAT26, BAT25, D5S346, D17S250 and D18S58. A Genomic instability is observed in only three FCD IIb of the present series. Clinically manifest TSC patients (+) are also listed and do not exhibit genomic instability in the microsatellite markers (gray circles non informative, white and black circles maintenance of heterozygosity, black circles LOH, circles with bar MSI). B Representative MSI observed at D17S250. C LOH at D2S123 as found in two FCD IIb (MSI microsatellite instability, LOH loss of heterozygosity, FCD focal cortical dysplasia, TSC tuberous sclerosis)

Frequency of genomic instability events in FCD IIb at different chromosomal localizations. The percentages of genomic instability are based only on informative FCD IIb cases for each marker. Therefore, BAT26 and BAT25 microsatellites are not listed. Substantially higher numbers of microsatellite marker alterations (MSI or LOH) can be observed in the 9q region of the TSC1 locus, i.e., markers D9S302, D9S303 and D9S319 as determined previously by our group [5]
Using an immunohistochemical approach, we have also studied whether defective rough endoplasmic reticulum due to an impairment of MSH2 and MLH1 may be related to MSI. However, no significant reduction or loss of expression for MSH2 and MLH1 was found in FCD IIb (Fig. 3).

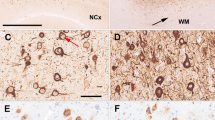
Immunohistochemical detection of the mismatch repair proteins MSH2 and MLH1 in FCD IIb. A An FCD IIb that exhibits the characteristic composition by dysplastic neuronal elements and balloon cells (hematoxylin-eosin staining). Immunohistochemical detection of the mismatch repair proteins MSH2 (B) and MLH1 (C) in FCD IIb components. Note that intensive immunolabeling can be observed in dysplastic neuronal and balloon cell nuclei for MSH2 and MLH1. A, B ×20; C ×40
Discussion
TSC1 and TSC2 represent tumor suppressor genes affected in different neurocutaneous lesions. The general pathogenetic concept for the action of tumor suppressor genes corresponds to the two-hit hypothesis for the inactivation of tumor suppressor genes, i.e., LOH of one allele and associated mutation in the second allele [21]. Mutations of TSC1 and TSC2 were found in patients with both sporadic and familial TSC [19, 37]. The frequency of LOH appeared to be significantly lower in brain lesions compared to other malformations of TSC patients [17].
In a laser-microdissection based study, LOH at the TSC1 locus was observed in 11 out of 24 patients with FCD IIb [5]. Of these 11 FCD IIb cases, 6 showed LOH in combination with sequence alterations in exon 14/intron 13, exon 17 or exon 22 of the TSC1 gene. In addition to LOH, instability of microsatellite markers was observed in 6 FCD IIb patients [5]. The remarkably high incidence of chromosomal instability, i.e., LOH and MSI, at 9q raised the question of whether chromosomal instability in FCD IIb is a widespread genomic phenomenon as has been demonstrated in glioneuronal tumors [2] or a more specific alteration confined to the region of the TSC1 locus [9].
Our present results showed chromosomal instability to occur only rarely at chromosomes 2 and 17. No LOH or MSI has been observed in informative markers on chromosomes 1, 5 and 18. In concert with a previous report from our laboratory on MSI and LOH at 9q [5], the present data using a microsatellite marker set with random genomic distribution underline that LOH and MSI preferentially affect the TSC1 locus on chromosome 9q in FCD IIb.
In gangliogliomas, i.e., glioneuronal tumors of non-TSC patients, several studies have determined the genomic status at 16p13.3 and at 9q34. No LOH at the TSC1 or TSC2 loci had been identified in 10 sporadic gangliogliomas [35]. Other investigators described 4 LOH events in 15 gangliogliomas (WHO grade I) [26], with a higher incidence of LOH at the TSC2 than TSC1 locus. These results are in good agreement with previous findings of our group demonstrating that gangliogliomas exhibit frequent polymorphisms and a single mutation in the TSC2 gene [4]. An anaplastic ganglioglioma was detected in the Eker mutant rat, a strain known to harbor genetic alterations of the TSC2 gene, whereas MSI was not observed in Eker rat tumors [18, 24]. Recently, a high incidence of MSI has been reported in gangliogliomas using the microsatellite markers BAT25 and BAT26 [2]. Also with respect to MSI, gangliogliomas appear distinct from FCD IIb. In contrast to the finding in gangliogliomas, BAT25 and BAT26 microsatellites were non-informative with respect to LOH, and did not exhibit MSI in the FCD IIb cohort of the present study (Fig. 1).
Regarding manifest TSC, LOH for alleles at 16p13.3 in brain lesions and at 9q34 in extracerebral manifestations has been described in lesions from individuals with TSC [7, 14, 15]. In brain lesions of TSC patients, TSC2 is more frequently altered than TSC1 [3, 8, 17]. We did not observe MSI or LOH in the two TSC patients by the microsatellite marker set employed in the present study.
Taken together, microsatellite analyses suggest that FCD IIb with chromosomal instability at the TSC1 gene locus is molecularly distinct from brain lesions in manifest TSC patients and from epilepsy-associated glioneuronal tumors. Nevertheless, FCD IIb characterized by a high incidence of TSC1 sequence alterations in concert with chromosomal instability at 9q, but not other genomic regions, appears to be pathogenetically related to TSC.
References
Aaltonen LA, Peltomaki P, Leach FS, Sistonen P, Pylkkanen L, Mecklin JP, Jarvinen H, Powell SM, Jen J, Hamilton SR, Peterson GM, Kinzler KW, Vogelstein B, Chapelle A de la (1993) Clues to the pathogenesis of familial colorectal cancer. Science 260:812–816
Alonso M, Hamelin R, Kim M, Porwancher K, Sung T, Parhar P, Miller D, Newcomb EW (2001) Microsatellite instability occurs in distinct subtypes of pediatric but not adult central nervous system tumors. Cancer Res 61:2124–2128
Al-Saleem T, Wessner LL, Scheithauer BW, Patterson K, Roach ES, Dreyer SJ, Fujikawa K, Bjornsson J, Bernstein J, Henske EP (1998) Malignant tumors of the kidney, brain, and soft tissues in children and young adults with the tuberous sclerosis complex. Cancer 83:2208–2216
Becker AJ, Löbach M, Klein H, Normann S, Nöthen MM, Deimling A von, Mizuguchi M, Elger CE, Schramm J, Wiestler OD, Blümcke I (2001) Mutational analysis of TSC1 and TSC2 genes in gangliogliomas. Neuropathol Appl Neurobiol 27:105–114
Becker AJ, Urbach H, Scheffler B, Baden T, Normann S, Lahl R, Pannek HW, Tuxhorn I, Elger CE, Schramm J, Wiestler OD, Blümcke I (2002) Focal cortical dysplasia of Taylor’s balloon cell type: mutational analysis of the TSC1 gene indicates a pathogenic relationship to tuberous sclerosis. Ann Neurol 52:29–37
Boland CR, Thibodeau SN, Hamilton SR, Sidransky D, Eshleman JR, Burt RW, Meltzer SJ, Rodriguez-Bigas MA, Fodde R, Ranzani N, Srivastava S (1998) A National Cancer Institute workshop on microsatellite instability for cancer detection and familial predisposition: development of international criteria for the determination of microsatellite instability in colorectal cancer. Cancer Res 58:5248–5257
Carbonara C, Longa L, Grosso E, Borrone C, Garre MG, Brisigotti M, Migone N (1994) 9q34 loss of heterozygosity in a tuberous sclerosis astrocytoma suggests a growth suppressor-like activity also for the TSC1 gene. Hum Mol Genet 3:1829–1832
Carbonara C, Longa L, Grosso E, Mazzucco G, Borrone C, Garre ML, Brisigotti M, Filippi G, Scabar A, Giannotti A, Falzoni P, Monga G, Garini G, Gabrielli M, Riegler P, Danesino C, Ruggieri M, Magro G, Migone N (1996) Apparent preferential loss of heterozygosity at TSC2 over TSC1 chromosomal region in tuberous sclerosis hamartomas. Genes Chromosomes Cancer 15:18–25
Crino PB (2002) Bourneville, Taylor: a developing story? Ann Neurol 52:6–8
Crino PB, Trojanowski JQ, Dichter MA, Eberwine J (1996) Embryonic neuronal markers in tuberous sclerosis: single-cell molecular pathology. Proc Natl Acad Sci USA 93:14152–14157
Crino PB, Duhaime AC, Baltuch G, White R (2001) Differential expression of glutamate and GABA-A receptor subunit mRNA in cortical dysplasia. Neurology 56:906–913
Dams E, Van de Kelft EJ, Martin JJ, Verlooy J, Willems PJ (1995) Instability of microsatellites in human gliomas. Cancer Res 55:1547–1549
Golden JA (2001) Cell migration and cerebral cortical development. Neuropathol Appl Neurobiol 27:22–28
Green AJ, Johnson PH, Yates JR (1994) The tuberous sclerosis gene on chromosome 9q34 acts as a growth suppressor. Hum Mol Genet 3:1833–1834
Green AJ, Smith M, Yates JR (1994) Loss of heterozygosity on chromosome 16p13.3 in hamartomas from tuberous sclerosis patients. Nat Genet 6:193–196
Henske EP, Neumann HP, Scheithauer BW, Herbst EW, Short MP, Kwiatkowski DJ (1995) Loss of heterozygosity in the tuberous sclerosis (TSC2) region of chromosome band 16p13 occurs in sporadic as well as TSC-associated renal angiomyolipomas. Genes Chromosomes Cancer 13:295–298
Henske EP, Scheithauer BW, Short MP, Wollmann R, Nahmias J, Hornigold N, Slegtenhorst M van, Welsh CT, Kwiatkowski DJ (1996) Allelic loss is frequent in tuberous sclerosis kidney lesions but rare in brain lesions. Am J Hum Genet 59:400–406
Hino O, Kobayashi E, Hirayama Y, Kobayashi T, Kubo Y, Tsuchiya H, Kikuchi Y, Mitani H (1995) Molecular genetic basis of renal carcinogenesis in the Eker rat model of tuberous sclerosis (Tsc2). Mol Carcinog 14:23–27
Jones AC, Shyamsundar MM, Thomas MW, Maynard J, Idziaszczyk S, Tomkins S, Sampson JR, Cheadle JP (1999) Comprehensive mutation analysis of TSC1 and TSC2-and phenotypic correlations in 150 families with tuberous sclerosis. Am J Hum Genet 64:1305–1315
Kang GH, Yoon GS, Lee HK, Kwon YM, Ro JY (1999) Clinicopathologic characteristics of replication error-positive gastric carcinoma. Mod Pathol 12:15–20
Knudson AG (1996) Hereditary cancer: two hits revisited. J Cancer Res Clin Oncol 122:135–140
Kral T, Clusmann H, Urbach J, Schramm J, Elger CE, Kurthen M, Grunwald T (2002) Preoperative evaluation for epilepsy surgery (Bonn Algorithm). Zentralbl Neurochir 63:106–110
Mathiak M, Rutten A, Mangold E, Fischer HP, Ruzicka T, Friedl W, Propping P, Kruse R (2002) Loss of DNA mismatch repair proteins in skin tumors from patients with Muir-Torre syndrome and MSH2 or MLH1 germline mutations: establishment of immunohistochemical analysis as a screening test. Am J Surg Pathol 26:338–343
Mizuguchi M, Takashima S, Yamanouchi H, Nakazato Y, Mitani H, Hino O (2000) Novel cerebral lesions in the Eker rat model of tuberous sclerosis: cortical tuber and anaplastic ganglioglioma. J Neuropathol Exp Neurol 59:188–196
Palmini A, Lüders HO (2002) Classification issues in malformations caused by abnormalities of cortical development. Neurosurg Clin N Am 13:1–16
Parry L, Maynard JH, Patel A, Hodges AK, Deimling A von, Sampson JR, Cheadle JP (2000) Molecular analysis of the TSC1 and TSC2 tumour suppressor genes in sporadic glial and glioneuronal tumours. Hum Genet 107:350–356
Roach ES, Gomez MR, Northrup H (1998) Tuberous sclerosis complex consensus conference: revised clinical diagnostic criteria. J Child Neurol 13:624–628
Schütze K, Lahr G (1998) Identification of expressed genes by laser-mediated manipulation of single cells. Nat Biotechnol 16:737–742
Schwartzkroin PA, Walsh CA (2000) Cortical malformations and epilepsy. Ment Retard Dev Disabil Res Rev 6:268–280
Sobrido MJ, Pereira CR, Barros F, Forteza J, Carracedo A, Lema M (2000) Low frequency of replication errors in primary nervous system tumours. J Neurol Neurosurg Psychiatry 69:369–375
Taylor DC, Ounsted C (1971) Biological mechanisms influencing the outcome of seizures in response to fever. Epilepsia 12:33–45
Thibodeau SN, Bren G, Schaid D (1993) Microsatellite instability in cancer of the proximal colon. Science 260:816–819
Urbach H, Scheffler B, Heinrichsmeier T, Oertzen J von, Kral T, Wellmer J, Schramm J, Wiestler OD, Blümcke I (2002) Focal cortical dysplasia of Taylor’s balloon cell type: a clinicopathological entity with characteristic neuroimaging and histopathological features, and favorable postsurgical outcome. Epilepsia 43:33–40
Van Slegtenhorst M, Hoogt R de, Hermans C, Nellist M, Janssen B, Verhoef S, Lindhout D, Ouweland van den A, Halley D, Young J, Burley M, Jeremiah S, Woodward K, Nahmias J, Fox M, Ekong R, Osborne J, Wolfe J, Povey S, Snell RG, Cheadle JP, Jones AC, Tachataki M, Ravine D, Sampson JR, Reeve MP, Richarson P, Wilmer F, Munro C, Hawkins TL, Sepp T, Ali JBM, Ward S, Green AJ, Yates JRW, Kwiatkowski J, Henske EP, Short MP, Haines JH, Jozwiak S, Kwiatkowski DJ (1997) Identification of the tuberous sclerosis gene TSC1 on chromosome 9q34. Science 277:805–808
Von Deimling A, Fimmers R, Schmidt MC, Bender B, Fassbender F, Nagel J, Jahnke R, Kaskel P, Duerr EM, Koopmann J, Maintz D, Steinbeck S, Wick W, Platten M, Müller DJ, Przkora R, Waha A, Blümcke B, Wellenreuther R, Meyer-Puttlitz B, Schmidt O, Mollenhauer J, Poustka A, Stangl AP, Lenartz D, Ammon K von, Henson JW, Schramm J, Louis DN, Wiestler OD (2000) Comprehensive allelotype and genetic anaysis of 466 human nervous system tumors. J Neuropathol Exp Neurol 59:544–558
White R, Hua Y, Scheithauer B, Lynch DR, Henske EP, Crino PB (2001) Selective alterations in glutamate and GABA receptor subunit mRNA expression in dysplastic neurons and giant cells of cortical tubers. Ann Neurol 49:67–78
Wilson PJ, Ramesh V, Kristiansen A, Bove C, Jozwiak S, Kwiatkowski DJ, Short MP, Haines JL (1996) Novel mutations detected in the TSC2 gene from both sporadic and familial TSC patients. Hum Mol Genet 5:249–256
Zhu J, Guo SZ, Beggs AH, Maruyama T, Santarius T, Dashner K, Olsen N, Wu JK, Black P (1996) Microsatellite instability analysis of primary human brain tumors. Oncogene 12:1417–1423
Acknowledgements
We thank Atakan Aydin for excellent technical support. Our work is funded by Deutsche Forschungsgemeinschaft (SFB TR3), BMBF (NGFN TP 11 and 15), Deutsche Krebshilfe and the BONFOR program of the University of Bonn Medical Center.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fassunke, J., Blümcke, I., Lahl, R. et al. Analysis of chromosomal instability in focal cortical dysplasia of Taylor’s balloon cell type. Acta Neuropathol 108, 129–134 (2004). https://doi.org/10.1007/s00401-004-0874-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00401-004-0874-7