
Abstract
Background
Many efforts have been performed in the last decade to accomplish the genomic and proteomic characterization of pediatric adamantinomatous craniopharyngioma with the purpose to elucidate the molecular mechanisms underlying the onset and development of this pediatric brain tumor, its high recurrence rate, and, although classified as a histologically benign neoplasm, its aggressive behavior.
Methods
The focus of this review is to perform the new comparison of the proteomic profiles of the solid component and the intracystic fluid of adamantinomatous craniopharyngioma based on our previous results, obtained by both the top-down and the bottom-up proteomic approaches, to disclose differences and similarities, and to discuss the results in the context of the most recent literature.
Results and conclusions
Proteins and peptides identified in the cyst fluid and in the solid component of adamantinomatous craniopharyngioma (AC) include beyond markers of inflammation (i.e., alpha-defensins), proteins involved in cell migration and protein degradation (i.e., beta-thymosin and ubiquitin peptides), whose main role might be in tumor growth and infiltration of the surrounding neural structures. These last appeared different in the solid components compared with the cyst fluid, missing their terminal part in the solid tissue, a feature generally associated to malignancies, which might represent a distinct molecular site for an aggressive behavior of AC.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In spite of the benign histological characteristics (WHO I), craniopharyngioma in children still represents a management challenge, namely because of the need to achieve the cure of the tumor and to preserve the patient development at the same time.
Biologically, almost all pediatric craniopharyngiomas show the features of the adamantinomatous type (AC). The true cellular origin of AC remains to be definitively confirmed, but its location in the sellar/suprasellar region, combined with the tumor epithelial features, implies that AC arises from early pituitary progenitors in Rathke’s pouch, the embryonic precursor of the anterior pituitary gland. Macroscopically, solid and cystic components, with micro- and/or gross calcifications, are the main findings. The solid portion is often quite amorphous and soft with numerous micro-calcifications, whereas the cyst fluid often described as “machine oil,” has a viscous consistence that is reflective of the content of cholesterol crystals and of the cellular debris within the fluid.
The best treatment option for AC is still a matter of debate. Beyond the different perspectives based on the prominence of a solid versus cystic nature of the lesion, the patient’s age and his/her endocrinological status assume a prominent role. It is actually almost universally agreed inside the neurosurgical community that the therapeutic goal has to be to treat the child with the least possible long-term (i.e., endocrinological, visual) morbidity and the longest possible disease control. Tumor removal has increased surgical risks, particularly in the case of intricate relationships of craniopharyngiomas with surrounding vascular and nervous tissues, which may not always be predicted on pre-operative imaging and might not be solved in all cases, in spite of the availability of various surgical approaches (i.e., micro-surgical, endoscopic transsphenoidal, endoscopic transventricular) and a continuous increasing number of available surgical tools. The most popular alternatives to surgery are represented for extensively cystic tumors by the intracystic injection of bleomycin or, more recently, of alpha-interferon. Secondary effects of drug leakage seem to be not significant with alpha-interferon compared with bleomycin. Both these treatments share the same encouraging results in the first years of follow-up (until 5–7 years) but with a criticism about the possibility that they can represent a permanent treatment (currently possible in no more than 30% of cases). Conformal radiotherapy and gamma-knife are a further option that has to be considered particularly to control small solid tumor residuals/recurrences, if their surgical removal is expected to carry an increased risk for their adherence to eloquent vascular and/or neural structures. In this context, the previously documented risks of whole brain irradiation are significantly reduced, nowadays, specifically for the focal restrictions and modulation of the radiation doses. However, in the late sequelae on the hypothalamic/pituitary axis, optic pathways (and cognitive functions) remain quite relevant.
On these grounds, we could conclude that the optimal management of this tumor is still under investigation. A benefit on AC knowledge certainly comes from the proteomic analysis of this tumor, which aims at reaching an individualized molecular perspective of cure.
Very recent reviews illustrate the advances of molecular knowledge of AC tumor achieved by both transcriptomics and proteomics studies [1, 2] and their evaluation to develop targeted therapies, mainly directed to the modulation of inflammation and of the immune system [3]. As recently outlined [2], as genomic-based studies, proteomics contributed to evidence the role of inflammation in AC and therefore to the design of targeted therapies. The efforts, which in the past and recent years were devoted to disclose the molecular features of this tumor, were very recently summarized [3]. Inside these results, the preeminent role of CTNNB1 gene mutation in producing beta catenin accumulation in cluster cells and the subsequent WNT signaling activation was first described. The WNT/beta catenin signaling would have a role in tumor growth and invasion through the paracrine secretion of cytokines and chemokines in the surrounding cells. In addition fascin, an actin-bundling protein, found overexpressed in AC cluster cells, could be also responsible for cancer cells migration and tumor infiltration. AC distinguishes for the proteomic characterization in the intracystic fluid of proteins and peptides involved in inflammation [2] and showed, with respect to other brain tumors, high levels of chemokines, cytokines, and immunosuppressive factors transcripts in both its cyst fluid and solid components [4].
The description of the detailed results obtained by molecular studies on AC is out of the purpose of the present paper. Indeed, herein, we present the original comparison of the proteomic data of the liquid cyst and the solid component of AC accomplished by our group by diverse proteomic strategies in order to investigate their differences and similarities and to discuss them in the context of the current literature.
Proteomic strategies: the top-down and the bottom-up platforms
With the purpose to disclose the molecular events responsible for the occurrence of AC and to understand its locally aggressive behavior, proteomic analysis gave its contribution by investigating either the intracystic fluid or the solid component of AC by top-down and bottom-up platforms based on either LC-MS or bi-dimensional electrophoretic analyses. These strategies gave precious information that complemented genomic and transcriptomic data in the assembling the molecular medley underlining AC pediatric brain tumor onset and progression. Indeed, the protein phenotype of a cell, a tissue, or a biological fluid is not the direct mirror of the expression profile of the genetic construct, because post-translation modifications (PTMs), such as phosphorylation, acetylation, glycosylation, oxidation, or C- and N-terminal truncations, can take place and, moreover, can be modulated under human development and pathological or epigenetic processes.
The top-down strategy, by characterizing the undigested, and therefore, the naturally occurring protein and peptide pattern of a biological matrix, is the strategy of excellence to investigate proteoforms and isoforms and to characterize the PTMs concurring to define deeply and to complete the drawing of the protein phenotype. Moreover, this approach interestingly allows studying the fragmentome, i.e., the intriguing pool of peptides generated in vivo from proteases activity on parent proteins, some of them to date called cryptides [5], which typically exhibit a specific activity different from that of the protein of origin. Many of them have been characterized in pediatric brain tumor tissues, as recently reviewed [5]. In spite of the advantages of this strategy, the complexity of the biological matrix under characterization sometimes limits its application. The sample pretreatment is indeed a critical step to carry out the identification of small proteins and peptides and minor elements present in the sample. Top-down proteomics is in fact limited to the analysis of small proteins (< 25–50 kDa), and therefore, the removal of high molecular weight proteins by accurate sample extraction is strongly recommended.
Differently, the proteome analysis after enzymatic or chemical digestion, under the name of bottom-up or shotgun proteomics, the most classical and widely applied approach, gives diverse information, being able to identify protein in a wide mass range, including high molecular weight proteins, and providing large set of data suitable for bioinformatics elaboration, gene ontology analysis, and pathway classifications. Conversely, following this approach, the disclosure of naturally occurring bioactive peptides or peptides fragments is tricky. Nonetheless, with their pros and cons, these approaches complement each other and, if both applied, constitute a valuable integrated analytical platform for a comprehensive proteomic characterization of pathological specimens.
AC intracystic fluid and solid component: comparing the proteomic profiles
To better understand the molecular mechanisms involved in AC onset and development, both the characterization of the tumor tissue and the fluid content of the cyst are of relevance. In fact, the liquid cyst, often associated to this tumor and possibly compressing the surrounding eloquent vascular and neural structures because of the large size of the cyst, may be responsible for important co-morbidities critically upsetting quality of life of affected children. Hence, efforts were initially devoted to characterize the proteomic content of the intracystic fluid to elucidate the mechanism of formation and growth of the cyst. The results were therefore integrated with the data obtained on AC solid component for a comprehensive characterization of the tumor.
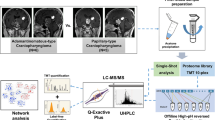
Our group investigated the protein composition of AC intracystic fluid [6,7,8] and AC solid component [9] by the application of a top-down/bottom-up LC-MS integrated proteomic platform following diverse pretreatments according to the analytical approach used (Fig. 1). A recent review summarizes our results on proteomic investigation of AC intracystic fluid [1].

Proteomic strategies and experimental workflow applied to adamantinomatous craniopharyngioma intracystic fluid and solid component
The proteomic analysis of AC intracystic fluid required the careful optimization of the pretreatment procedure due to its peculiar composition. With respect to other tumor intracystic fluids, it is characterized by a yellow-brown liquid feature containing calcium flecks, bone residues, cholesterol granule solid components, and a noticeable amount of lipids, especially cholesterol, resembling a “motor oil fluid” [10]. The top-down proteomic analysis was applied to the soluble fraction obtained after sample acidification, while different pretreatment procedures were applied for protein digestion by bottom-up approach, including untreated intracystic fluid or treated for high abundant protein depletion and lipid removal. A different procedure was applied to the solid component of AC. After tissue homogenization, different aliquots of the resulting supernatant were submitted to top-down or bottom-up analyses after a protein precipitation step. For top-down analyses, the protein pellet was re-dissolved in acidic solution and the supernatant was analyzed. For bottom-up analyses, the protein pellet was re-dissolved and processed for enzymatic digestion following the protocol reported in [9].
Behind the diverse proteomic strategies, the key discovery was the characterization, in AC intracystic fluid, of proteins and peptides involved in inflammation. In particular, the analysis of AC intracystic fluid by top-down approach allowed the identification of alpha-defensins 1–4 and beta-thymosins as the most interesting peptide components, together with other proteins involved in inflammation, mineralization processes, and lipid transport and metabolism, i.e., alpha-2-HS-glycoprotein, alpha1-antichymotrypsin, and apolipoproteins [8]. Alpha-defensins 1–4 (HNP1–4), antimicrobial peptides characterized by multiple disulfide bonds in their structure, are mainly expressed by neutrophils and are released in response to inflammation [11, 12]. The mechanism by which HNP-1 exerts its anti-inflammatory properties has been recently elucidated [13]. The intracystic injection treatment with interferon-α produced a decrease of alpha-defensins peptides and a reduction of the cyst dimension, evidencing a possible correlation between cyst development and growth and innate immunity response and inflammation. Recent transcriptomic studies disclosed elevated levels of inflammatory mediators, i.e., chemokines and cytokines (IL-1, IL-8, CXCL1) in both the cystic fluid and the solid component of AC with respect to other brain tumors [4], thus confirming the important role of inflammation in the pathogenesis of this tumor and its potential targeting for therapeutics. In this study, it was for the first time evidenced the presence in AC of elevated levels of IL-10 and IDO-1 immunosuppressive factors with respect to other tumors.
Beta-thymosins are about 5 kDa peptides found in many cells and exerting several biological activities [14, 15]. In addition to a potential anti-inflammatory role of thymosin beta-4 in AC, as recently reported specifically in the brain [16], this peptide could be involved in tumor growth and infiltration. Thymosin beta-4 is in fact the main sequestering agent of monomeric actin and, through regulation of its polymerization, plays a role in cell differentiation, migration, in the morpho- and organogenesis processes and cytoskeletal organization [17, 18]. Thymosin beta-4 could act as promoting agent of tumor development and invasion via E-cadherin downregulation and activation of beta-catenin signaling [19, 20]. On the other hand, thymosin beta-4 was reported to exert its anti-inflammatory action by the downregulation of pro-inflammatory cytokines, chemokines, and matrix metalloproteinases and through its action on NFkB. Moreover, the anti-inflammatory effect of the peptide was also possibly ascribed to its capacity to upregulate IL-10 [17]. The characterization of thymosin beta-4 in AC and the high levels of IL-10 in the tumor tissue [4] could be therefore an interesting correlation.
Thymosin beta-4 and beta-10 are very similar peptides, both involved in monomeric acting binding and exerting similar functions [21, 22]. These peptides have been characterized both in the intracystic fluid and in the solid component of AC following the same proteomic approach, the top-down strategy.
It is not to be excluded that the characterization of thymosin beta-4 in AC could be in some way linked to the high levels of fascin protein recognized in the tumor in addition to beta catenin accumulation [3]. Fascin, an actin-bundling protein, was ascribed to have a role, in connection with WNT signaling, in promoting cancer cells migration and invasion in adjacent structures. Actin could be the unquestionable link between thymosin beta 4 and fascin, the latter possibly concurring in tumor progression.
The role of beta-thymosin peptides in tumors has been long time discussed [23], and their role as potential biomarkers or therapeutic targets for the management of cancers has been recently proposed [24,25,26]. Thymosin beta-4 has found to promote tumor metastasis and angiogenesis [27] and to be overexpressed in several cancers [28,29,30]. The levels of thymosin beta-10, however, mainly associated to malignant tumors [25, 31], varied depending on the tumor type and its role is still largely unclear. Recently, the mechanisms of action of beta-thymosins in some type of tumors have been outlined [32,33,34]. It was in fact reported that thymosin beta 10 promotes breast cancer invasion and proliferation through the activation of AKT/FOXO signaling [32]. Particularly, in relation to brain tumors, thymosin beta 4 was recently established to promote the invasiveness of glioblastoma, because the gene silencing reduced the tumor progression [33]. Very recently, in a study on human cervical cancer cells, thymosin beta 4 overexpression was associated to increased formation of primary cilia, involved with their assembling/disassembling processes in tumorigenesis and tumor invasion, through the capability of the peptide to interact with and regulate the NPHP3 ciliary protein [34].
Previous studies by our group recognized thymosin beta 4 overexpression in posterior fossa medulloblastoma with respect to pilocytic astrocytoma tissue pools [35] and the exclusive detection of thymosin beta 10 in the most aggressive histotype. In a proteomic study on posterior fossa ependymoma, thymosin beta 4 was statistically significant overexpressed in the supratentorial tumor location [36].
Both alpha-defensins and beta-thymosin peptides have been characterized in the solid component of AC as well using a top-down strategy based on LC-MS analysis of the acid-soluble protein fraction of tissue homogenates.
The characterization of alpha-defensins also in AC tissue is a further confirmation of the role of inflammation and innate immunity in the pathology.
Concerning beta-thymosins, it is interesting to underline that in tumor tissue, the two peptides showed the presence of different proteoforms. Particularly, the oxidized form of thymosin beta 4 and the des-IS-truncated form of thymosin beta 10, missing two C-terminal amino acid residues, were co-identified [9]. It is noteworthy to point out that when secreted, i.e., in AC intracystic fluid, both the beta-thymosins appeared exclusively in their intact form while, in tissues, various PTMs, such as oxidation, fragmentation, or C-terminal truncation, were detected with diverse ratios depending on tumor type. C-terminal-truncated forms of both beta-thymosin peptides have been in fact previously characterized in the brain tissue of Alzheimer’s disease model mice [37] and in tumor tissue pools of posterior fossa pediatric brain tumors [35]. Particularly, in medulloblastoma, marked by thymosin beta 10 as above described, the thymosin beta 10 des-IS–truncated form was particularly abundant and was prevalent over the entire form [35].
The process of formation and, particularly, the biological significance of the truncated proteoforms of beta-thymosin peptides are still to be clarified. However, their absence in fluids of secretion could be indicative of a different intracellular and extracellular role of these peptides to be deeply investigated and considered in the perspective of potential targeted therapies, especially in cancer. The presence of beta-thymosin peptides in the fluid and the solid component of AC, on the light of their reported role in cancer above described, could be correlated to the aggressive behavior of this, still classified benign, tumor.
The absence of C-terminal truncation of thymosin beta 4 in AC tissue and the presence of its sulfoxide form at Met6 residue are other interesting findings. Thymosin beta 4 sulfoxide has been identified in human saliva, colon cancer, and humor vitreous [38]. Thymosin beta 4 sulfoxide has the same affinity as thymosin beta 4 for actin monomers. However, it has no inhibitory action but only an effect of delay on actin polymerization [39]. The formation of thymosin beta 4 sulfoxide seems to occur naturally, and it is probably part of the fine system of regulation of filament formation. The oxidation at methionine site was reported to interestingly enhance the extracellular signaling of thymosin beta 4 peptide in exerting its anti-inflammatory action [40]. Moreover, recently, the capacity of thymosin beta 4 sulfoxide of immune cell scattering has been specifically evidenced [41]. In fact, at the wound site, thymosin beta 4 sulfoxide, formed by hydrogen peroxide, would act as a multifunction dispersing agent of the immune cells, preventing chronic inflammation, and controlling tissue repair. It is then surprising that thymosin beta 4 sulfoxide was undetected in AC intracystic fluid while instead detected in its solid component. Probably, in the cystic fluid, the conditions are not proper for the formation of the sulfoxide derivative from thymosin beta 4, maybe due to the absence of hydrogen peroxide or to a different function of the thymosin beta 4 peptide.
Interestingly, thymosin beta 4 sulfoxide is, in contrast to the parent peptide, a potent inhibitor of mouse paw carrageenin-induced edema, thus exhibiting a potent anti-inflammatory activity [40]. It could be speculated that the absence of detectable levels of thymosin beta 4 sulfoxide in AC intracystic fluid might be somehow involved in an uncontrolled mechanism of cyst growth and inflammation, instead probably promoted by thymosin beta 4 through its regulation capability of actin polymerization and marked by alpha-defensins peptides.
In analogy to what has been observed for thymosin beta 10, while AC intracystic fluid was characterized by the detection of only the entire form of ubiquitin monomer (MW 8560.62, [M + H]+ monoisotopic) [8], the solid component of AC exhibited the presence of its des-GG- and des-RGG-truncated forms missing two or three C-terminal amino acid residues, respectively [9]. As for the beta-thymosin-truncated proteoforms, the biological significance of the C-terminal-truncated ubiquitin monomers is still under investigation, although they were often associated to malignancies and are indicative of the possible alteration of the processes of ubiquitination. C-terminal-truncated ubiquitin, as well as truncated beta-thymosins, has been described in melanoma patients in association with low survival [42]. Similarly to des-IS thymosin beta 10, the des-GG ubiquitin was overexpressed and was more abundant than the unmodified ubiquitin in the most aggressive medulloblastoma with respect to pilocytic astrocytoma [35]. C-terminal-truncated ubiquitin was reported to have a distinguished profile in childhood leukemia [43] and breast cancer [44]. Recently, the des-GG ubiquitin was reported to be a potential biomarker of risk of cancer development in cirrhotic patients, due to its recognized accumulation during the process of transition of cirrhosis to hepatocellular carcinoma, ascribed to the action of kallikrein-related peptidase 6 on monomeric ubiquitin substrates [45].
In the solid component of AC, the levels of ubiquitin monomer and its des-GG-truncated forms were similar. The ubiquitin-truncated proteoforms are possibly linked to the activity of the ubiquitin carboxyl-terminal hydrolase isoenzyme L1 (UCHL1), one of the most represented proteins characterized by the bottom-up approach [9].
The discovery of the C-terminal-truncated proteoforms of ubiquitin and beta-thymosins in AC solid component could therefore be a clue of the aggressive behavior of the tumor, since these forms have been generally found in high-grade tumors. Table 1 summarizes the diverse proteoforms of beta-thymosin peptides and ubiquitin identified in AC.
The AC solid component and intracystic fluid further shared the identification of specific hemoglobin fragments of the alpha globin chain, namely the fragments 2-33 and 2-34. These fragments of molecular mass of 3326.70 and 3473.77 Da ([M + H]+, monoisotopic), respectively, belong to a group of tissue-specific cryptides of hemoglobin origin previously identified in cerebrospinal fluid in relation to pediatric brain tumors of posterior fossa [46]. However, their biological significance has not been fully elucidated yet [5].
The results of top-down analysis of the solid component of AC revealed a wider number of identifications with respect to intracystic fluid, in accordance with the type of biological matrices and the diverse sample pretreatment procedure applied. It is noteworthy to underline that some peptides identified by top-down proteomic analysis of the solid component were also identified in the intracystic fluid following the bottom-up approach. It is for instance the case of the fragment 387-423 of alpha-1-antichymotrypsin, corresponding to the C-terminal part of the protein and interestingly reported to be involved in the acute response to inflammation, as identified as candidate biomarker of glioblastoma in cerebrospinal fluid [47] and of acute renal allograft rejection [48]. This peptide was suggested as produced in vivo by the action of cathepsin D [49]. Although identified by bottom-up approach, this peptide seemed naturally occurring in the cystic fluid rather than produced by the tryptic digestion of the parent protein.
All proteins identified in AC intracystic fluid by bottom-up approach, namely alpha-2-HS-glycoprotein (AHSG), apolipoproteins C-I, A-I, A-II and J (Clusterin), vitamin D–binding protein (DBP), and alpha-1-acid glycoprotein 1 (AGP1), have been likewise identified in the tumor solid component. They demonstrate the strict correlation of the cyst to the solid tumor part, not only on morphological but also on molecular basis. These proteins are in accordance with the characteristics of the cyst and the behavior of the tumor, since they are involved in mineralization processes (particularly of calcium and phosphate), lipid transport and metabolism, and inflammatory response [1].
Conclusions
It is worth outlining that the liquid cyst and the solid components of AC share many protein and peptide elements involved in inflammation, oncogenesis, and calcium metabolism, with a clear indication of their similar molecular features and of common molecular events occurring during the tumor pathogenesis. The proteomic approaches can be considered a successful integrated analytical platform for the comprehensive characterization of AC tumor, complementing the data of its cystic fluid with its solid part. The AC intracystic fluid showed a poorer proteomic profile than the relative solid component. However, the diverse pretreatment procedures of the specimens and the different biological matrix composition, i.e., the high lipid content, have to be taken into account. Nonetheless, the results allowed to outline interesting discussion on the protein and peptides commonly identified in AC intracystic fluid and solid component, hypothesizing their possible role in the disease, either as marker of inflammation, i.e., alpha-defensins, or involved in cell migration and motility, i.e., beta-thymosin peptides, or involved in tumor growth and infiltration as well as in anti-inflammatory processes.
An interesting aspect was the identification of proteoforms of thymosins and ubiquitin peptides in the AC solid component, instead found only in the intact form in the cystic fluid. The top-down proteomic approach originally disclosed in AC solid component the C-terminal-truncated forms of thymosin beta 10 and ubiquitin, missing the last two or three amino acid residues, generally associated to malignancies and therefore representing a possible indication of the aggressive behavior of AC. On the other hand, the finding of the oxidized form of thymosin beta 4 could have instead an anti-inflammatory protective role. The absence of these proteoforms in the intracystic fluid, where only the entire forms of beta-thymosin peptides were detected, could suggest a different intra- and extracellular mechanisms of action of the peptides and/or of their different modulation through various PTMs. Overall, the comparison of the proteomic profiles of the solid component and of the intracystic fluid of AC disclosed many similarities from a proteomic point of view, further confirming the role of inflammation in the disease pathogenesis and giving new hints to deeply investigate the molecular mechanisms involved in the aggressive behavior and high recurrence of this tumor.
References
Massimi L, Martelli C, Caldarelli M, Castagnola M, Desiderio C (2017) Proteomics in pediatric cystic craniopharyngioma. Brain Pathol 27:370–376
Hengartner AC, Prince E, Vijmasi T Hankinson TC (2020) Adamantinomatous craniopharyngioma: moving toward targeted therapies. Neurosurg Focus 48:E7
Whelan R, Prince E, Gilani A, Hankinson T (2020) The inflammatory milieu of adamantinomatous craniopharyngioma and its implications for treatment. J Clin Med 9:519
Donson AM, Apps J, Griesinger AM, Amani V, Witt DA, Anderson RCE, Niazi TN, Grant G, Souweidane M, Johnston JM, Jackson EM, Kleinschmidt-DeMasters B, Handler MH, Tan AC, Gore L, Virasami A, Gonzalez-Meljem JM, Jacques TS, Martinez-Barbera JP, Foreman NK, Hankinson TC, Advancing Treatment for Pediatric Craniopharyngioma Consortium (2017) Molecular analyses reveal inflammatory mediators in the solid component and cyst fluid of human adamantinomatous craniopharyngioma. J Neuropathol Exp Neurol 76:779–788
Iavarone F, Desiderio C, Vitali A, Messana I, Martelli C, Castagnola M, Cabras T (2018) Cryptides: latent peptides everywhere. Crit Rev Biochem Mol Biol 53:246–263
Pettorini BL, Inzitari R, Massimi L, Tamburrini G, Caldarelli M, Fanali C, Cabras T, Messana I, Castagnola M, Di Rocco C (2010) The role of inflammation in the genesis of the cystic component of craniopharyngiomas. Childs Nerv Syst 26:1779–1784
Desiderio C, Martelli C, Rossetti DV, Di Rocco C, D’Angelo L, Caldarelli M, Tamburrini G, Iavarone F, Castagnola M, Messana I, Cabras T, Faa G (2013) Identification of thymosins β4 and β 10 in paediatric craniopharyngioma cystic fluid. Childs Nerv Syst 29:951–960
Martelli C, Iavarone F, Vincenzoni F, Rossetti DV, D’Angelo L, Tamburrini G, Caldarelli M, Di Rocco C, Messana I, Castagnola M, Desiderio C (2014) Proteomic characterization of pediatric craniopharyngioma intracystic fluid by LC-MS top-down/bottom-up integrated approaches. Electrophoresis 35:2172–2183
Martelli C, Serra R, Inserra I, Rossetti DV, Iavarone F, Vincenzoni F, Castagnola M, Urbani A, Tamburrini G, Caldarelli M, Massimi L, Desiderio C (2019) Investigating the protein signature of adamantinomatous craniopharyngioma pediatric brain tumor tissue: towards the comprehension of its aggressive behavior. Dis Markers 2;2019:3609789 https://doi.org/10.1155/2019/3609789
Fernandez-Miranda JC, Gardner PA, Snyderman CH, Devaney KO, Strojan P, Suárez C, Genden EM, Rinaldo A, Ferlito A (2012) Craniopharyngioma: a pathologic, clinical, and surgical review. Head Neck 34:1036–1044
Maneerat Y, Prasongsukarn K, Benjathummarak S, Dechkhajorn W, Chaisri U (2016) Increased alpha-defensin expression is associated with risk of coronary heart disease: a feasible predictive inflammatory biomarker of coronary heart disease in hyperlipidemia patients. Lipids Health Dis 15:117
Ramasundara M, Leach ST, Lemberg DA, Day AS (2009) Defensins and inflammation: the role of defensins in inflammatory bowel disease. J Gastroenterol Hepatol 24:202–208
Brook M, Tomlinson GH, Miles K, Smith RW, Rossi AG, Hiemstra PS, van ‘t Wout EF, Dean JL, Gray NK, Lu W, Gray M (2016) Neutrophil-derived alpha defensins control inflammation by inhibiting macrophage mRNA translation. Proc Natl Acad Sci U S A 113:4350–4355
Hannappel E (2007) Beta-thymosins. Ann N Y Acad Sci 1112:21–37
Goldstein AL, Badamchian M (2004) Thymosins: chemistry and biological properties in health and disease. Expert Opin Biol Ther 4:559–573
Pardon MC (2018) Anti-inflammatory potential of thymosin β4 in the central nervous system: implications for progressive neurodegenerative diseases. Expert Opin Biol Ther 18(sup1):165–169
Crockford D, Turjman N, Allan C, Angel J (2010) Thymosin beta4: structure, function, and biological properties supporting current and future clinical applications. Ann N Y Acad Sci 1194:179–189
Sanders MC, Goldstein AL, Wang YL (1992) Thymosin beta 4 (Fx peptide) is a potent regulator of actin polymerization in living cells. Proc Natl Acad Sci U S A 89:4678–4682
Qiu P, Wheater MK, Qiu Y, Sosne G (2011) Thymosin beta4 inhibits TNF-alpha-induced NF-kappaB activation, IL-8 expression, and the sensitizing effects by its partners PINCH-1 and ILK. FASEB J 25:1815–1826
Nemolato S, Restivo A, Cabras T, Coni P, Zorcolo L, Orrù G, Fanari M, Cau F, Gerosa C, Fanni D, Messana I, Castagnola M, Casula G, Faa G (2012) Thymosin β 4 in colorectal cancer is localized predominantly at the invasion front in tumor cells undergoing epithelial mesenchymal transition. Cancer Biol Ther 13:191–197
Yu FX, Lin SC, Morrison-Bogorad M, Yin HL (1994) Effects of thymosin beta 4 and thymosin beta 10 on actin structures in living cells. Cell Motil Cytoskeleton 27:13–25
Kuzan A (2016) Thymosin β as an actin-binding protein with a variety of functions. Adv Clin Exp Med 25:1331–1336
Chen C, Li M, Yang H, Chai H, Fisher W, Yao Q (2005) Roles of thymosins in cancers and other organ systems. World J Surg 29:264–270
Goldstein AL (2003) Thymosin beta4: a new molecular target for antitumor strategies. J Natl Cancer Inst 95:1646–1647
Sribenja S, Li M, Wongkham S, Wongkham C, Yao Q, Chen C (2009) Advances in thymosin beta10 research: differential expression, molecular mechanisms, and clinical implications in cancer and other conditions. Cancer Investig 27:1016–1022
Xiao Y, Chen Y, Wen J, Yan W, Zhou K, Cai W (2012) Thymosin β4: a potential molecular target for tumor therapy. Crit Rev Eukaryot Gene Expr 22:109–116
Cha HJ, Jeong MJ, Kleinman HK (2003) Role of thymosin beta4 in tumor metastasis and angiogenesis. J Natl Cancer Inst 95:1674–1680
Zhang Y, Feurino LW, Zhai Q, Wang H, Fisher WE, Chen C, Yao Q, Li M (2008) Thymosin Beta 4 is overexpressed in human pancreatic cancer cells and stimulates proinflammatory cytokine secretion and JNK activation. Cancer Biol Ther 7:419–423
Oh SY, Song JH, Gil JE, Kim JH, Yeom YI, Moon EY (2006) ERK activation by thymosin-beta-4 (TB4) overexpression induces paclitaxel-resistance. Exp Cell Res 312:1651–1657
Gemoll T, Strohkamp S, Schillo K, Thorns C, Habermann JK (2015) MALDI-imaging reveals thymosin beta-4 as an independent prognostic marker for colorectal cancer. Oncotarget 6:43869–43880
Theunissen W, Fanni D, Nemolato S, Di Felice E, Cabras T, Gerosa C, Van Eyken P, Messana I, Castagnola M, Faa G (2014) Thymosin beta 4 and thymosin beta 10 expression in hepatocellular carcinoma. Eur J Histochem 58:2242
Zhang X, Ren D, Guo L, Wang L, Wu S, Lin C, Ye L, Zhu J, Li J, Song L, Lin H, He Z (2017) Thymosin beta 10 is a key regulator of tumorigenesis and metastasis and a novel serum marker in breast cancer. Breast Cancer Res 19:15
Wirsching HG, Krishnan S, Florea AM, Frei K, Krayenbühl N, Hasenbach K, Reifenberger G, Weller M, Tabatabai G (2014) Thymosin β 4 gene silencing decreases stemness and invasiveness in glioblastoma. Brain 137(Pt 2):433–448
Lee JW, Kim HS, Moon EY (2019) Thymosin β-4 is a novel regulator for primary cilium formation by nephronophthisis 3 in HeLa human cervical cancer cells. Sci Rep 9:6849
Martelli C, Iavarone F, D’Angelo L, Arba M, Vincenzoni F, Inserra I, Delfino D, Rossetti DV, Caretto M, Massimi L, Tamburrini G, Di Rocco C, Caldarelli M, Messana I, Castagnola M, Sanna MT, Desiderio C (2015) Integrated proteomic platforms for the comparative characterization of medulloblastoma and pilocytic astrocytoma pediatric brain tumors: a preliminary study. Mol BioSyst 11:1668–1683
Rossetti DV, Massimi L, Martelli C, Vincenzoni F, Di Silvestre S, Scorpio G, Tamburrini G, Caldarelli M, Urbani A, Desiderio C (2020) Ependymoma pediatric brain tumor protein fingerprinting by integrated mass spectrometry platforms: a pilot investigation. Cancers (Basel) 12:674
Delfino D, Rossetti DV, Martelli C, Inserra I, Vincenzoni F, Castagnola M, Urbani A, Scarpa S, Fuso A, Cavallaro RA, Desiderio C (2019) Exploring the brain tissue proteome of TgCRND8 Alzheimer’s disease model mice under B vitamin deficient diet induced hyperhomocysteinemia by LC-MS top-down platform. J Chromatogr B Analyt Technol Biomed Life Sci 1124:165–172
Cabras T, Iavarone F, Martelli C, Delfino D, Rossetti DV, Inserra I, Manconi B, Desiderio C, Messana I, Hannappel E, Faa G, Castagnola M (2015) High-resolution mass spectrometry for thymosins detection and characterization. Expert Opin Biol Ther 15:S191–S201
Heintz D, Reichert A, Mihelic-Rapp M, Stoeva S, Voelter W, Faulstich H (1994) The sulfoxide of thymosin beta 4 almost lacks the polymerization-inhibiting capacity for actin. Eur J Biochem 223:345–350
Young JD, Lawrence AJ, MacLean AG, Leung BP, McInnes IB, Canas B, Pappin DJ, Stevenson RD (1999) Thymosin beta 4 sulfoxide is an anti-inflammatory agent generated by monocytes in the presence of glucocorticoids. Nat Med 5:1424–1427
Evans MA, Smart N, Dubé KN, Bollini S, Clark JE, Evans HG, Taams LS, Richardson R, Lévesque M, Martin P, Mills K, Riegler J, Price AN, Lythgoe MF, Riley PR (2013) Thymosin β4-sulfoxide attenuates inflammatory cell infiltration and promotes cardiac wound healing. Nat Commun 4:2081
Hardesty WM, Kelley MC, Mi D, Low RL, Caprioli RM (2011) Protein signatures for survival and recurrence in metastatic melanoma. J Proteome 74:1002–1014
Hegedus CM, Gunn L, Skibola CF, Zhang L, Shiao R, Fu S, Dalmasso EA, Metayer C, Dahl GV, Buffler PA, Smith MT (2005) Proteomic analysis of childhood leukemia. Leukemia 19:1713–1718
Gonçalves A, Charafe-Jauffret E, Bertucci F, Audebert S, Toiron Y, Esterni B, Monville F, Tarpin C, Jacquemier J, Houvenaeghel G, Chabannon C, Extra JM, Viens P, Borg JP, Birnbaum D (2008) Protein profiling of human breast tumor cells identifies novel biomarkers associated with molecular subtypes. Mol Cell Proteomics 7:1420–1433
Laouirem S, Le Faouder J, Alexandrov T, Mestivier D, Léger T, Baudin X, Mebarki M, Paradis V, Camadro JM, Bedossa P (2014) Progression from cirrhosis to cancer is associated with early ubiquitin post-translational modifications: identification of new biomarkers of cirrhosis at risk of malignancy. J Pathol 234:452–463
Desiderio C, D’Angelo L, Rossetti DV, Iavarone F, Giardina B, Castagnola M, Massimi L, Tamburrini G, Di Rocco C (2012) Cerebrospinal fluid top-down proteomics evidenced the potential biomarker role of LVV- and VV-hemorphin-7 in posterior cranial fossa pediatric brain tumors. Proteomics 12:2158–2166
Schuhmann MU, Zucht HD, Nassimi R, Heine G, Schneekloth CG, Stuerenburg HJ, Selle H (2010) Peptide screening of cerebrospinal fluid in patients with glioblastoma multiforme. Eur J Surg Oncol 36:201–207
Ziegler ME, Chen T, LeBlanc JF, Wei X, Gjertson DW, Li KC, Khalighi MA, Lassman CR, Veale JL, Gritsch HA, Reed EF (2011) Apolipoprotein A1 and C-terminal fragment of α-1 antichymotrypsin are candidate plasma biomarkers associated with acute renal allograft rejection. Transplantation 92:388–395
Pimenta DC, Chen VC, Chao J, Juliano MA, Juliano L (2000) Alpha1-antichymotrypsin and kallistatin hydrolysis by human cathepsin D. J Protein Chem 19:411–418
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Desiderio, C., Rossetti, D.V., Castagnola, M. et al. Adamantinomatous craniopharyngioma: advances in proteomic research. Childs Nerv Syst 37, 789–797 (2021). https://doi.org/10.1007/s00381-020-04750-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00381-020-04750-z