
Abstract
A comparative evaluation of arsenic (As)-induced toxicity has been made in selected Western Himalayan ferns belonging to the family Pteridaceae (Pteris vittata L. and P. cretica L.) and Polypodiaceae (Polypodiodes microrhizoma (C. B. Clarke ex Baker) Ching and Lepisorus contortus (H. Chirst) Ching) employing a rapid leaf (pinna) slice test. The exposure of pinna segments to As(V) (Na2HAsO4; 0–1000 µM), led to a concentration-dependent increase in K+ leakage into the incubation medium owing to membrane damage. Strong species-specific differences were evident. Thus, the magnitude of K+ leakage enhancement was much lower in Pteridaceae members (lowest in P. vittata) than in those of Polypodiaceae. The As effects were like those of Cu, a redox active element like As, and H2O2 suggesting them to be mediated by oxidative stress. Involvement of K+ channels in As-induced K+ leakage is unlikely, as no effect of tetraethylammonium chloride (TEA), a K+ channel inhibitor was evident. Furthermore, the activities of antioxidant enzymes, superoxide dismutase (SOD), catalase (CAT) and ascorbate peroxidase (APX) were markedly altered due to As treatment in a species-specific manner pointing to a role of antioxidant defence in differential response and thereby tolerance of the concerned ferns to As. Findings have implications for phytoremediation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Arsenic (As) contamination of the environment, owing to natural as well as anthropogenic release, is a matter of serious concern (Bhattacharya et al. 2007; Punshon et al. 2017; Kumar et al. 2018; Thakur et al. 2019). Indeed, the As levels in groundwater are well above the safe limits as per the WHO standards in some Asian countries like Bangladesh as well as in USA (Henke 2009). Arsenic persists in the food chain following its uptake and accumulation by diverse crops such as rice that eventually poses a threat to human health. The increased As intake through food or drinking water is associated with incidences of cancer, skin diseases, kidney problems, diabetes and heart ailments (Meliker et al. 2007).
Phytoremediation, employing metal(loid) hyperaccumulator plants, offers a reasonable means of lowering the toxic heavy metal(loid) burden on environment (Pilon-Smits 2005; Krämer 2010). In the context of As toxicity, identification of Chinese brake fern (Pteris vittata) as the first As hyperaccumulator was a breakthrough (Ma et al. 2001). This led to the screening of other fern species for their ability to hyperaccumulate As. Several species e.g., P. cretica, P. longifolia, P. umbrosa, Pityrogramma calomelanos and P. melanocaulon exhibited substantial As accumulation rates in their fronds (Francesconi et al. 2002; Zhao et al. 2002; Claveria et al. 2019) although they did not match the capabilities of P. vittata (Luongo and Ma 2005). In contrast, several other species e.g., Asplenium nidus, Davillia canarensis, Polypodium aureum, and Polystichum tsus-simense did not have such ability (Zhao et al. 2002). P. vittata represents an attractive model system for getting insight into the mechanisms of As tolerance (Ma et al. 2001). Despite the identification and availability of a large number of metal hyperaccumulators, implementation of phytoremediation in the field remains limited (Gerhardt et al. 2017). Interestingly, intercropping of P. vittata was reported to reduce As accumulation in certain cash crops including maize (Ma et al. 2018) and Morus alba (Wan and Lei 2018).
Arsenic is taken up by P. vittata as As(V), the predominant form in aerobic environments, through specific inorganic phosphate (Pi) uptake systems in competition with Pi (Meharg and Hartley-Whitaker 2002; Tu and Ma 2003). Subsequently, As(V) is converted to As(III) in the roots by arsenate reductases (Ellis et al. 2006) and is accumulated as the major As form in the fronds (Bleeker et al. 2006). In contrast, the uptake of As(III), abundant in reducing environments (pH < 8) such as in the paddy fields occurs via nodulin26-like intrinsic proteins (NIPs) (Ma et al. 2008; Lindsay and Maathuis 2017). PvTIP4;1, an aquaporin localized to plasma membrane, is involved in As(III) uptake in P. vittata (He et al. 2016). Vacuolar compartmentalization is an important component of cellular detoxification of As (Sharma et al. 2016). A tonoplast localized As(III) transporter, PvACR3, has been reported in P. vitatta (Indriolo et al. 2010). As(III) is complexed by phytochelatin (PC) and As(III)-PC complex is transported into the vacuole via two ABC transporters, AtABCC1 and AtABCC2, in Arabidopsis thaliana (Zhao et al. 2003; Song et al. 2010). Recently, a novel mechanism of arsenic detoxification in P. vittata has been reported. Here, the glyceraldehyde 3-phosphate dehydrogenase PvGAPC1 converts arsenate into 1-arseno-3-phosphoglycerate that is transported by PvOCT4 into the As metabolizing vesicles where the released As(V) is reduced by PvGSTF1 to As(III) that is stored in the vacuoles (Cai et al. 2019). Apparently, As hyperaccumulation is linked to the mechanisms of tolerance. The toxicity due to metal(loid)s including As could inter alia be ascribed to the cellular redox imbalance; conversely, the antioxidant defence contributes to As tolerance (Sharma and Dietz 2009). Arsenic altered activities of antioxidant enzymes like superoxide dismutase (SOD) and catalase (CAT) in P. vittata and other ferns (Srivastava et al. 2005). Also, the biological membranes are involved in determining the tolerance to metal(loids) through selective uptake and efflux e.g. in Agrostis tenuis in response to Cu and Zn (Wainwright and Woolhouse 1977) and Mimulus guttatus in response to Cu (Strange and Macnair 1991).
The As hyperaccumulation, associated with elevated As tolerance ability in fronds, is a constitutive trait in Pteris vittata. Nevertheless, As uptake and accumulation rates vary among P. vittata populations originating from distinct geographical locations (Srivastava et al. 2010). P. vittata populations differed in As decreasing efficiency when grown as intercrops with Morus alba (Wan and Lei 2018). Such ecotypic variations likely occur in case of other species of Pteridaceae and related families that might influence the response to arsenic as well as toxic heavy metals (HMs). To exploit this genetic and functional potential, it is of significance to screen the ferns for species- as well as population-specific differences in tolerance to metal(loids). In view of the cross tolerance to metals with common toxicity pathways (Schat and Vooijs 1997), it is interesting to examine the responses of As-hyperaccumulator species to toxic HMs. For example, P. melanocaulon, besides being an As hyperaccumulator, also accumulates and tolerates high concentrations of Cu (Claveria et al. 2019). In the present study, we compared the As-induced toxicity in Western Himalayan populations of P. vittata and P. cretica (Pteridaceae) with that in Polypodiodes microrhizoma and Lepisorus contortus (Polypodiaceae) in terms of membrane integrity and antioxidant defence parameters. Besides, in order to get an idea whether As-imposed toxic effects were a consequence of cellular redox imbalance, they were compared with those of Cu, a redox active metal, and H2O2, a strong oxidant.
Materials and Methods
Plant Source
Pteris vittata L. and Pteris cretica L. (Pteridaceae) and Polypodiodes microrhizhoma (C. B. Clarke ex Baker) Ching and Lepisorus contortus (H. Christ) Ching (Polypodiaceae) were collected from the wild populations in Western Himalayan region; the first from Hamirpur (Himachal Pradesh; 700 m asl) and all others from Shimla hills (Himachal Pradesh; 2300 m asl). The fronds were used in the described experiments.
Determination of the Effects of As, Cu and H2O2 on Membrane Integrity
Membrane integrity of the selected ferns was assessed in a short-term pinna segment assay adopted from a rapid leaf slice test introduced by Cho et al. (2003) for comparing the tolerance of Arabidopsis halleri and A. thaliana to certain HMs. The pinnae from uniform fern fronds were sliced into 1–2 mm wide segments (~ 100 mg), that were vacuum-infiltrated for 5 min with solutions containing As (Na2HAsO4; 0–1000 µM), Cu (50 µM; CuSO4) or H2O2 (50 mM) and incubated in glass Petri plates at 25 °C under constant illumination (PAR; 40 µmol photons m−2 s−1). K+-leakage into the incubation medium and activities of antioxidant enzymes namely, catalase, ascorbate peroxidase and superoxide dismutase in tissue extracts were measured after the stated durations.
Determination of the Effects of Phosphate and TEA on As-Induced K+ Leakage
The effects of simultaneously applied Pi, a structural analogue of As, and tetraethylammonium chloride (TEA), a K+ channel blocker, were examined on As-induced K+ leakage from pinna segments of P. microrhizoma. The pinna segments were vacuum-infiltrated with As (250, 500 µM), NH4H2PO4 (100, 500 and 1000 µM;) and TEA (10 mM) individually or in the stated combinations. K+-leakage was assessed after 18 h of incubation.
Determination of K+ Leakage
K+ content in the pinna segment incubation medium was estimated after 3, 6 and 18 h of treatment using a flame photometer (Corning 400, UK) calibrated using KCl (0–20 ppm).
Antioxidant Enzyme Assays
The fern pinna segments (~ 100 mg) were homogenised with 100 mM HEPES–NaOH buffer (pH 7.6, supplemented with 5 mM ascorbate) and the obtained homogenate centrifuged at 8000×g for 5 min at 4 °C. The supernatant was used for catalase (CAT) and ascorbate peroxidase (APX) activity measurement. CAT activity was assayed polarographically using a Clark-type O2 electrode (Hansatech, UK), calibrated using the signal difference between air- saturated water and water deprived of O2 using sodium dithionite. The reaction mixture contained 890 µl HEPES–NaOH buffer (100 mM, pH 7.6), 10 µl of enzyme extract and 100 µl H2O2 (100 mM). The enzyme activity was calculated from the slopes recorded on chart paper using the value of dissolved O2 at 25ºC (Goldstein 1968; Sharma et al. 2004). APX activity was assayed according to Hossain and Asada (1984). The reaction mixture contained 50 µl ascorbate (5 mM), 1 ml 50 mM HEPES–NaOH buffer (pH 7.6), 50 µl enzyme extract and 100 µl H2O2 (3 mM). Following the addition of H2O2, change in absorbance was monitored at 290 nm and enzyme activity determined by using the extinction coefficient 2.8 mM−1 cm−1 for ascorbate. Superoxide dismutase (SOD) activity was measured using method modified after Beauchamp and Fridovich (1971). Extraction buffer used was 100 mM K-phosphate (pH 7.0). The reaction mixture contained 680–695 µl assay buffer (100 mM K-phosphate, pH 7.8), 100 µl riboflavin (130 µM), 100 µl methionine (13 mM), 100 µl nitro-blue tetrazolium chloride (NBT, 1.26 mM) and 5–20 µl enzyme extract. The reaction mixture was illuminated for 10 min. and the absorbance read at 560 nm. A 50% reduction in absorbance was taken as one-unit activity. Specific activity for all the antioxidant enzymes was calculated from the protein content determined with the Bradford reagent (Bradford 1976).
Tissue Staining for Superoxide (O2 −·) and H2O2 Accumulation
In order to ascertain the role of ROS in As-induced toxicity, accumulation of O2−· and H2O2 in As treated pinna segments was monitored by nitro blue tetrazolium (NBT) and diaminobenzidine (DAB) staining, respectively. The pinna segments treated with As and Cu for 6 h were incubated with NBT (0.05% in PBS buffer, pH 7.8) and DAB (0.05% in 0.1 N HCl) staining solutions overnight in dark. They were then shifted to de-staining solution (ethanol and chloroform in a ratio of 1:5 containing 0.15% trichloroacetic acid) to make stained areas visible through chlorophyll extraction. The stained pinna segments were fixed in PBS-glycerol (1:1). The results are shown in the form of representative images from two biological replicates.
Statistics
Data are presented as arithmetic means ± SE for all parameters except CAT and SOD activities that are presented in box plots with each hollow dot representing a data point (showing different data points available). Data points that are not connected represent outliers. All data were tested for statistical significance using one-way ANOVA followed by Tukey’s test or t-test as described in respective figure legend.
Results
As-Induced K+ Leakage from Pinna Segments
Changes in K+ leakage from fern pinna segments, an indicator of membrane damage, were monitored in response to As(V), Cu(II) and H2O2 treatments (Figs. 1, 2, 3). Species-specific differences were evident which were concentration and treatment time-dependent. Arsenic did not markedly change the K+-leakage from pinna segments of Pteris vittata, P. cretica and Polypodiodes microrhizoma after 3 and 6 h except a marginal increase of 13% after 3 h in P. vittata and 19% after 6 h in P. microrhizoma, at 1000 µM As (Fig. 1). However, As-induced K+ leakage was substantial after 18 h with strong species-specific differences. In P. vittata, an increase of 27% at 1000 µM was observed while in P. cretica this was 52 and 59% at 500 and 1000 µM, respectively. In contrast, P. microrhizoma showed a massive K+ leakage with 3.3-, 4-, and 5-fold increase at 250, 500 and 1000 µM As, respectively. Apparently, P. microrhizoma was much more sensitive to As than the two Pteris species. The control values for K+ contents in incubation medium correspond to basal leakage and cell damage occurred during pinna segment preparation, which also depends on the pinna texture and thickness. As-induced K+-leakage enhancement was paralleled by a similarly increased electrical conductivity of incubation medium (data not shown).

As- and Cu-induced K+-leakage from pinna segments of selected fern species into the incubation medium. The pinna segments (1–2 mm width, ~ 100 mg) of Pteris vittata, P. cretica and Polypodiodes microrhizoma were vacuum-infiltrated for 5 min. with desired treatment solutions and were incubated under continuous light (40 µmol photons m−2 s−1) in the treatment solutions (10 ml) for 3, 6 and 18 h. K+ leakage was measured in the incubation medium by a flame photometer (Corning, 400). Values are means ± SE, n = 6 (all means compared for treatment effects; one-way ANOVA, Tukey’s test p ≤ 0.05)
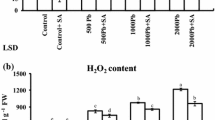
To get an idea whether As effects are a consequence of oxidative stress, they were compared with those of Cu, a redox active element, and H2O2. As in case of As(V), Cu (50 µM) treatment of 18 h enhanced the K+ leakage from pinna segments by a magnitude of 1.56-, 3.01- and 3.9-fold in P. vittata, P. cretica and P. microrhizoma, respectively (Fig. 1). In P. microrhizoma, Cu effect could be observed as early as 3 h after start of the treatment (24% increase). This value increased to 91% after 6 h. Overall, the Cu-sensitivity after 18 h was more alike among different ferns than the As response at any concentration; however, the maximum change was observed for P. microrhizoma. An entirely different response of K+ leakage was seen when fern pinna segments were treated with 50 mM H2O2 for 3 h (Fig. 2A). P. vittata showed a slight increase of 18%, P. cretica lost 21% more K+ to the medium than under control conditions, and only P. microrhizoma showed a substantially increased (77%) K+ leakage. This H2O2-induced pattern after 3 h tentatively reminded of the K+ leakage pattern seen with 50 µM Cu after 18 h, but was poorly related to the As response pattern, apart from the fact that the highest value was denoted for P. microrhizoma. To substantiate the hypothesis that a major difference in membrane susceptibility to As stress exists between Pteridaceae and Polypodiaceae species, another fern from the family Polypodiaceae, namely Lepisorus contortus, was tested. Its pinna segments were exposed to As and Cu for 18 h and to H2O2 for 3 h, and K+ losses recorded (Fig. 2B). The L. contortus pinna segments showed a sharp increase in K+ leakage under As (250, 500 and 1000 µM), Cu (50 µM) and particularly in response to 50 mM H2O2 with 3.13-, 3.73-, 3.04-, 3.85- and 8.73-fold increases, respectively, as compared to controls.

H2O2-induced K+-leakage from the pinna segments of selected fern species. K+-leakage measured in the incubation medium from the pinna segments of P. vittata, P. cretica and Polypodiodes microrhizoma (A) and Lepisorus contortus (B) after 3 h of incubation in H2O2 (50 mM). Also, given is the K+-leakage from the pinna segments of Lepisorus contortus as affected by As and Cu (B) after 18 h of vacuum infiltration. Values are means ± SE, n = 6 (t-test results are given with different letters which represent significantly different means)
As-Induced K+ Leakage as Influenced by Phosphate and K+ Channel Blocker
Since As(V) uptake occurs through phosphate uptake transporters, phosphate could be expected to suppress As-uptake and thereby its toxic effects. However, co-supplementation of phosphate did not affect the magnitude of As-induced K+ leakage in P. microrhizoma (Fig. 3A). The K+ leakage induced by arsenate was similar in the presence and absence of phosphate. Phosphate alone at 500 µM enhanced K+ leakage by 45% as compared to control, but the change was not statistically significant (Fig. 3A). To investigate the involvement of K+ channels in As-induced increase in K+ leakage, the pinna segments of P. microrhizoma were treated with As in presence of tetraethylammonium chloride (TEA), a frequently used K+ channel blocker. The unchanged K+ leakage in the presence of TEA revealed no effect of K+ channel blocker on As (250 µM)-induced K+ leakage (Fig. 3B).

Effect of Pi and TEA (a K+-channel blocker) on As-induced K+-leakage from pinna segments of Polypodiodes microrhizoma. K+-leakage was measured in incubation medium from pinna segments of P. microrhizoma as indicated, A effect of Pi, B effect of tetraethylammonium chloride (TEA). Measurements were done after 18 h of vacuum infiltration. Values are means ± SE, n = 3 (t-test, different letters represent significantly different means)
As-Induced Oxidative Stress
The fern pinna segments were analysed for metal(loid)-induced oxidative stress by in-vivo localization of the reactive oxygen species O2−· and H2O2 after 6 h of treatment. NBT staining showed strong accumulation of O2−· in pinna segments of P. microrhizoma after 6 h of As treatment (Fig. 4). The NBT staining in As-treated P. vittata and P. cretica pinna segments was faint (Fig. 4). Cu also induced higher accumulation of O2−· in pinna segments of P. microrhizoma as compared to those of P. vittata and P. cretica. DAB staining showed As-induced enhanced H2O2 accumulation in P. microrhizoma pinna segments but not in case of P. vittata and P. cretica. Exposure to Cu induced H2O2 accumulation in P. microrhizoma (Fig. 4). The observed species-specific differences in As-induced ROS accumulation prompted us to analyse the activities of antioxidant enzymes namely, CAT, SOD and APX after 3, 6 and 18 h of treatment.

In-vivo localization of superoxide radicals and hydrogen peroxide in the fern pinna segments. Effect of different treatments was observed on the generation of O2−· and H2O2 in differentially treated pinna segments of Pteris vittata, P. cretica and Polypodiodes microrhizoma by NBT and DAB staining, respectively, after 6 h of vacuum infiltration. The images are representative of two independent experiments
CAT Activity
The basal CAT activity in pinna segments was higher in P. cretica and P. vittata than in P. microrhizoma. In response to As treatment, CAT activity in P. vittata tended to increase after 3 and 6 h of treatment although statistical significance was not evident. CAT activity in P. cretica increased after 6 h of As treatment by 67 and 51% at 500 and 1000 µM As, respectively (Fig. 5). However, after 18 h of As treatment, CAT activity decreased in P. vittata compared to control while no change was observed in P. cretica. Arsenic also induced an increase in CAT activity in the pinna segments of P. microrhizoma at 500 and 1000 µM after 3 and 6 h of exposure; however, the activity was significantly lower than that of respective treatments in P. cretica and P. vittata (Fig. 5). For example, after 6 h of As (500 and 1000 µM) exposure, the CAT activity was 2.70-, 2.74-fold and 3.37-, 3.45-fold higher in P. vittata and P. cretica, respectively, as compared to that in P. microrhizoma. After 18 h of treatment, the As-induced increase was only marginal. Due to Cu (50 µM) treatment, CAT activity was inhibited in the pinna segments of P. microrhizoma but remained unchanged in those of P. vittata and P. cretica (Fig. 5).

Catalase activity in differentially treated pinna segments. The activity of catalase was measured in the pinna segments of P. vittata, P. cretica and P. microrhizoma treated with different concentrations of As and Cu after 3, 6 and 18 h of vacuum infiltration. The data are presented in box plots with each hollow dot showing an available data point. In the box plot, centre lines show the medians; box limits indicate the 25th and 75th percentiles as determined by R software; whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles; not connected data points represent outliers, n = 3 (t-test, different letters represent significantly different means; A, B, C… for significant differences between values for different fern species at a specific treatment, while a, b, c… for significant differences among different treatments for a specific fern species). The box plots were generated using the online tool shiny.chemgrid.org/boxplotr
SOD Activity
In response to As treatment, SOD activity increased in P. microrhizoma pinna segments as early as after 3 h although the magnitude of increase did not correspond with As concentrations. The increased activity was maintained until 6 h except at 1000 µM As (Fig. 6). SOD activity was not affected by As after 18 h. In P. vittata, SOD activity was not affected by As after 3 h of treatment; however, it increased in a concentration-dependent manner after 6 h; a 1.9-fold increase was evident at 1000 µM. SOD activity was marginally enhanced by As in P. cretica after 3 h particularly at 250 and 500 µM As. The As-induced activity enhancement was substantial and concentration-dependent after 6 h with a maximal increase of 7.13-fold at 1000 µM. After 18 h, As-induced increase in SOD activity was observed only at 1000 µM both in P. vittata and P. cretica. Interestingly, SOD activity was higher in P. microrhizoma than in P. vittata and P. cretica after 18 h As-exposure. Cu strongly increased the SOD activity at all time points. The magnitude of increase was higher in P. vittata after 6 and 18 h of treatment (Fig. 6).

Superoxide dismutase activity in differentially treated pinna segments. The activity of superoxide dismutase (SOD) was measured in pinna segment extracts of P. vittata, P. cretica and P. microrhizoma treated with As and Cu as indicated after 3, 6 and 18 h of vacuum infiltration. The data are presented in box plots with each hollow dot showing an available data point. In the box plot, centre lines show the medians; box limits indicate the 25th and 75th percentiles as determined by R software; whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles; not connected data points represent outliers. Values are means ± SE, n = 6 (t-test, different letters represent significantly different means; A, B, C… for significant differences between values for different fern species at a specific treatment, while a, b, c… for significant differences among different treatments for a specific fern species)
APX Activity
Constitutive APX activity was higher in pinna segments of P. cretica and P. microrhizoma than that in P. vittata. It changed with the lapse of incubation period particularly in P. vittata and P. microrhizoma with a substantial increase or decrease, respectively (Table 1). In response to As, APX activity increased after 3 h in P. vittata by a factor of 2.39, 7.67 and 6.94 at 250, 500 and 1000 µM As, respectively. A similar pattern with a lower magnitude of increase was observed in P. cretica after 3 h. Thus, the activity was 1.34-, 1.77- and 2.27-fold higher at 250, 500 and 1000 µM As, respectively. Subsequently, however, no definite pattern of change was observed. APX activity tended to increase at higher As concentrations (500 µM in P. cretica and 1000 µM in P. vittata) after 6 h. After 18 h, As suppressed the APX activity in P. cretica at all concentrations and at 1000 µM in P. vittata (Table 1). As suppressed the APX activity in P. microrhizoma after 3 h particularly at 500 µM. The activity was not affected by As after 6 h but was promoted after 18 h by 66, 84 and 50% at 250, 500 and 1000 µM, respectively (Table 1). In response to Cu (50 µM) treatment, APX activity increased in P. vittata after 3 and 6 h by 1.46- and 4.81-fold, respectively and then decreased by 50% after 18 h. In P. cretica, 84% increase in APX activity was observed after 3 h that was followed by a decline of 22 and 50% after 6 and 18 h, respectively. APX activity increased due to Cu after 6 and 18 h in P. microrhizoma by 99 and 40%, respectively (Table 1).
Discussion
Screening for As Tolerance
The initial steps in developing phytoremediation strategies based on hyperaccumulation traits need to ascertain the metal tolerance levels of the species under consideration. In the best case, one should also understand the underlying mechanisms of metal(loid) accumulation. In view of the extraordinary root-to-shoot metal(loid) translocation efficiency of hyperaccumulators the tolerance of shoot organs, in particular the leaves, to the metal(loid)s is of utmost importance. To shorten the process of tolerance screening, Cho et al. (2003) introduced an easy and rapid leaf slice test for deciphering the HM-specific tolerance of Arabidopsis halleri and A. thaliana. On similar lines, in present study, As- tolerance of P. vittata, an As hyperaccumulator, and some related ferns from Western Himalayan region has been evaluated in terms of membrane integrity parameters by direct application of As to the leaf cells using pinna slices. Quick screening through such simple assays could be expected to reveal novel As hyperaccumulator species and the tissue-specific tolerance strategies. As such, a rich fern diversity from the region awaits physiological and ecological scrutiny.
Species Specificity of As-Induced Membrane Damage
Damage to the cell membranes, especially plasma membrane, constitutes one of the primary events in development of HM toxic effects in plants (Janicka-Russak et al. 2008). Conversely, the improved plasma membrane integrity has an adaptive value, in protecting membrane functions and in turn cell integrity. Strong species-specific differences in As-induced K+ leakage, a measure of damage to the membrane permeability barrier, were evident. Arsenic was applied through vacuum infiltration of pinna segments to ensure rapid entry into the intercellular air space and equilibration with the apoplastic fluid. Whereas As induced a massive K+ leakage in case of P. microrhizoma and L. contortus, both members of Polypodiaceae, the magnitude of the same was much lower in case of Pteridaceae ferns P. vittata and P. cretica. The findings clearly point to substantially elevated As tolerance of latter species where the membrane damage was negligible even at 1000 µM As. Indeed, the membrane stability index in P. vittata under As exposure exceeds that in arsenic-sensitive P. ensiformis (Singh et al. 2006). Metal(loid)s cause membrane damage via generation of ROS and subsequent lipid peroxidation or activation of enzymes like lipoxygenases (Sharma and Dietz 2009). In order to get insight into the possibility of mediation of As toxicity by oxidative stress, the As effects were compared with those of Cu, a redox active element like As, on the one hand and H2O2, a strong oxidant, on the other. That both Cu and H2O2 produced species-specific effects resembling qualitatively to those of As strengthens an assumed role of ROS and in turn oxidative stress in the observed As-induced damage to membrane integrity. Cu-induced damage to the root cell plasma membrane was reported in Silene cucubalus (De Vos et al. 1989) and Mimulus guttatus (Strange and Macnair 1991). A stronger metal(loid) tolerance capability of P. vittata and P. cretica as compared to P. microrhizoma and L. contortus might be due to common tolerance strategies for As and Cu, including generally strengthened plasma membrane and/or some specific ecological adaptations. As an example of ecological adaptation, multiple tolerance and co-tolerance to HMs in Silene vulgaris has been reported (Schat and Vooijs 1997). Also, in P. vittata (ecotypes from Kerala, India), Cr tolerance and hyperaccumulation along with that of As was shown (Kalve et al. 2011). There is a need to screen P. vittata as well as other related ferns for tolerance to different HMs.
K+ ions are involved in several cellular functions in plants including activation of enzymes and ribosomes, counterbalancing of ions, turgor adjustment and maintenance of membrane potentials (Maathuis et al. 1997). Indeed, cytosolic K+-contents work as “master switches” in the transition of plant metabolism from the normal to ‘adjusted’ state under abiotic stresses (Shabala and Pottosin 2014). Electrolyte leakage is considered a constituent of plant stress response (Demidchik et al. 2014). The stress mediated K+-efflux provides an adaptive mechanism, e.g. in guard cells in order to initiate stomatal closure. There is a possibility that As, in addition to causing membrane damage via ROS generation, might have stimulated the K+ channels as was suggested in case of Cu treated A. thaliana (Murphy and Taiz 1995). The K+ channels include voltage-dependent, slow activating, outward rectifying K+-selective ‘Shakers’ channels encoded by guard cell outward rectifying K+-channel (GORK), selective K+-outward rectifying channel (SKOR) and annexin genes. Incidentally, exposure to HMs stimulates the generation of ROS such as O2−· and H2O2 (Sharma and Dietz 2009) that in-turn could activate GORK, SKOR and annexins catalysing K+-efflux (Demidchik 2010). Besides, a hypothetical possibility of involvement of voltage-independent, instantaneously activating, cation non-selective ‘Shaker like’ cyclic nucleotide-gated channels (CNGC) and ionotropic glutamate receptors (GLRs) channels is suggested (Demidchik et al. 2014). To examine the possible involvement of K+ channels in the leakage of K+ ions, TEA, a specific K+-channel blocker for voltage-dependent K+-selective ‘Shakers’ (Thompson and Begenisich 2003), was applied simultaneously with As (250 µM) in P. microrhizoma. However, TEA did not alter the As-induced K+ leakage. A slight increase in K+ leakage due to TEA alone might be a result of TEA toxicity. It is pertinent to add that for different categories of voltage-dependent K+-selective ‘Shakers’, the required TEA concentration for effective blocking could vary several fold owing to the differences in the amino acid composition of TEA binding site (Kavanaugh et al. 1991). The likely involvement of other K+ permeable channels in As-dependent K+ leakage has not been examined in this study.
The possible alleviation of As-inflicted membrane damage by the hypothesized competition between Pi and As for uptake pathway was also investigated in P. microrhizoma. Mesophyll cells show a rapid Pi uptake from IWF using proton motive force generated by H+-ATPase (Mimura et al. 1990). The As-induced effect on K+ leakage was not altered by the presence of Pi. However, there is a need to check actual As uptake into the cell. Besides, there is also a possibility of cell wall binding and apoplastic sequestration of As in these ferns as a strategy to avert toxicity as was demonstrated for barley for certain HMs (Brune et al. 1995). Arsenic likely affected the membranes by acting on the apoplastic surface.
Altered Antioxidative Defence System
The stress induced electrolyte leakage is often accompanied by stimulated generation of ROS (Demidchik et al. 2014). Plasmalemma-bound NAD(P)H oxidases as well as cell wall-associated peroxidases are the main O2−· and H2O2 producing apoplastic enzymes induced by HMs (Sagi and Fluhr 2006; Sharma and Dietz 2009). The As-induced increased SOD activity in P. vittata (6 h) and P. cretica (6 and 18 h) pinna segments seems adequate for scavenging of O2−· as is also evident from NBT staining. CAT and peroxidases likely dealt with the subsequently produced H2O2. In accordance, both Pteris species exhibited high constitutive CAT activity vis-à-vis the other fern that was further enhanced due to As. In addition, the higher As concentrations slightly enhanced the APX activity during initial phase (3 h) of treatment. This might be particularly significant for degradation of low concentrations of H2O2 in the stroma, matrix and cytosol in view of the lower Km of APX. Efficient scavenging of the H2O2 in these two ferns was evident from DAB staining.
Low doses of O2−· and H2O2 are known to induce acclimation responses against oxidative and abiotic stresses (Gechev et al. 2002; Mittler 2017). The increased activities of the antioxidant enzymes in Pteris species might be responsible, in a large part, for lower K+ leakage in response to As. The activities of antioxidant enzymes have been reported to increase in response to different HMs. For example, SOD, CAT and guaiacol peroxidase (POD) increased in response to Pb, Cd and Hg in metal tolerant Kandelia candel (Zhang et al. 2007) and those of SOD and CAT in P. vittata due to As (Srivastava et al. 2005). The decrease in enzyme activity after 18 h of As treatment (SOD in P. vittata) and (CAT in P. vittata and P. cretica) might reflect a stabilised cell metabolic state. A similar Cu-dependent enhancement of SOD and APX activities in P. vittata and P. cretica is consistent with the role of these enzymes in imparting tolerance against membrane damage.
The responses of antioxidant enzymes to As in P. microrhizoma differed strongly from those in P. vittata and P. cretica. Thus, the SOD activity increased abruptly at lower As concentrations followed by a decline. This was apparently not sufficient as is clear from the accumulation of O2−· in the As-treated pinna segments. The conversion of O2−· to H2O2 as well as the As-indifferent CAT activities possibly led to the accumulation of damaging levels of H2O2 concentrations. The DAB staining results support the assertion. The increased activity of APX after 18 h in As-treated segments of P. microrhizoma was obviously inadequate in coping with H2O2. The elevated H2O2 levels trigger alterations in Ca2+ fluxes and activities (Harper et al. 2004) which have rapid and short-term effects. The stress-specific transient Ca2+ oscillations can stimulate enzymes as NAD(P)H oxidase through specific Ca2+-interacting proteins (Harper et al. 2004). The high doses of O2−· and H2O2 were shown to trigger cell death (Gechev et al. 2002; Stone and Yang 2006). The present findings are in agreement with those of Mascher et al. (2002) who linked the As sensitivity to oxidative stress resulting from cellular damage due to enhanced lipid peroxidation, H2O2 accumulation and up-regulation or deactivation of several scavenging enzymes. Taken together, the differential As tolerance of the ferns belonging to Pteridaceae and Polypodiaceae, assigned based on the rapid leaf segment assay, could be ascribed to the difference in As-índuced oxidative stress and antioxidant defence abilities. The findings have implications for selection of As tolerant and hyperaccumulator ferns for possible inclusion in phytoremediation applications.
References
Beauchamp C, Fridovich I (1971) Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal Biochem 44:276–287
Bhattacharya P, Welch AH, Stollenwerk KG, McLaughlin MJ, Bundschuh J, Panaullah G (2007) Arsenic in the environment: biology and chemistry. Sci Total Environ 379(2):109–120
Bleeker PM, Hakvoort HW, Bliek M, Souer E, Schat H (2006) Enhanced arsenate reduction by a CDC25-like tyrosine phosphatase explains increased phytochelatin accumulation in arsenate-tolerant Holcus lanatus. Plant J 45:917–929
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1–2):248–254
Brune A, Urbach W, Dietz KJ (1995) Differential toxicity of heavy metals is partly related to a loss of preferential extraplasmic compartmentation: a comparison of Cd-, Mo-, Ni- and Zn-stress. New Phytol 129:403–409
Cai C, Lanman NA, Withers KA, DeLeon AM, Wu Q, Gribskov M, Salt DE, Banks JA (2019) Three genes define a bacterial-like arsenic tolerance mechanism in the arsenic hyperaccumulating fern Pteris vittata. Curr Biol 29(10):1625–1633
Cho M, Chardonnens AN, Dietz KJ (2003) Differential heavy metal tolerance of Arabidopsis halleri and Arabidopsis thaliana: a leaf slice test. New Phytol 158(2):287–293
Claveria RJR, Perez TR, Apuan MJB, Apuan DA, Perez REC (2019) Pteris melanocaulon Fée is an As hyperaccumulator. Chemosphere. https://doi.org/10.1016/j.chemosphere.2019.124380
De Vos CH, Schat H, Vooijs R, Ernst WHO (1989) Copper-induced damage to the permeability barrier in roots of Silene cucubalus. J Plant Physiol 135(2):164–169
Demidchik V (2010) Reactive oxygen species, oxidative stress and plant ion channels. In: Demidchik V, Maathuis FJM (eds) Ion channels and plant stress responses. Springer, Berlin
Demidchik V, Straltsova D, Medvedev SS, Pozhvanov GA, Sokolik A, Yurin V (2014) Stress-induced electrolyte leakage: the role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J Exp Bot 65(5):1259–1270
Ellis DR, Gumaelius L, Indriolo E, Pickering IJ, Banks JA, Salt DE (2006) A novel arsenate reductase from the arsenic hyperaccumulating fern Pteris vittata. Plant Physiol 141(4):1544–1554
Francesconi K, Visoottiviseth P, Sridokchan W, Goessler W (2002) Arsenic species in an arsenic hyperaccumulating fern, Pityrogramma calomelanos: a potential phytoremediator of arsenic-contaminated soils. Sci Total Environ 284(1):27–35
Gechev T, Gadjev I, Van Breusegem F, Inze D, Dukiandjiev S (2002) Hydrogen peroxide protects tobacco from oxidative stress by inducing a set of antioxidant enzymes. Cell Mol Life Sci 59:708–714
Gerhardt KE, Gerwing PD, Greenberg BM (2017) Opinion: taking phytoremediation from proven technology to accepted practice. Plant Sci 256:170–185
Goldstein DB (1968) A method for assay of catalase with the oxygen cathode. Anal Biochem 24(3):431–437
Harper JE, Breton G, Harmon A (2004) Decoding Ca+ signals through plant protein kinases. Annu Rev Plant Biol 55:263–288
He Z, Yan H, Chen Y, Shen H, Xu W, Zhang H, Shi L, Zhu Y-G, Ma M (2016) An aquaporin PvTIP4;1 from Pteris vittata may mediate arsenite uptake. New Phytol 209:746–761
Henke K (2009) Arsenic: environmental chemistry, health threats and waste treatment. Wiley, Great Britain
Hossain MA, Asada K (1984) Inactivation of ascorbate peroxidase in spinach chloroplasts on dark addition of hydrogen peroxide: its protection by ascorbate. Plant Cell Physiol 25:1285–1295
Indriolo E, Na G, Ellis D, Salt DE, Banks JA (2010) A vacuolar arsenite transporter necessary for arsenic tolerance in the arsenic hyperaccumulating fern Pteris vittata is missing in flowering plants. Plant Cell 22(6):2045–2057
Janicka-Russak M, Kabała K, Burzyński M, Kłobus G (2008) Response of plasma membrane H+-ATPase to heavy metal stress in Cucumis sativus roots. J Exp Bot 59(13):3721–3728
Kalve S, Ketan Sarangi B, Pandey RA, Chakrabarti T (2011) Arsenic and chromium hyperaccumulation by an ecotype of Pteris vittata-prospective for phytoextraction from contaminated water and soil. Curr Sci 100(6):888–894
Kavanaugh MP, Varnum MD, Osborne PB, Christie MJ, Busch AE, Adelman JP, North RA (1991) Interaction between tetraethylammonium and amino acid residues in the pore of cloned voltage-dependent potassium channels. J Biol Chem 266(12):7583–7587
Krämer U (2010) Metal hyperaccumulation in plants. Annu Rev Plant Biol 61:517–534
Kumar V, Vogelsang L, Seidel T, Schmidt R, Weber M, Reichelt M et al (2018) Interference between arsenic-induced toxicity and hypoxia. Plant Cell Environ 42(2):574–590
Lindsay ER, Maathuis FJM (2017) New molecular mechanisms to reduce arsenic in crops. Trends Plant Sci 22:1016–1026
Luongo T, Ma LQ (2005) Characteristics of arsenic accumulation by Pteris and non-Pteris ferns. Plant Soil 277(1–2):117–126
Ma LQ, Komar KM, Tu C, Zhang W, Cai Y, Kennelley ED (2001) A fern that hyperaccumulates arsenic. Nature 409(6820):579–579
Ma JF, Yamaji N, Mitani N, Xu XY, Su YH, McGrath SP, Zhao FJ (2008) Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc Natl Acad Sci 105(29):9931–9935
Ma J, Lei E, Lei M, Liu Y, Chen T (2018) Remediation of arsenic contaminated soil using malposed intercropping of Pteris vittata L. and maize. Chemosphere 194:737–744
Maathuis FJ, Ichida AM, Sanders D, Schroeder JI (1997) Roles of higher plant K+ channels. Plant Physiol 114(4):1141–1149
Mascher R, Lippmann B, Holzinger S, Bergmann H (2002) Arsenate toxicity: effects on oxidative stress response molecules and enzymes in red clover plants. Plant Sci 163(5):961–969
Meharg AA, Hartley-Whitaker J (2002) Arsenic uptake and metabolism in arsenic resistant and non resistant plant species. New Phytol 154(1):29–43
Meliker JR, Wahl RL, Cameron LL, Nriagu JO (2007) Arsenic in drinking water and cerebrovascular disease, diabetes mellitus and kidney disease in Michigan: a standardized mortality ratio analysis. Environ Health 6(4):1–11
Mimura T, Dietz KJ, Kaiser W, Schramm MJ, Kaiser G, Heber U (1990) Phosphate transport across biomembranes and cytosolic phosphate homeostasis in barley leaves. Planta 180(2):139–146
Mittler R (2017) ROS are good. Trends Plant Sci 22(1):11–19
Murphy A, Taiz L (1995) A new verical mesh transfer technique for metal- tolerance studies in Arabidopsis (Ecotypic variation and copper-sensitive mutants). Plant Physiol 108:29–38
Pilon-Smits E (2005) Phytoremediation. Annu Rev Plant Biol 56:15–39
Punshon T, Jackson BP, Meharg AA, Warczack T, Scheckel K, Guerinot ML (2017) Understanding arsenic dynamics in agronomic systems to predict and prevent uptake by crop plants. Sci Total Environ 581:209–220
Sagi M, Fluhr R (2006) Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol 141:336–340
Schat H, Vooijs R (1997) Multiple tolerance and co-tolerance to heavy metals in Silene vulgaris: a co-segregation analysis. New Phytol 136(3):489–496
Shabala S, Pottosin I (2014) Regulation of potassium transport in plants under hostile conditions: implications for abiotic and biotic stress tolerance. Physiol Plant 151(3):257–279
Sharma SS, Dietz KJ (2009) The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci 14(1):43–50
Sharma SS, Kaul S, Metwally A, Goyal KC, Finkemeier I, Dietz KJ (2004) Cadmium toxicity to barley (Hordeum vulgare) as affected by varying Fe nutritional status. Plant Sci 166(5):1287–1295
Sharma SS, Dietz KJ, Mimura T (2016) Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants. Plant Cell Environ 39(5):1112–1126
Singh N, Ma LQ, Srivastava M, Rathinasabapathi B (2006) Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L. and Pteris ensiformis L. Plant Sci 170(2):274–282
Song WY, Park J, Mendoza-Cozatl DG, Suter-Grotemeyer M, Shim D, Hortensteiner S, Geisler M, Weder B, Rea PA, Rentsch D, Schroeder JI, Lee Y, Martinoia E (2010) Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin transporters. Proc Natl Acad Sci USA 107:21187–21192
Srivastava M, Ma LQ, Singh N, Singh S (2005) Antioxidant responses of hyper-accumulator and sensitive fern species to arsenic. J Exp Bot 56(415):1335–1342
Srivastava M, Santos J, Srivastava P, Ma LQ (2010) Comparison of arsenic accumulation in 18 fern species and four Pteris vittata accessions. Bioresour Technol 101(8):2691–2699
Stone JR, Yang S (2006) Hydrogen peroxide: a signalling messenger. Antioxid Redox Signal 8(3–4):243–270
Strange J, Macnair MR (1991) Evidence for a role for the cell membrane in copper tolerance of Mimulus guttatus Fischer ex DC. New Phytol 119(3):383–388
Thakur S, Choudhary S, Majeed A, Singh A, Bhardwaj P (2019) Insights into the molecular mechanism of arsenic phytoremediation. J. Plant Growth Reg 38(3):1141–2115
Thompson J, Begenisich T (2003) External TEA block of shaker K+ channels is coupled to the movement of K+ ions within the selectivity filter. J Gen Physiol 122(2):239–246
Tu C, Ma LQ (2003) Effects of arsenate and phosphate on their accumulation by an arsenic-hyperaccumulator Pteris vittata L. Plant Soil 249(2):373–382
Wainwright SJ, Woolhouse HW (1977) Some physiological aspects of copper and zinc tolerance in Agrostis tenuis Sibth. cell elongation and membrane damage. J Exp Bot 28(4):1029–1036
Wan X, Lei M (2018) Intercropping efficiency of four arsenic hyperaccumulator Pteris vittata populations as intercrops with Morus alba. Environ Sci Pollut Res 25(13):12600–12611
Zhang FQ, Wang YS, Lou ZP, Dong JD (2007) Effect of heavy metal stress on antioxidative enzymes and lipid peroxidation in leaves and roots of two mangrove plant seedlings Kandelia candel and Bruguiera gymnorrhiza. Chemosphere 67(1):44–50
Zhao FJ, Dunham SJ, McGrath SP (2002) Arsenic hyperaccumulation by different fern species. New Phytol 156(1):27–31
Zhao FJ, Wang JR, Barker JHA, Schat H, Bleeker PM, McGrath SP (2003) The role of phytochelatins in arsenic tolerance in the hyperaccumulator Pteris vittata. New Phytol 159:403–410
Acknowledgements
VK thankfully acknowledges the financial assistance from ICMR (India) and DAAD (Germany). Valuable suggestions concerning data presentation, statistical analysis and manuscript writing by Lara Vogelsang are gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Kumar, V., Dietz, KJ. & Sharma, S.S. Differential Arsenic-Induced Membrane Damage and Antioxidant Defence in Isolated Pinna Segments of Selected Fern Species from Western Himalaya. J Plant Growth Regul 40, 855–866 (2021). https://doi.org/10.1007/s00344-020-10145-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-020-10145-w