
Abstract
Analyses of maximum early tree ring growth within a regional curve standardization (RCS) framework were used to construct ring width chronologies in mm year−1 to estimate historical growth rates in southwest Yukon (Canada), without the necessity of correcting for age related changes. The analysis relied on the use of the 11th–50th year growth rings to standardize chronology comparisons. This age range represents the period of peak radial growth for Picea albertiana and P. glauca (white spruces), which are common trees on upland boreal sites in the study area. Of 1,460 available Picea ring series, 1,029 fit the criteria for inclusion in the study and they represented five commonly occurring RCS curve types. When converted to chronologies, all curve types had year-to-year fluctuations in average ring widths (RW11–50), but nine of the 12 RCS types (four types in three study areas) also had long-term growth rate increases of 15–132% between ad 1850 and 2000, based on regression modeling (R/r = 0.54–0.76, p < 0.001). Associated with these increases were changes through time in RCS curve type and the timing of peak growth. Among studied areas, RW11–50 values on average increased 47% from 1.59 to 2.35 mm year−1 between 1850 and 2000. The greatest increase (71% vs 35–38%) occurred in the coldest study area. RW11–50 values were significantly (p < 0.001) and strongly correlated (r > 0.94, n > 100) with conventional tree ring width index values derived from multiple RCS curves. This indicates that the two chronology construction approaches have very similar temporal patterns despite the maximum early growth analysis technique representing ring widths in mm.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In boreal forests, annual tree ring widths typically vary because of year-to-year fluctuations in climate. Climate is influential because radial growth is a physiological response by the vascular cambium to the simultaneous presence of adequate heat and moisture, which control production of hormones that stimulate cell division and growth (Catesson and Lachaud 1993; Oberhuber et al. 2014). Therefore, growth rings are used to model long-term changes in climate and they could be useful for interpreting the historical development of present-day forests and their associated archaeologically significant events (Bleicher 2014). Radial growth rate (mm year−1) studies for these purposes are rare (for example, Nicolussi et al. 1995; Helama 2015) compared to those that use conventional ring width index values. The primary reason for this lack of use is probably the decrease in annual ring widths as trees age, which makes it difficult to calculate long-term changes in tree growth rates without conducting some form of numerical data standardization.
Removal of the age related growth pattern from ring series through curve fitting and the calculation of tree ring width index values (RWI) derived from measured widths divided by expected widths (Melvin and Briffa 2008) rescales and standardizes ring widths through time and among chronosequences. Within RWI chronologies, a value of 1.0 corresponds to the average growth rate regardless of the actual width, with greater or lesser values representing proportional changes. Because ring growth is represented on a relative scale, actual radial growth (mm) and rates of change with time are lost. These data losses are not typically an issue when RWI values are used as proxies for palaeotemperature (for example, Youngblut and Luckman 2008) and palaeoprecipitation reconstructions (for example, Szymczak et al. 2014). They are, however, essential when interpreting forest development. If variation in actual ring widths could be standardized and temporally modeled, it would provide a basis for better understanding the historical development of present-day forests (Helama 2015). Tree ring chronologies based on width (mm) might also be useful for predicting the effects of future climate warming on boreal forest productivity, since the long-term implications are considered “highly uncertain” (Intergovernmental Panel on Climate Change 2013, p. 71).
The purpose of this study was to determine if, when, and how much upland Picea albertiana (western white spruce) and P. glauca (white spruce) potential radial growth rates (mm year−1) have changed in the boreal forests of southwest Yukon, northwest Canada since the mid 19th century.
Study areas
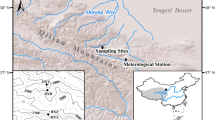
Tree ring cores were collected from the Beaver Creek, Carmacks, and Takhini Valley areas of southwest Yukon, all with high latitude boreal climates. Each area has widespread P. albertiana or P. glauca, and these trees dominate the western half of the Takhini Valley and the uplands of the Beaver Creek study area, respectively (Table 1). Although considered different taxa for phylogenic reasons (Strong and Hills 2006), both have similar ecological requirements as well as the same physiognomy and morphology, except for the shapes of their cone scales.
In addition to Picea spp., Pinus contorta var. latifolia (lodgepole pine) stands commonly occur in the eastern third of the Takhini Valley and sporadically in the Carmacks area as a postfire species. Populus tremuloides (trembling aspen) stands occur in the eastern half of the Takhini Valley, and on warmer submesic sites at low elevations in the Carmacks area (Table 1). Grasslands occupy the warmest and driest sites in these two areas (Chambers 2010; Strong 2015), but are absent in the Beaver Creek area. The presence of P. tremuloides and P. contorta stands in the local vegetation was primarily due to the sporadic occurrence of wildfires, which impede the development of Picea-dominated forests that form the climatic climax vegetation for all three areas. The last major wildfires in the Carmacks and Takhini Valley areas occurred in 1958 and at some unknown time before 1946 in the Beaver Creek area (Natural Resources Canada 2003).
The topography of each study area includes both low and steep gradient terrain, but in different configurations (Table 1). From May through September, the summer temperatures in 1984–2007 were similar in the three areas, with Beaver Creek being the coolest (Table 1). The Beaver Creek area also received ~ 40% more summer precipitation than the Carmacks and Takhini Valley areas, which resulted in less severe summer heat-moisture index values (Spittlehouse 2006). Because of differences in vegetation, each study area was considered to represent a somewhat different boreal climate (Table 1).
Materials and methods
Tree cores were collected only from P. albertiana and P. glauca, with these species treated as a single taxon for ring width analysis purposes. Field sampling was done in 2013–2014 in the Takhini Valley and in 2017 at Beaver Creek and Carmacks. Tree coring was done at a median height of 130 cm above the ground. Core preparation and ring width measurement followed procedures described by Strong (2017). Briefly, cores were collected on well drained, low gradient sites and southerly slopes, which were scattered across each study area. Cores were extracted from the largest available tree on a site, a stand dominant but not necessarily old growth, and only the longest and best quality core was retained for analysis due to the common occurrence of asymmetrical stem growth. It was assumed that the inter-annual radial growth variation was proportional along different stem radii (Speer 2010). The collection of two cores per tree has been suggested (Speer 2010), but use of a single core is not an uncommon practice (for example, Griesbauer and Green 2012; Chavardés et al. 2013; Helama et al. 2018).
Tree ring width series with similar temporal growth patterns and levels of peak growth (mm) were grouped by visual comparisons to create different regional curve standardization (RCS) curve types (Helama et al. 2017). The pith of each core was used as the common time reference. When a core did not intersect the pith, the number of missing rings was estimated from the curvature and width of the adjacent rings (Bergin and Kimberley 2012). For inclusion in the analysis, RCS curves had to include 25 or more ring series from an individual study area and to have maximum ring widths within the first 50 years of growth at coring height. The data are listed in ESM 1.
To investigate variation in growth rates with time (mm year−1) within and among RCS curve types, analysis was limited to the 11th–50th tree rings counted from the pith. All other ring data were excluded from consideration. This analysis approach is conceptually similar to that used by Nicolussi et al. (1995), except that, first, the current analysis was based on the period of maximum annual radial growth (mm) and second, ring widths were averaged rather than separately considered; hereafter referred to as “maximum early growth analysis” (MEGA). The use of a restricted age range in tree ring series avoids the age dependent bias (Nicolussi et al. 1995, p. 183) that is commonly associated with chronologies based on full length tree ring series. The first ten rings were excluded from consideration to avoid the rapidly increasing and sometimes erratic ring widths associated with early tree growth, and to negate the problem of missing, rotten, or otherwise unavailable rings, usually three or fewer. RW11–50 values were assigned calendar dates and averaged by year to create a chronology for each RCS curve type. The year of reference for each set of RW11–50 values was the pith date plus 30 years.
An RW11–50 chronology for each study area was created by averaging all available ring series values by calendar year regardless of their RCS curve type (Fig. 1w–y). However, the presented results were limited to time periods when all RCS curve types simultaneously occurred. Temporal height-radial growth relationships within each study area were assumed to have been similar between ad 1850 and 2000.

Diagrams a–e represent the most common regional curve standardization (RCS) curve types found in the Beaver Creek, Carmacks, and Takhini Valley areas of southern Yukon. The number of ring widths used to construct RCS curves a–e is indicated in diagrams f–j, respectively. Temporal variation in average 11th–50th year ring widths among RCS curves by study area is illustrated in scatter diagrams (k–v). Diagrams w–y show changes in ring width at the study area level based on the amalgamation of the diagram data from k–n (Beaver Creek), o–r (Carmacks), and s–v (Takhini Valley). Diagram z is a direct comparison of the w–y regression trend lines. The independent regression variable (x) in diagrams w–y is based on 2013 minus the calendar year. The parameter “m” indicates the number of ring series included in each analysis and SEE values are standard errors of estimate
For comparative purposes, standardized ring width index (RWI) values were calculated for each study area through the use of CRUST software (Melvin and Briffa 2013, 2014). The calculation of RWI values for each RCS curve was based on the signal-free, arithmetic mean, and ratio calculation options. The resulting values were weighted according to the number of associated samples and averaged to produce year specific sets of RWI values for each study area, thus the multiple RCS approach to chronology development (Helama et al. 2017). This manual averaging procedure was necessary, because most curves exceeded the 40 ring series per RCS curve limit of the CRUST software for multiple RCS chronology development (Melvin and Briffa 2013, p. 9).
Simple and polynomial regressions were used to model temporal trends among RW11–50 values, and to compare chronologically corresponding RWI and RW11–50 values. Pearson product moment correlation was used to assess the degree of numerical association between variables. Kolmogorov–Smirnov one-sample tests were used to assess data normality.
Results
Data characteristics
A total of 1,460 Picea growth increment cores were collected from the three study areas (Table 1). Among these, 1,029 ring series formed five RCS curve types that satisfied the data inclusion criteria (Fig. 1a–e). The remaining 29.5% of the series belonged to other RCS curve types with fewer than 25 members within a study area, or they had peak growth that began after the 50th growth year at coring height. Ninety-one percent of marker rings among the five RCS curve types complied with the established cross-dating criterion of markers being bracketed by larger rings and fewer than 2% were bracketed by smaller rings. Average ring widths ranged from 1.25 to 1.35 mm in the three study areas, with Carmacks having the widest rings (Table 1). The first 50 years of radial growth yield cores up to 166 mm in length, with study area averages of 82–94 mm that had normal distributed values (Fig. 2). All RCS curves included more than 100 ring series, with RCS curve A (RCS-A, Fig. 1a) the most common type (Fig. 1f–j).

Average tree core lengths and standard deviations (sd) of 50 year old Picea trees in the three southwest Yukon study areas
Pith formation in the RCS ring series occurred between the mid to late 18th and the mid 20th century, with an average date of 1889 (± 5–6 years) regardless of the study area (Table 1). All RCS curves reached peak growth by the age of 38 years at coring height. RCS-A had average peak ring widths of 2.25 mm, when the trees were 20–25 years old (Fig. 1a, Table 2). Two curves (Fig. 1b, d) had slightly greater average peak ring widths than RCS-A, and two had values < 1.8 mm (Table 2). The period of peak growth began either within five or after 19 years of growth (Table 2). The duration of peak growth lasted for 6–13 years, with the RCS-D ring series persisting longest (Fig. 1d, Table 2). Unlike other curve types, the RCS-E curve did not have an obvious growth peak, but rather had steadily declining ring widths after its fourth year of growth (Fig. 1e). When peak growth was initially reached, the trees were either < 4 m or ≥ 10 m tall (Table 2).
Changes in RCS growth rates
Four RCS curve types dominated each of the three study areas (Fig. 1k–v). Of these 12 RCS chronologies, nine had increasing rates of radial growth. The Beaver Creek RCS-A (Fig. 1k) and Takhini Valley RCS-E (Fig. 1v) curves had linear and relatively consistent annual ring widths that were not correlated with chronology years (p > 0.05). The longest commonly available time period represented by Fig. 1k–v was 1850–2000. Annual growth rates were consistently lower in 1850 than in the modern portion of the RCS chronologies, except the two with no significant change over time and Carmacks RCS-B with a decrease in growth (Table 3). Average Picea radial growth rates in 1850 ranged from a low of 1.070 mm year−1 in the Beaver Creek RCS-E curve up to 2.742 mm year−1 in the Carmacks RCS-B curve (Table 3). The Carmacks and Takhini RCS-B curves (Fig. 1p, t) with their sinuous regression trend lines were anomalous compared to other curve patterns from the three study areas. In the nine RCS curve types with rising growth rates, the increases ranged from 15 to 132% over the 150 year reference period, although 30–90% increases were more typical (Table 3). No consistent growth trend occurred among the RCS curve types, but the RCS-C and RCS-D types tended to increase the most.
Among the three study areas, growth rates increased by 71% in Beaver Creek during 1850–2000, but by only about half as much in the other areas (Table 3). The lines in Fig. 1z represent the regression trends of Fig. 1w–y. The Carmacks and Takhini Valley results had parallel long-term trends, with the former having a slightly greater growth rate (difference ≤ 0.18 mm year−1). These regression trend lines, however, were not significantly different at the α = 0.05 level, since they were separated by less than one standard error of estimate, even if based on the smaller of the two possibilities (SEE = 0.324, Fig. 1y). Although the Beaver Creek RW11–50 trend line was similar to those of the other two areas, its trajectory was steeper (Fig. 1z). On average, growth rates in the three study areas increased by 47% (Table 3). Based on the change in radial growth, the representative height for a 50 year old Picea tree was estimated to have increased by 3.8 m between 1850 and 2000.
Changes in RCS composition
In the early portions of Fig. 1w–y, more than half of the Beaver Creek and Carmacks chronologies are dominated by a single RCS curve type, whereas the Takhini Valley has a mixed composition (Table 4). These occurrence patterns are reversed in the modern portions of these chronologies. In the Beaver Creek chronology, a large increase occurs in the proportion of the RCS-D series at the expense of the RCS-A and -C types. The Carmacks RCS-B type and Takhini Valley RCS-A types are more common at the modern end of their chronology. No major changes (± 5% or less) occur in the abundance of ring series of the RCS-E type (Table 4). Associated with these shifts in RCS type are also changes in the timing of peak growth. In the Beaver Creek and partially in the Takhini Valley areas, peak growth is delayed up to 14 years, whereas the change toward greater RCS-B membership in the Carmacks area advances peak growth by 16 years between 1850 and 2000 (Tables 2, 4).
RWI versus RW11-50 values
Statistically significant and very strong correlations occur between tree ring width index (RWI) and RW11–50 values (Fig. 3). The poorest bivariate relationship explains 88% of the variance in RW11–50 values (r = 0.941, n = 104) (Fig. 3a). Standard errors of estimate (SEE) among study areas are less than 0.15 mm, with RW11–50 values generally ranging from 1–3 mm. Among study areas, the correlation between RWI and RW11–50 values is 0.926, with a standard error of 0.149 (n = 408).

Comparisons of Picea tree ring width indices (RWI) and 11th–50th year ring width (RW11–50) values by study area based on regression analysis. Values in brackets indicate the range of calendar years used for modeling (cf. Fig. 1w–y). Each time period includes a full complement of co-occurring regional curve standardization curve types. SEE values are standard errors of estimate
Discussion
RW11–50 chronology design considerations
The development of RW11–50 chronologies relies on the use of peak growth during the early portion of the Picea life cycle to standardize comparisons among tree ring series. When unconstrained by factors such as competition, maximum ring widths represent the most growth by Picea trees under a given set of environmental conditions, before the age related decline in growth begins to have a noticeable effect. The focus on peak radial growth and use of locally dominant trees suggests that RW11–50 values should be regarded as the realistically maximum achievable growth rates rather than average amounts. Theoretical maxima would approximate the upper limit of RW11–50 values in Fig. 1w–y (for example, RW11–50 + 1.95 SEE). Maximum achievable growth is probably a better measure for interpreting forest history, because average growth includes the limitations imposed by stand density and competition, which can vary considerably, even within small areas. Hence maximum achievable growth better reflects the realistic climatic growth potential. Use of dominant trees and the longest available core is consistent with the aim of estimating maximum achievable growth.
Besides the possibility of estimating tree annual ring widths on a standardized basis, the maximum early growth analysis (MEGA) approach to chronology construction has practical advantages. First, its development does not require the collection of new data, just their reorganization, assuming that raw ring width measurements already exist, such as those previously used for RWI chronology construction. For this reason, the RWI and the MEGA approaches could be simultaneously developed for interpretation in parallel, with minimal additional effort. Secondly, the MEGA approach allows differentiation of study areas based on growth rates, because RWI chronologies have means of 1.0, even if differences exist. Lastly, the MEGA approach is a more simplistic method than the procedures described by Briffa et al. (2001) and Helama (2015). A disadvantage of the approach is the need for a large number of tree ring series compared with what might be collected from small fixed area sampling plots. For this reason, the MEGA approach is better suited for interpreting forest histories at a landscape rather than a plot level.
“Modern sampling bias” was not considered an issue for the RW11–50 chronologies (Fig. 1w–y) and, therefore, not for the associated growth rate calculations, because: 1, multiple RCS curves largely negate the problem (Melvin et al. 2012); 2, inclusion of only calendar years when all RCS curve types simultaneously occur avoids under- and over-representation of RCS curve types with different growth rates; 3, use of large numbers of ring series helps to ensure that a broad spectrum of growth rates is included in the dataset, although oriented towards dominant trees; and 4, normally distributed 50 year core lengths (Fig. 2) indicate that growth rates are not skewed towards representing faster or slower growing trees. In addition, the period covered by the analysis is short (< 200 years). Therefore, tree ages of 150–200 years or less are well within the 300–350 year natural life expectancy of Picea albertiana and P. glauca (Nienstaedt and Zasada 1990), so age related mortality biases due to differences in growth rates should be minimal.
The temporal limits of the peak growth period may require adjustment to accommodate other tree species in different ecological settings. A review of published RCS curves suggests the 11–50 year period may be too broad when RCS growth patterns approximate a negative exponential or a Hugershoff distribution (for example, Becker 1989; Helama et al. 2008, 2018). When setting biological age limits for a maximum early growth analysis, some balance is needed among RCS curve types so as to maximize the length of the analysis period, while minimizing age related effects both before and after peak growth. Longer inclusion periods increase the number of ring widths available for analysis, but have the disadvantage of increasing growth variation that could weaken an analysis.
Forest tree growth
In addition to a 47% increase in 50 year old Picea growth rates, tree heights potentially increased 26% and stem cross-sectional areas at coring height more than doubled between 1850 and 2000 in southwest Yukon (Table 3). With respect to radial growth, Nicolussi et al. (1995) and Bergès et al. (2000) reported 40–50% increases respectively in European subalpine Pinus cembra and Quercus petraea between the mid 19th and late 20th century, whereas much greater rates, 73% and 106%, were estimated for western North American Pinus longaeva (LaMarche et al. 1984). This suggests that the estimated growth rate changes in southern Yukon are not unrealistic. Despite the increase in RW11–50 values, contemporary regional growth rates of 2.348 mm year−1 are only moderate compared to ~ 4 mm year−1 reported elsewhere in western North America (for example, Huang and Titus 1995, Fig. 2; Sattler and Stewart 2016, estimated from Table 1). This lower radial growth rate likely reflects the cooler and shorter growing seasons of high latitude environments compared to their lower latitude counterparts. Presumably, the poorer growth around 1850 was associated with even colder climatic conditions than occurred in subsequent years. An extrapolation from Takhini Valley RWI values of 0.6 suggests RW11–50 growth rates of 1.166 mm year−1 from Strong (2017), based on the regression equation in Fig. 3c, and show that stem diameters (12.9 cm) and areas (130 cm2), and tree heights (11.8 m) under favourable growing conditions, were likely much less during the early 19th century (for calculation method, see Table 3, footnote “d”). On a relative basis, contemporary 50 year old trees are probably 2–3.7 times larger than their 1850 and early 1800 s counterparts, respectively, based on differences in tree stem area (Table 3). These substantial differences likely occurred because the early 19th century coincided with lower temperatures in southwest Yukon (Davi et al. 2003; Youngblut and Luckman 2008) and the rest of the northern hemisphere (D’Arrigo et al. 2006), the coldest during the past two centuries. Specifically, the early 19th century coincided with part of the Little Ice Age, ad 1450–1850 (Intergovernmental Panel on Climate Change 2013) with the compounding affects of the Dalton solar minimum ~ 1795–1820 (Vaquero and Trigo 2015). This suggests that increases in tree growth since the early 19th century were a response to a long term climatic warming pattern. Because the three study areas likely experienced similar levels of atmospheric warming (Davi et al. 2003; Youngblut and Luckman 2008) and CO2 enrichment (Tissue et al. 1997) after 1850, greater annual radial growth rate increases probably occurred in the Beaver Creek area due to longer periods of sustained moisture availability during the growing season than were the case in the Carmacks and Takhini Valley areas (Table 1). This greater availability of moisture occurred because of both greater summer precipitation and cooler summer temperatures. In contrast, evapotranspiration demands in the Carmacks and Takhini Valley areas often substantially exceed the early summer moisture supply (Jätzold 2000), when radial growth typically begins (Rossi et al. 2013), which would tend to limit tree growth.
Associated with increases in Picea radial growth rates are also changes in the timing of peak growth due to shifts in the RCS curve type composition. The delayed occurrence of peak growth in the Beaver Creek and Takhini Valley areas between 1850 and 2000 prolonged the period of continually increasing radial growth rates before reaching a plateau. This suggests that juvenile trees were establishing themselves in forest canopy gaps where access to photosynthetic radiation and growing space was more readily available than under a forest canopy. In the Carmacks area, an earlier peak (~ 16 years) resulted in Picea reaching its maximum growth rate when < 6 years old at coring height. This change implies that present day Picea most commonly grow in forest understories rather than in gaps in the canopy, and that forest canopies are now more closed than they were in the past. This shift in recruitment patterns, a combination of seed germination, survival and subsequent growth (Eriksson and Ehrlén 2008), also suggests that Picea stands were more abundant in the 1850s than today, or that postfire forest lacked continuous overstories.
A change in Picea recruitment patterns appears to be supported by the trend of increasing ring widths in the middle of Fig. 1p, which coincides with a time of extensive logging that occurred to fuel sternwheeler river boats, which travelled the Yukon River from the late 1890s to the mid 1950s (Dobrowolsky and Johnson 2013). The cessation of logging due to a lack of demand would have allowed the development of structurally mature forests, which would have suppressed juvenile Picea growth beneath their canopies, and this could be represented by the steady post 1950 decline in RW11–50 values (Fig. 1p). Understory vegetation alone, due to its low stature, typically less than 75 cm tall and of modest density in the Carmacks area (Strong 2009, 2014), is unlikely to have slowed juvenile Picea growth after the trees reached coring height.
Conclusions
The very strong correlation between tree ring width index (RWI) and RW11–50 values indicates that the latter is as valid a method as the conventional RWI approach for tree ring chronology construction. An advantage of the MEGA approach is the ability to estimate annual ring widths from which long term growth rates and past tree metrics can be estimated for forest history interpretation. Furthermore, essentially no numerical manipulation of the raw ring width data is necessary. RWI and RW11–50 chronologies represent complementary rather than alternative approaches to tree ring analysis.
References
Becker M (1989) The role of climate on present and past vitality of silver fir forests in the Vosges Mountains of Northeastern France. Can J For Res 19:1,110–1,117
Bergès L, Dupouey J-L, Franc A (2000) Long-term changes in wood density and radial growth of Quercus petraea Liebl. in northern France since the middle of the nineteenth century. Trees 14:398–408
Bergin DO, Kimberley MO (2012) Reliability of increment core growth ring counts as estimates of stand age in totara (Podocarpus totara D. Don). N Z J For Sci 42:131–141
Bleicher N (2014) Four levels of patterns in tree-rings: an archaeological approach to dendroecology. Veget Hist Archaeobot 23:615–627
Briffa KR, Osborn TJ, Schweingruber FH et al (2001) Low-frequency temperature variations from a northern tree ring density network. J Geophys Res 106D:2,929–2,941
Catesson A-M, Lachaud S (1993) Le cambium, structure, fonctionnement et contrôle de l’activité saisonnière. Acta Bot Gallica 140:337–350
Chambers JHS (2010) Habitat use and ecologically sustainable carrying capacity for elk (Cervus elaphus) in the Takhini Valley, Yukon. Thesis, University of Calgary
Chavardès RD, Daniels LD, Waeber PO et al (2013) Unstable climate—growth relations for white spruce in southwest Yukon, Canada. Clim Change 116:593–611
D’Arrigo R, Wilson R, Jacoby G (2006) On the long-term context for late twentieth century warming. J Geophys Res 111:D031303
Davi NK, Jacoby GC, Wiles GC (2003) Boreal temperature variability inferred from maximum latewood density and tree-ring width data, Wrangell Mountain region, Alaska. Quat Res 60:252–262
Dobrowolsky H, Johnson L (2013) Whitehorse, an illustrated history. Figure 1 Publishing, Vancouver
Environment Canada (2018) Historical climate data. http://climate.weather.gc.ca. Accessed 2 Aug 2018
Eriksson O, Ehrlén J (2008) Seedling recruitment and population ecology. In: Leck MA, Parker VT, Simpson RL (eds) Seedling ecology and evolution. Cambridge University Press, Cambridge, pp 239–254
Government of British Columbia (2019) HectaresBC, summer heat-moisture index. https://www.hectaresbc.org/app/habc/HaBc.htm|?type=rasterquery=climatenormal.shm. Accessed 30 May 2019
Griesbauer HP, Green DS (2012) Geographical and temporal patterns in white spruce climate—growth relationships in Yukon, Canada. For Ecol Manag 267:215–227
Helama S (2015) Expressing tree-ring chronology as age-standardized growth measurements. For Sci 61:817–828
Helama S, Vartiainen M, Koström T et al (2008) X-ray microdensitometry applied to subfossil tree-rings: growth characteristics of ancient pines form the southern boreal forest zone in Finland at intra-annual and centennial time-scales. Veget Hist Archaeobot 17:675–686
Helama S, Melvin TM, Briffa KR (2017) Regional curve standardization: state of the art. Holocene 27:172–177
Helama S, Sohar K, Läänelaid A et al (2018) Reconstruction of precipitation variability in Estonia since the eighteenth century, inferred from oak and spruce tree rings. Clim Dyn 50:4,083–4,101
Huang S, Titus SJ (1995) An individual tree diameter increment model for white spruce in Alberta. Can J For Res 25:1,455–1,465
Intergovernmental Panel on Climate Change (2013) Climate change 2013: The physical science basis, technical summary. Cambridge University Press, Cambridge
Jätzold R (2000) Semi-arid regions of the boreal zone as demonstrated in the Yukon Basin. Erdkunde 54:1–19
LaMarche VC Jr, Graybill DA, Fritts HC et al (1984) Increasing atmospheric carbon dioxide: Tree ring evidence for growth enhancement in natural vegetation. Science 225:1,019–1,021
Malone T, Liang J (2009) A bark thickness model for white spruce in Alaska northern forests. Int J For Res 2009:876,965
Melvin TM, Briffa KR (2008) A “signal-free” approach to dendroclimatic standardization. Dendrochronologia 26:71–86
Melvin TM, Briffa KR (2013) CRUST: Climatic Research Unit standardization program documentation. University of East Anglia, Norwich. https://crudata.uea.ac.uk/cru/software/crust/
Melvin TM, Briffa KR (2014) CRUST: software for the implementation of regional chronology standardisation: Part 1. Signal-free RCS. Dendrochronologia 32:7–20
Melvin TM, Gradd H, Briffa KR (2012) Potential bias in ‘updating’ tree-ring chronologies using regional curve standardization: re-processing 1500 years of Torneträsk density and ring-width data. Holocene 23:364–373
Natural Resources Canada (2003) Yukon fire history atlas: 1946–2001. http://emrlibrary.gov.yk.ca/maps/fire-history-atlas/start.html. Accessed 26 Sept 2018
Nicolussi K, Bortenschlager S, Körner C (1995) Increase in tree-ring width in subalpine Pinus cembra from the central Alps that may be CO2-related. Trees 9:181–189
Nienstaedt H, Zasada JC (1990) Picea glauca (Moench) Voss (white spruce). In: Burns RM, Honkala BH (Tech Co-ords) Silvics of North America: 1. Conifers. Agriculture Handbook 654 USDA Forestery Service, Washington pp 204–226
Oberhuber W, Gruber A, Kofler W et al (2014) Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur J For Res 133:467–479
Oswald ET, Brown BN (1986) Forest communities in Lake Laberge Ecoregion, Yukon Territory. Inf Rep BC-X-282. Canadian Forestry Service, Victoria
Rossi S, Anfodillo T, Čufur K et al (2013) A meta-analysis of cambium phenology and growth: linear and non-linear patterns in conifers of the northern hemisphere. Ann Bot 112:1,911–1,920
Sattler DF, Stewart JD (2016) Climate, location, and growth relationships with wood stiffness at the site, tree, and ring levels in white spruce (Picea glauca) in the Boreal Plains ecozone. Can J For Res 46:1,235–1,245
Speer JH (2010) Fundamentals of tree-ring research. University of Arizona Press, Tucson
Spittlehouse D (2006) ClimateBC: your access to interpolated climate data for BC. Streamline 9:16–21
Strong WL (2009) Populus tremuloides Michx. postfire stand dynamics in the northern boreal-cordilleran ecoclimatic region of central Yukon Territory, Canada. For Ecol Manag 258:1,110–1,120
Strong WL (2013) Ecoclimatic zonation of Yukon and ecoclinal variation in vegetation. Arctic 66:52–67
Strong WL (2014) Northernmost North American Pinus contortai var. latifolia (lodgepole pine) sociations and vegetation diversity relative to its central range east of the Rocky Mountains. Nord J Bot 32:222–232
Strong WL (2015) High-latitude Yukon boreal-cordilleran grassland plant communities. Arctic 68:69–78
Strong WL (2017) Episodic and systematic tree ring-width variation (ad 1763–2013) in the Takhini Valley, southwest Yukon, Canada. Arctic 70:389–402
Strong WL, Hills LV (2006) Taxonomy and origin of present-day morphometric variation in Picea glauca (× engelmannii) seed-cone scales in North America. Can J Bot 84:1,129–1,141
Szymczak S, Hetzer T, Bräuning A et al (2014) Combining wood anatomy and stable isotope variations in a 600-year multi-parameter climate reconstruction from Corsican black pine. Quat Sci Rev 101:146–158
Tissue DT, Thomas RB, Strain BR (1997) Atmospheric CO2 enrichment increases growth and photosynthesis of Pinus taeda: a 4 year experiment in the field. Plant Cell Environ 20:1,123–1,134
Vaquero JM, Trigo RM (2015) Redefining the limit dates for the Maunder Minimum. New Astron 34:120–122
Youngblood D, Luckman B (2008) Maximum June-July temperatures in the southwest Yukon over the last 300 years reconstructed from tree rings. Dendrochronologia 25:153–166
Acknowledgements
Three anonymous reviewers provided constructive comments that improved the completeness of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by K. Brown.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Strong, W.L. Picea (spruce) growth rate (mm year−1) changes in southwest Yukon (Canada) since the mid 19th century. Veget Hist Archaeobot 29, 91–100 (2020). https://doi.org/10.1007/s00334-019-00737-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-019-00737-2